

Abstract
Adenomyosis is a condition characterised by the invasion of endometrial tissues into the uterine myometrium, the molecular pathogenesis of which remains incompletely elucidated. Lesion profiling with next-generation sequencing (NGS) can lead to the identification of previously unanticipated causative genes and the detection of therapeutically actionable genetic changes. Using an NGS panel that included 275 cancer susceptibility genes, this study examined the occurrence and frequency of somatic mutations in adenomyotic tissue specimens collected from 17 women. Extracted DNA was enriched using targeted formalin-fixed paraffin-embedded tissue cores prior to the identification of lesion-specific variants. The results revealed that KRAS and AT-rich interactive domain 1A (ARID1A) were the two most frequently mutated genes (mutation frequencies: 24% and 12%, respectively). Notably, endometrial atypical hyperplasia did not involve adenomyotic areas. We also identified, for the first time, two potentially pathogenic mutations in the F-box/WD repeat-containing protein 7 (FBXW7) and cohesin subunit SA-2 (STAG2) genes. These findings indicate that mutations in the KRAS, ARID1A, FBXW7 and STAG2 genes may play a critical role in the pathogenesis of adenomyosis. Additional studies are needed to assess whether the utilisation of oncogenic driver mutations can inform the surveillance of patients with adenomyosis who had not undergone hysterectomy.
What is already known on this subject? Although somatic point mutations in the KRAS oncogene have been recently detected in adenomyosis, the molecular underpinnings of this condition remains incompletely elucidated. Lesion profiling with next-generation sequencing (NGS) can lead to the identification of previously unanticipated causative genes and the detection of therapeutically actionable genetic changes.
What do the results of this study add? The results of NGS revealed that KRAS and AT-rich interactive domain 1A (ARID1A) were the two most frequently mutated genes (mutation frequencies: 24% and 12%, respectively). We also identified, for the first time, two potentially pathogenic mutations in the F-box/WD repeat-containing protein 7 (FBXW7) and cohesin subunit SA-2 (STAG2) genes.
What are the implications of these findings for clinical practice and/or further research? The utilisation of oncogenic driver mutations has the potential to inform the surveillance of patients with adenomyosis who had not undergone hysterectomy.
Impact statement
Introduction
Adenomyosis, a condition that affects up to 20% of women in their reproductive age, is characterised by the invasion of endometrial tissues (glands and stroma) into the uterine myometrium (Vavilis et al. Citation1997). In general, women diagnosed with adenomyosis are multiparas between 40 and 50 years of age, although an increasing number of affected patients are nulligravidae (Li et al. Citation2018). Characteristic symptoms include dysmenorrhoea, excessive blood loss leading to anaemia, and infertility (Levgur et al. Citation2000, Sunkara and Khan Citation2012). While magnetic resonance imaging is currently considered the best diagnostic standard (Bradley et al. Citation2000), pelvic ultrasound remains the cornerstone for routine diagnosis (Basak and Saha Citation2009). Although anti-oestrogen therapy may lead to partial regression of adenomyotic foci, hysterectomy may be required in presence of severe symptomatic disease (Bergeron et al. Citation2006). While the molecular underpinnings of this condition are still incompletely elucidated (Ferenczy Citation1998), previous studies have pointed to a role for immune cells, adhesion molecules, extracellular matrix metalloproteinase and pro-inflammatory cytokines in creating the conditions for differentiation, adhesion, proliferation and survival of ectopic endometrial cells (Arnold et al. Citation2013, Lagana et al. Citation2020). In addition, the pathogenesis of adenomyosis-associated pain has been linked to an increased expression of neurotrophic factors in eutopic and ectopic endometrium (Barcena de Arellano et al. Citation2012, Citation2013, Lagana et al. Citation2018).
In recent years, broad applications of genomic characterisation to adenomyosis and related diseases (i.e. endometriosis and leiomyoma) have become mainstays of therapeutic development (Li et al. Citation2014, Anglesio et al. Citation2017, Suda et al. Citation2018, Inoue et al. Citation2019). In this scenario, next-generation sequencing (NGS) provides a powerful tool in the field of mutation analysis, allowing the extraction of high-throughput data from multiple samples per run. However, to our knowledge, no published NGS studies have reported on the occurrence of somatic mutations in cancer susceptibility genes in adenomyosis and paired normal tissues. Lesion profiling with NGS can lead to the identification of previously unanticipated causative genes and the detection of therapeutically actionable genetic changes.
Using an NGS panel that included 275 cancer susceptibility genes, this study examined the occurrence and frequency of somatic mutations in adenomyotic tissue specimens collected from 17 women. We also assessed whether the identified mutations were associated with the general characteristics of the study patients.
Methods
Patients and clinical variables
The study protocol was approved by the Institutional Review Board of the Chang Gung Memorial Hospital (reference number: 201601628B0), and written informed consent was obtained from all participants. Patients with adenomyosis who had undergone laparoscopic hysterectomy between 2006 and 2018 at the Division of Gynaecologic Oncology, Chang Gung Memorial Hospital, Taoyuan, Taiwan, were eligible for inclusion; excluded were women who received a final diagnosis of cancerous lesions concomitant with adenomyosis. The clinical variables of interest included age, body mass index (BMI), gravidity, parity, serum cancer antigen (CA) 125 levels, pathological findings and preoperative treatment approaches. The study report is consistent with the ‘Strengthening the Reporting of Observational studies in Epidemiology’ (STROBE) checklist.
Tissue specimens, DNA extraction and next-generation sequencing
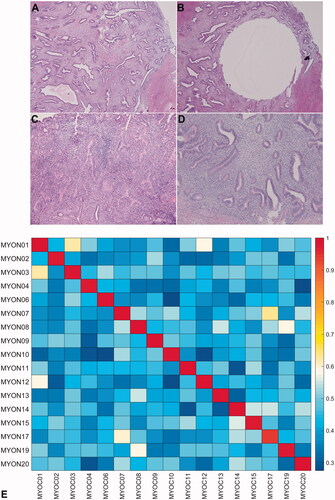
Formalin-fixed paraffin-embedded (FFPE) hysterectomy specimens – containing adenomyotic tissue and paired normal tissue samples – were obtained from all participants. As the outer half of the myometrium contained only a minor portion of adenomyotic tissue, targeted FFPE tissue cores were used to enrich adenomyotic foci (, panels A–D). Histology slides were independently reviewed by two expert pathologists (R.C.W. and L.Y.L.). Tissue DNA (40 ng) was extracted using a commercially available kit (Qiagen Inc., Valencia, CA) separately for adenomyotic lesions and matched normal tissue. DNA libraries containing barcoded universal primers (Kapa Biosystems; New England Biolabs, Ipswich, MA) were prepared, and NGS was performed using a panel (QIAseq Human Comprehensive Cancer Panel, DHS-3501Z, Qiagen, Hilden, Germany) comprising 275 cancer-related genes. Each PCR was subjected to deep-coverage sequencing (paired-end, 2 × 150 bp) in a single paired-end lane of an Illumina Next-Seq 500 platform (Illumina, San Diego, CA) following a previously described methodology (Tan et al. Citation2015).
Figure 1. Haematoxylin and eosin staining of an adenomyotic focus from patient #11 before (A) and after (B) obtaining tissue cores (H&E, ×4 objective). The superficial endometrium in patient #4 was involved by the presence of atypical hyperplasia (C), whereas the adenomyotic foci (D) were not (H&E, ×10 objective). (E) Correlation matrix (heatmap) for the 17 patients with adenomyosis included in the study.
Bioinformatic analysis
NGS data were analysed on the Qiagen Gene Globe Data Portal. After removal of the adapter sequences, high-quality reads were coupled with molecular tag (MT) sequences and aligned to the human reference genome (hg19) (Xu et al. Citation2017). Somatic mutations were detected using the Qiagen molecular tag-aware algorithm (Bhagwate et al. Citation2019). Variants in adenomyotic tissue were called when detected in at least 5% of the total reads and at least five unique reads. Variants were filtered out when their MT depth was less than 10 and their MT fraction in adenomyotic tissue was <0.05. We also excluded samples from three patients who showed a 99th percentile of the estimated minimum detectable allele fraction <0.15. The final study cohort therefore consisted of 17 patients. Somatic variants assigned to a ‘deleterious’ or ‘suspected deleterious’ category in the ClinVar data set (https://www.ncbi.nlm.nih.gov/clinvar/) were classified as pathogenic mutations according to the American College of Medical Genetics and Genomics (ACMG) recommendations (Richards et al. Citation2015).
Data analysis
Variables are summarised using descriptive statistics (means, medians, ranges, counts and frequencies). We used univariate analyses to explore the associations between the presence of specific mutations and the general characteristics of the study patients. Results are expressed as odds ratios with their 95% confidence intervals. All tests were two-sided at a 5% level of significance.
Results
Patient characteristics
Individual data of the 17 study patients (mean age: 46 years; range: 40–49 years) are presented in . Seven women (41.18%) were diagnosed with both adenomyosis and leiomyoma, whereas one (5.88%) had both adenomyosis and endometriosis. Of the four women with concomitant endometrial hyperplasia, one had evidence of atypia (#04) whereas the remaining three (#07, 11 and 17) did not. All participants had serum CA125 levels >35 U/mL.
Table 1. Clinical characteristics of patients with adenomyosis (n = 17).
Somatic mutation spectrum
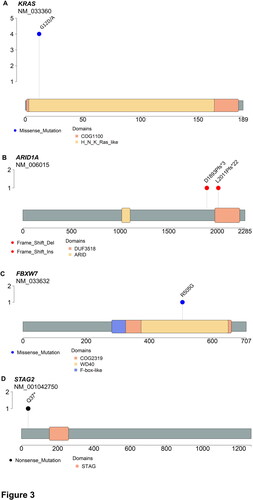
NGS was used to obtain the somatic mutation spectra of 34 tissue samples, i.e. one adenomyotic area and one normal tissue area for each of the 17 study participants. The average coverage depth was 4443.1 ± 2425.4×. Independently generated high-density SNP data were used for detecting mislabelled samples (Wang et al. Citation2016). As shown in , panel E, the results of targeted sequencing revealed a clear discrimination between paired samples obtained from the same individual (intraindividual variability) as well as different individuals (interindividual variability). An analysis of non-synonymous single nucleotide variants revealed that KRAS and ARID1A were the two most commonly mutated genes in adenomyotic tissues (mutation frequencies: 24% and 12%, respectively). Specifically, the p.Gly12Asp and the p.Gly12Ala mutations in the KRAS oncogene were identified in four adenomyotic tissue samples, whereas two frameshift mutations (p.Asp1893fs and p.Leu2011fs) in the tumour suppressor gene ARID1A were detected in two specimens (; , panels A–B). Other genes harbouring somatic mutations included AT-rich interactive domain 1B (ARID1B), adenomatous polyposis coli (APC), DICER1, deoxyribonucleic acid methyltransferases 3A (DNMT3A), epidermal growth factor receptor (EGFR), F-box/WD repeat-containing protein 7 (FBXW7), fibroblast growth factor receptor 1 (FGFR1), guanine nucleotide binding protein, alpha (GNAS), speckle-type POZ protein (SPOP), cohesin subunit SA-2 (STAG2) and telomerase reverse transcriptase (TERT) (). Of them, the p.Arg505Gly mutation in FBXW7 and the p.Gln37* mutation in STAG2 were deemed to be pathogenic (, panels C–D).
Figure 2. Heatmap of the somatic mutations identified by targeted next-generation sequencing in adenomyotic tissue specimens. AH: endometrial atypical hyperplasia; EH: endometrial hyperplasia without atypia.
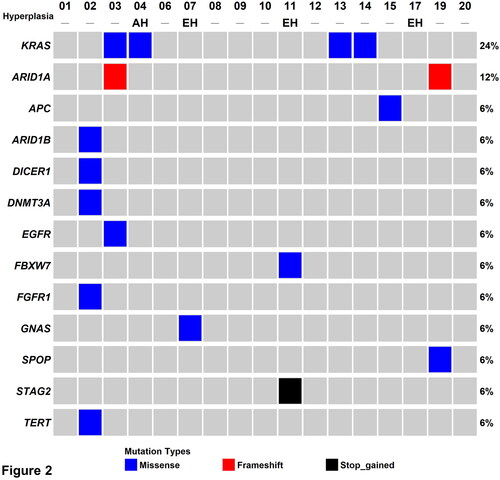
Figure 3. Diagram showing the detected mutant sites in the (A) ARID1A, (B) KRAS, (C) FBXW7 and (D) STAG2 genes.
Associations between somatic mutations in adenomyotic tissue and patient clinical characteristics
The clinical characteristics of the study participants were analysed in relation to the presence of mutations in KRAS or ARID1A (), or KRAS, ARID1A, FBXW7, or STAG2 (). However, no significant associations were observed. Of the four women with concomitant endometrial hyperplasia and adenomyosis, the only one (patient #4) who had evidence of atypical endometrial hyperplasia was a carrier of pathogenetic mutations in KRAS. Notably, atypical hyperplasia did not involve the presence of adenomyosis (, panels C,D).
Table 2. Characterisation of patients with KRAS and/or ARID1A mutation status in adenomyosis.
Table 3. Characterisation of patients with pathogenic/likely pathogenic mutation status in adenomyosis.
Discussion
The therapeutic management of adenomyosis remains challenging due to multiple factors, including poorly understood pathophysiology, heterogeneity and the lack of targetable genetic drivers. On analysing the somatic mutation profile of adenomyotic tissue obtained from 17 Taiwanese women, we identified KRAS and ARID1A as the two most frequently mutated genes (mutation frequencies: 24% and 12%, respectively). The finding related to somatic KRAS mutations is consistent with what has been reported in a previous study of adenomyosis using whole exome sequencing and deep targeted sequencing (Inoue et al. Citation2019). However, the frequency of somatic KRAS mutations reported by Inoue et al. (Citation2019) in Japan was higher (37.1%; 26/70) than that observed in the current study (24%). The exact reasons are currently unclear but may be related to different sequencing platforms and/or ethnic variability. Herein, the two somatic KRAS mutations identified in adenomyotic tissue (p.Gly12Asp and the p.Gly12Ala) were considered pathogenic – being among the most common KRAS alterations identified in cancer tissues (Krishnan et al. Citation2022). In our study, ARID1A was the second most commonly mutated gene in adenomyotic tissue. We speculate that this tumour suppressor gene may be involved in the pathogenesis of adenomyosis through its effects on several signalling pathways and the control of cell cycle (Reisman et al. Citation2009). The two ARID1A variants identified in our study were frameshift mutations (p.Asp1893fs, p.Leu2011fs) that likely caused mRNA decay and loss of protein expression (Guan et al. Citation2012). Our findings are consistent with the known role of ARID1A as an endometriosis-associated gene and its reduced protein expression in endometriotic lesions (Anglesio et al. Citation2017). Notably, ARID1A-deficient tumours are characterised by the activation of the PI3K/AKT signalling pathway.
To our knowledge, neither the p. Arg505Gly mutation in FBXW7 nor the p.Gln37* mutation in STAG2 have been previously described in adenomyosis. The FBXW7 gene – which is a critical tumour suppressor – encodes for a protein that regulates proteasome-mediated degradation of several oncoproteins (Yeh et al. Citation2018). Furthermore, FBXW7 missense mutations, especially those affecting arginine residues (as the p. Arg505Gly variant identified in our study), have a profound impact on the FBXW7 binding activity by affecting the substrate-binding interface (Hao et al. Citation2007). Interestingly, multiple FBXW7 mutations have been described in both ovarian endometriotic lesions and normal endometrium (Suda et al. Citation2018).
STAG2, a gene located on the X chromosome, is involved in sister chromatid cohesion, DNA repair, transcriptional regulation and genome topology (De Koninck and Losada Citation2016). STAG2 mutations have been identified in several human cancers – including endometrial stromal tumours (Akaev et al. Citation2021) – and have been linked to the presence of aneuploidy (Solomon et al. Citation2011). Interestingly, mutations in STAG2 can potentially be targeted by inhibitors of the DNA repair enzyme PARP (e.g. olaparib) (Bailey et al. Citation2014).
Although the detected somatic mutations were not associated with the clinical characteristics of the study patients, this result should be taken cautiously due to the small sample size. However, of the four women with concomitant adenomyosis and endometrial hyperplasia, one who had evidence of atypia – a known precursor of endometrial cancer (Russo et al. Citation2020) – was found to harbour a pathogenic KRAS mutation. These findings confirm that KRAS may drive different forms of proliferative gynaecologic lesions. While adenomyosis does not seem to affect the clinical outcomes of endometrial malignancies (Yetimalar et al. Citation2022), patients harbouring oncogenic driver mutations should be strictly followed-up for early detection of malignant transformation.
A strength of our study is that we obtained FFPE tissue cores prior to DNA isolation to precisely select adenomyotic versus normal tissue. This approach can significantly enrich the extracted DNA for the identification of lesion-specific variants of interest. Our results are expected to provide clues for understanding the molecular underpinnings of adenomyosis. In addition, the identified mutations are potentially druggable and can be used to promote the advancement of precision medicine in the treatment of this condition. Further research using well-designed clinical trials with sufficient statistical power is needed to confirm this hypothesis.
Several limitations of our study are worth noting. First, this is a single-centre investigation carried out in patients in Taiwan; therefore, the question as to whether our results are generalisable to different ethnicities remains unanswered. Second, the number of adenomyotic tissue samples was limited and replication of our findings in independent data sets is necessary to draw definitive conclusions. Lastly, the pathogenic significance of the FBXW7 p. Arg505Gly and STAG2 p.Gln37* mutations should be not be overinterpreted unless independently validated.
Conclusions
In summary, the results of targeted NGS confirmed that somatic mutations in KRAS and ARID1A may be involved in the molecular pathogenesis of adenomyosis. Furthermore, we demonstrate, for the first time, a potential pathogenic role of FBXW7 and STAG2 mutations. We suggest that patients harbouring oncogenic driver mutations who had not undergone hysterectomy should be followed-up for early detection of malignant transformation.
Author contributions
A.C. and C.J.W.: study conception and design; A.C., R.C.W., C.Y.L. and C.J.W.: drafting the manuscript or revising it critically for important intellectual content; R.C.W., C.Y.L. and C.L.T.: acquisition, analysis and interpretation of data; Y.S.L.: study supervision, critical revision of the manuscript for important intellectual content. All authors have made substantial contributions to conception and design, or acquisition of data, or analysis and interpretation of data, have been involved in drafting the manuscript or revising it critically for important intellectual content, have given final approval of the version to be published, and agree to be accountable for all aspects of the work.
Acknowledgements
We thank Jung-Erh Yang and Chu-Chun Huang for their excellent technical assistance. We are also grateful to the NGS Core Lab of the Molecular Medicine Research Centre at the Chang Gung University.
Disclosure statement
The authors declare that they have no conflicts of interest.
Data availability statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Additional information
Funding
References
- Akaev, I., Yeoh, C.C., and Rahimi, S., 2021. Update on endometrial stromal tumours of the uterus. Diagnostics, 11 (3), 429.
- Anglesio, M.S., et al., 2017. Cancer-associated mutations in endometriosis without cancer. New England Journal of Medicine, 376 (19), 1835–1848.
- Arnold, J., et al., 2013. Neuroimmunomodulatory alterations in non-lesional peritoneum close to peritoneal endometriosis. Neuroimmunomodulation, 20 (1), 9–18.
- Bailey, M.L., et al., 2014. Glioblastoma cells containing mutations in the cohesin component STAG2 are sensitive to PARP inhibition. Molecular Cancer Therapeutics, 13 (3), 724–732.
- Barcena de Arellano, M.L., et al., 2013. Evidence of neurotrophic events due to peritoneal endometriotic lesions. Cytokine, 62 (2), 253–261.
- Barcena de Arellano, M.L., et al., 2012. Eutopic endometrium from women with endometriosis does not exhibit neurotrophic properties. Journal of Neuroimmunology, 249 (1–2), 49–55.
- Basak, S. and Saha, A., 2009. Adenomyosis: still largely under-diagnosed. Journal of Obstetrics and Gynaecology, 29 (6), 533–535.
- Bergeron, C., Amant, F., and Ferenczy, A., 2006. Pathology and physiopathology of adenomyosis. Best Practice & Research. Clinical Obstetrics & Gynaecology, 20 (4), 511–521.
- Bhagwate, A.V., et al., 2019. Bioinformatics and DNA-extraction strategies to reliably detect genetic variants from FFPE breast tissue samples. BMC Genomics, 20 (1), 689.
- Bradley, L.D., Falcone, T., and Magen, A.B., 2000. Radiographic imaging techniques for the diagnosis of abnormal uterine bleeding. Obstetrics and Gynecology Clinics of North America, 27 (2), 245–276.
- De Koninck, M. and Losada, A., 2016. Cohesin mutations in cancer. Cold Spring Harbor Perspectives in Medicine, 6 (12), a026476.
- Ferenczy, A., 1998. Pathophysiology of adenomyosis. Human Reproduction Update, 4 (4), 312–322.
- Guan, B., et al., 2012. Functional analysis of in-frame indel ARID1A mutations reveals new regulatory mechanisms of its tumor suppressor functions. Neoplasia, 14 (10), 986–993.
- Hao, B., et al., 2007. Structure of a Fbw7–Skp1–cyclin E complex: multisite-phosphorylated substrate recognition by SCF ubiquitin ligases. Molecular Cell, 26 (1), 131–143.
- Inoue, S., et al., 2019. Uterine adenomyosis is an oligoclonal disorder associated with KRAS mutations. Nature Communications, 10 (1), 5785.
- Krishnan, T., et al., 2022. Targeting mutated KRAS genes to treat solid tumours. Molecular Diagnosis & Therapy, 26 (1), 39–49.
- Lagana, A.S., et al., 2020. Evaluation of M1 and M2 macrophages in ovarian endometriomas from women affected by endometriosis at different stages of the disease. Gynecological Endocrinology, 36 (5), 441–444.
- Lagana, A.S., et al., 2018. Stem cell trafficking during endometriosis: may epigenetics play a pivotal role? Reproductive Sciences, 25 (7), 978–979.
- Levgur, M., Abadi, M.A., and Tucker, A., 2000. Adenomyosis: symptoms, histology, and pregnancy terminations. Obstetrics and Gynecology, 95 (5), 688–691.
- Li, X., et al., 2014. Whole-exome sequencing of endometriosis identifies frequent alterations in genes involved in cell adhesion and chromatin-remodeling complexes. Human Molecular Genetics, 23 (22), 6008–6021.
- Li, Y.W., et al., 2018. Clinical manifestations of adenomyosis patients with or without coexisting endometriosis. Chinese Medical Journal, 131 (20), 2495–2498.
- Reisman, D., Glaros, S., and Thompson, E.A., 2009. The SWI/SNF complex and cancer. Oncogene, 28 (14), 1653–1668.
- Richards, S., et al., 2015. Standards and guidelines for the interpretation of sequence variants: a joint consensus recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genetics in Medicine, 17 (5), 405–424.
- Russo, M., et al., 2020. Mutational profile of endometrial hyperplasia and risk of progression to endometrioid adenocarcinoma. Cancer, 126 (12), 2775–2783.
- Solomon, D.A., et al., 2011. Mutational inactivation of STAG2 causes aneuploidy in human cancer. Science, 333 (6045), 1039–1043.
- Suda, K., et al., 2018. Clonal expansion and diversification of cancer-associated mutations in endometriosis and normal endometrium. Cell Reports, 24 (7), 1777–1789.
- Sunkara, S.K. and Khan, K.S., 2012. Adenomyosis and female fertility: a critical review of the evidence. Journal of Obstetrics and Gynaecology, 32 (2), 113–116.
- Tan, M.C., et al., 2015. GNAS and KRAS mutations define separate progression pathways in intraductal papillary mucinous neoplasm-associated carcinoma. Journal of the American College of Surgeons, 220 (5), 845–854.e1.
- Vavilis, D., et al., 1997. Adenomyosis at hysterectomy: prevalence and relationship to operative findings and reproductive and menstrual factors. Clinical and Experimental Obstetrics & Gynecology, 24 (1), 36–38.
- Wang, P.P., et al., 2016. BAM-matcher: a tool for rapid NGS sample matching. Bioinformatics, 32 (17), 2699–2701.
- Xu, C., et al., 2017. Detecting very low allele fraction variants using targeted DNA sequencing and a novel molecular barcode-aware variant caller. BMC Genomics, 18 (1), 5.
- Yeh, C.H., Bellon, M., and Nicot, C., 2018. FBXW7: a critical tumor suppressor of human cancers. Molecular Cancer, 17 (1), 115.
- Yetimalar, M.H., Kilic, D., and Bezircioglu, I. 2022. The impact of uterine adenomyosis on the histopathological risk factors and survival in patients with endometrial adenocarcinoma. Journal of Obstetrics and Gynaecology, 42, 1–7.