Abstract
Bone morphogenetic proteins (BMPs) are multi-functional growth factors that belong to the transforming growth factor β (TGFβ) superfamily. The roles of BMPs in embryonic development and cellular functions in postnatal and adult animals have been extensively studied in recent years. Signal transduction studies have revealed that Smad1, 5 and 8 are the immediate downstream molecules of BMP receptors and play a central role in BMP signal transduction. Studies from transgenic and knockout mice and from animals and humans with naturally occurring mutations in BMPs and related genes have shown that BMP signaling plays critical roles in heart, neural and cartilage development. BMPs also play an important role in postnatal bone formation. BMP activities are regulated at different molecular levels. Preclinical and clinical studies have shown that BMP-2 can be utilized in various therapeutic interventions such as bone defects, non-union fractures, spinal fusion, osteoporosis and root canal surgery. Tissue-specific knockout of a specific BMP ligand, a subtype of BMP receptors or a specific signaling molecule is required to further determine the specific role of a BMP ligand, receptor or signaling molecule in a particular tissue.
BMPs are members of the TGFβ superfamily. The activity of BMPs was first identified in the 1960s (Urist, M.R. (1965) “Bone formation by autoinduction”, Science 150, 893–899), but the proteins responsible for bone induction remained unknown until the purification and sequence of bovine BMP-3 (osteogenin) and cloning of human BMP-2 and 4 in the late 1980s (Wozney, J.M. et al. (1988) “Novel regulators of bone formation: molecular clones and activities”, Science 242, 1528–1534; Luyten, F.P. et al. (1989) “Purification and partial amino acid sequence of osteogenin, a protein initiating bone differentiation”, J. Biol. Chem. 264, 13377–13380; Wozney, J.M. (1992) “The bone morphogenetic protein family and osteogenesis”, Mol. Reprod. Dev. 32, 160–167). To date, around 20 BMP family members have been identified and characterized. BMPs signal through serine/threonine kinase receptors, composed of type I and II subtypes. Three type I receptors have been shown to bind BMP ligands, type IA and IB BMP receptors (BMPR-IA or ALK-3 and BMPR-IB or ALK-6) and type IA activin receptor (ActR-IA or ALK-2) (Koenig, B.B. et al. (1994) “Characterization and cloning of a receptor for BMP-2 and BMP-4 from NIH 3T3 cells”, Mol. Cell. Biol. 14, 5961–5974; ten Dijke, P. et al. (1994) “Identification of type I receptors for osteogenic protein-1 and bone morphogenetic protein-4”, J. Biol. Chem. 269, 16985–16988; Macias-Silva, M. et al. (1998) “Specific activation of Smad1 signaling pathways by the BMP7 type I receptor, ALK2”, J. Biol. Chem. 273, 25628–25636). Three type II receptors for BMPs have also been identified and they are type II BMP receptor (BMPR-II) and type II and IIB activin receptors (ActR-II and ActR-IIB) (Yamashita, H. et al. (1995) “Osteogenic protein-1 binds to activin type II receptors and induces certain activin-like effects”, J. Cell. Biol. 130, 217–226; Rosenzweig, B.L. et al. (1995) “Cloning and characterization of a human type II receptor for bone morphogenetic proteins”, Proc. Natl Acad. Sci. USA 92, 7632–7636; Kawabata, M. et al. (1995) “Cloning of a novel type II serine/threonine kinase receptor through interaction with the type I transforming growth factor-β receptor”, J. Biol. Chem. 270, 5625–5630). Whereas BMPR-IA, IB and II are specific to BMPs, ActR-IA, II and IIB are also signaling receptors for activins. These receptors are expressed differentially in various tissues. Type I and II BMP receptors are both indispensable for signal transduction. After ligand binding they form a heterotetrameric-activated receptor complex consisting of two pairs of a type I and II receptor complex (Moustakas, A. and C.H. Heldi (2002) “From mono- to oligo-Smads: the heart of the matter in TGFβ signal transduction” Genes Dev. 16, 67–871). The type I BMP receptor substrates include a protein family, the Smad proteins, that play a central role in relaying the BMP signal from the receptor to target genes in the nucleus. Smad1, 5 and 8 are phosphorylated by BMP receptors in a ligand-dependent manner (Hoodless, P.A. et al. (1996) “MADR1, a MAD-related protein that functions in BMP2 signaling pathways”, Cell 85, 489–500; Chen Y. et al. (1997) “Smad8 mediates the signaling of the receptor serine kinase”, Proc. Natl Acad. Sci. USA 94, 12938–12943; Nishimura R. et al. (1998) “Smad5 and DPC4 are key molecules in mediating BMP-2-induced osteoblastic differentiation of the pluripotent mesenchymal precursor cell line C2C12”, J. Biol. Chem. 273, 1872–1879). After release from the receptor, the phosphorylated Smad proteins associate with the related protein Smad4, which acts as a shared partner. This complex translocates into the nucleus and participates in gene transcription with other transcription factors (Fig. 1). A significant advancement about the understanding of in vivo functions of BMP ligands, receptors and signaling molecules has been achieved in recent years.
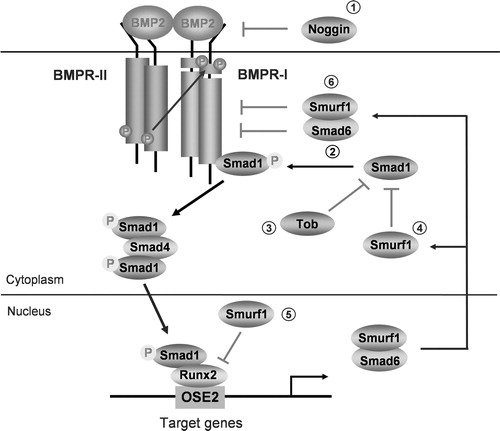
Figure 1. Figure 1 BMP signaling and its regulation. BMP signals are mediated by type I and II BMP receptors and their downstream molecules Smad1, 5 and 8. Phosphorylated Smad1, 5 and 8 proteins form a complex with Smad4 and then are translocated into the nucleus where they interact with other transcription factors, such as Runx2 in osteoblasts. BMP signaling is regulated at different molecular levels: (1) Noggin and other cystine knot-containing BMP antagonists bind with BMP-2, 4 and 7 and block BMP signaling. Over-expression of noggin in mature osteoblasts causes osteoporosis in mice (Devlin et al., 2003; Wu et al., 2003). (2) Smad6 binds type I BMP receptor and prevents Smad1, 5 and 8 to be activated (Imamura et al., 1997). Over-expression of Smad6 in chondrocytes causes delays in chondrocyte differentiation and maturation (Horiki et al., 2004). (3) Tob interacts specifically with BMP activated Smad proteins and inhibits BMP signaling. In Tob null mutant mice, BMP signaling is enhanced and bone formation is increased (Yoshida et al., 2000). (4) Smurf1 is a Hect domain E3 ubiquitin ligase. It interacts with Smad1 and 5 and mediates the degradation of these Smad proteins (Zhu et al., 1999). (5) Smurf1 also recognizes bone-specific transcription factor Runx2 and mediates Runx2 degradation (Zhao et al., 2003). (6) Smurf1 also forms a complex with Smad6, is exported from the nucleus and targeted to the type I BMP receptors for their degradation (Murakami et al., 2003). Over-expression of Smurf1 in osteoblasts inhibits postnatal bone formation in mice (Zhao et al., 2004).