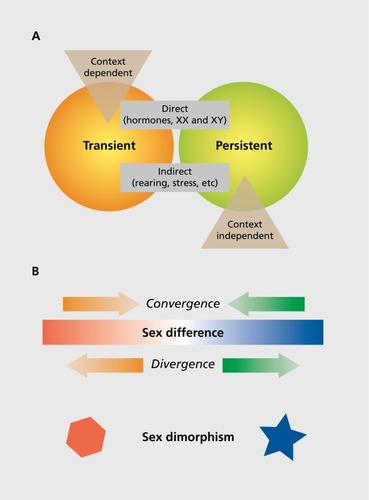
Figure 1. Conceptualizing sex differences. (A) The effects of sex on the brain organize along overlapping domains. Some sex differences are programmed early in life and persist throughout the life span, whereas others might also be programmed early but appear or disappear as a function of context (eg, age, circumstances, etc). Other sex differences are transient, which can be due to context or the result of the adult hormonal milieu, which differs in males and females and profoundly effects brain and behavior. Many sex differences that are programmed early in life in response to hormones also require hormonal activation in adulthood in order to manifest. Many sex differences can be considered direct, ie, due to hormones or sex chromosome complement, whereas others are indirect. Indirect effects include different rearing of male versus female offspring, physical constraints due to somatic sex differentiation, and in the case of humans, the impact of sex.Citation7 (B) Additional consideration in the study of sex is whether an end point is continuous or dimorphic. End points that are continuous can exhibit sex differences, meaning males and females vary on average along that continuum. Sometimes, both sexes are at the same point on the continuum at baseline but diverge in response to a challenge, such as stress. Other times, the two sexes converge on the same point from divergent beginnings. This can occur as a consequence of the cost of the reproductive profile of either sex, such as the lack of a natural induction of parenting behavior in males, which do not experience the hormonal profile of pregnancy, parturition, and lactation. Lastly, there are some end points that take on two forms, one in males and one in females, and these are considered sexually dimorphic.
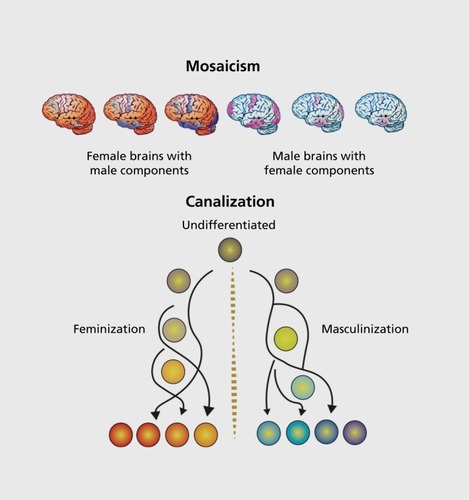
Figure 2. Mosaicism and canalization. (A) Sexual differentiation of the brain occurs independently in multiple regions. Although the same hormones (ie, testosterone and estradiol) induce sex differences, the cellular signaling pathways are unique for each region that responds. As a result, there are multiple nodes for modulation by genetic, environmental, and experiential factors, which prevent the development of a uniformly "male" versus "female" brain. Instead, the brain of any one individual is likely to be a mosaic of relative maleness, femaleness, and sameness, with the brain of no two individuals being exactly alike. This variability within the brain is countered by somatic sex, which is generally binary. (B) Canalization is a process both for constraining cell fate once differentiation begins and for maintaining species phe-notypic robustness in the face of continuous challenge and insult. This same principle can be applied to sexual differentiation of neuroanatomical end points, many of which vary along a continuum, but there is a clear separation on that continuum such that males cluster at one end and females at the other. This suggests there are biological forces that maintain a clear distinction between the sexes, for instance, by the generation of thresholds as in the case of microRNAs. But there are also forces that act to keep the sexes within range of each other as this is essential for ultimate reproductive success. Behavior is only loosely tied to neuroanatomy, and so effects of canalization are much less in evidence.
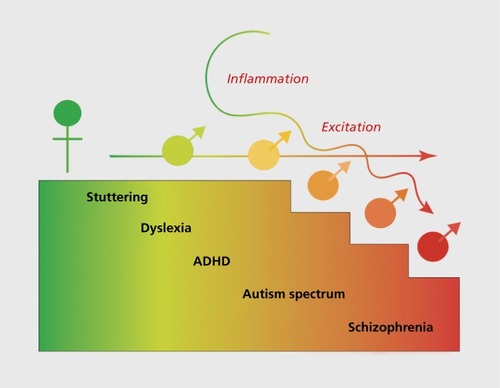
Figure 3. Mechanisms of mascuslinization put males at risk. The cellular mechanisms mediating sexual differentiation of specific regions vary, but there are unifying themes. Overall, males experience more excitation and more inflammation. In some brain regions, developing males have more microglia, and they are in a more activated state. These innate immune cells produce excess prostaglandins and other inflammatory mediators in males. Astrocytes are also more activated in developing males and are a source of increased glutamate. γ-Aminobutyric acid (GABA) is an excitatory transmitter early in development but is more excitatory and for a longer developmental duration in males than females. Two dominant features of neuropsychiatric disorders with origins in development are an imbalance of excitation and inhibition and exposure to inflammation during a critical window. Thus, males may be more susceptible to dysregulation as a consequence of the natural mediators of brain masculinization. ADHD, attention deficit/hyperactivity disorder.
JoelD.McCarthyMM.Incorporating sex as a biological variable in neuropsychiatric research: Where are we now and where should we be?Neuropsychopharmacology. 2016 Jun 22. Epub ahead of print. doi:10.1038/ npp. 2016.79.