

Abstract
BTG4 arrests the cell cycle and suppresses oocyte and embryonic development. We performed a bioinformatic analysis of BTG4 expression. BTG4 expression was downregulated in breast cancer compared with normal tissues (p < .05), but the opposite was observed in cervical, endometrial and ovarian cancers (p < .05). BTG4 methylation was negatively correlated with its mRNA expression in breast, cervical and endometrial cancers (p < .05). BTG4 mRNA expression was negatively correlated with T staging and distant metastasis of breast cancer; and with tumor invasion, clinical stage, low weight and BMI, low histological grade and no diabetes in endometrial cancer but positively with T stage and non-keratinizing squamous carcinoma in endometrial cancer. BTG4 expression was negatively correlated with the survival of ovarian cancer patients (p < .05), but positively for breast, cervical and endometrial cancers (p < .05). BTG4 expression is thus a potential marker reflecting the carcinogenesis, aggressiveness and prognosis in gynecological cancers.
What is already known on this subject? Previous studies have revealed the structure and location of BTG4. BTG4 inhibit cell proliferative, promote apoptosis, induce G1 cell cycle arrest. BTG4 promotes the development of mouse embryos from cell stage 1 to 2. The methylation and biological function of BTG4 were clarified in gastric and/or colorectal cancer cells.
What do the results of this study add? BTG4 is found to closely link to reflect the carcinogenesis, histogenesis, aggressive behaviors and prognosis of gynecological cancers, and involved in ligand-receptor interaction, microtubule motor activity, dynein light chain binding, cilium organization, assembly, and movement in endometrial and ovarian cancers.
What are the implications of these finding for clinical practice and/or further research? Aberrant BTG4 mRNA expression can be employed as a marker of the tumorigenesis, histogenesis, aggressiveness and prognosis of gynecological cancers in the future practice and guide the investigation of BTG4-related signal pathways.
Impact Statement
1. Introduction
The BTG4 gene (B-cell translocation gene 4) maps to human chromosome 11q23.1, and the protein it encodes belongs to the TOB/BTG family. It has anti-proliferative effects and can induce G1 cell cycle arrest, similar to other BTG family members (Buanne et al. Citation2000). Zhang et al. (Citation2020) demonstrated the presence of Sp1 and p53 binding sites in the core promoter of BTG4 within the 388 bp region upstream of its transcription initiation site. However, both p53 overexpression and activation were shown to lead to the transcriptional upregulation of BTG4, finally blocking cell growth and apoptotic inhibition in cancer cells. BTG4 interacts with CNOT7 and CCR4-NOT, an RNA de-adenylation complex, to foster mRNA degradation, delay maternal mRNA clearance, and arrest embryonic development (Winkler Citation2010, Liu et al. Citation2016, Yu et al. Citation2016). Zheng et al. (Citation2020) identified four homozygous mutations in BTG4 (73 C>T, 1 A>G, 475- 478del, and 166 G>A) causing zygotic cleavage failure (ZCF) due to the interaction between BTG4 and CNOT7, which exhibit a recessive inheritance pattern. BTG4 transcripts are highly enriched in the ovary and accumulate in oocytes within the germinal vesicle during oocyte growth, but are not translated until meiotic maturation of fully grown oocytes, resulting in nuclear envelope rupture (Pennetier et al. Citation2005, Elis et al. Citation2008, Liu et al. Citation2016, Yu et al. Citation2016, Zheng et al. Citation2020). The similar expression patterns of pERK1/2 and BTG4 protein in mouse oocytes and embryos, and the disruptive effect of pERK1/2 inactivation on BTG4 mRNA polyadenylation and translation indicated that pERK1/2 is an upstream factor that activates BTG4 protein expression (Yu et al. Citation2016).
The development of mouse embryos lacking BTG4 is arrested at the one- to two-cell stage. Owing to this early developmental arrest, BTG4-null females produce morphologically normal oocytes but are infertile (Liu et al. Citation2016, Zheng et al. Citation2020). Mano et al. (Citation2015) found that transient BTG4 expression induced alkaline phosphatase activity in human embryonic kidney cells. In addition, BTG4 mRNA was detected in pharynx, larynx, trachea, oviduct, ovary, caput epididymis, and testis, but not in lung, intestine, or liver. BTG4 protein is present in epithelial cells of the tongue, palate, pharynx, internal nose, and trachea. Findings also revealed that both protein and mRNA levels of BTG4 were reduced by aging when comparing 4- and 48-week-old mice. Moreover, Mori et al. (Citation2011) found that BTG4 methylation could be used to distinguish colorectal cancer from age-matched control mucosa. Furthermore, Seo et al. (Citation2017) demonstrated that hinokitiol induced DNA demethylation and mRNA restoration of BTG4 via DNMT1 in colon cancer cells. Dong et al. (Citation2009) also reported that the BTG4 level was significantly reduced in gastric cancer compared with that in normal gastric tissues. BTG4 re-expression in gastric cancer then inhibited its growth, as confirmed by colony assays and a xenograft model.
In terms of cancer risk, women are particularly susceptible to breast, vulvar, vaginal, cervical, endometrial, and ovarian cancers (Börner et al. Citation2018, Králíčková et al. Citation2020, Dellino et al. Citation2021, Alves et al. Citation2022, Giannini et al. Citation2022, Khoramdad et al. Citation2022; Mulongo et al. Citation2022). Using bioinformatic databases, we aimed to clarify the clinicopathological and prognostic significance of BTG4 mRNA expression and signal pathways related to such expression in breast, cervical, endometrial, and ovarian cancers using Oncomine, TCGA, Xiantao, UALCAN, and Kaplan–Meier plotter datasets as shown in Supplementary Figure 1.
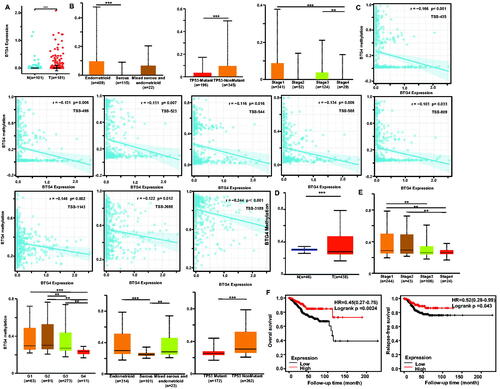
Figure 1. Clinicopathological and prognostic significance of BTG4 mRNA expression in endometrial cancer. According to Xiantao (A) data, BTG4 mRNA was upregulated in endometrial cancer compared with the level in normal tissue (p < .05). BTG4 mRNA expression was negatively associated with serous subtype, TP53 mutation, and low TNM stage of endometrial cancer according to UALCAN (B, p <.05). The negative relationship between BTG4 mRNA expression and methylation was analyzed in endometrial cancer using Xiantao database (C, p < .05). Its methylation was lower in endometrial cancer than in normal tissues (D, p < .05), and its correlations with N staging, clinicopathological stage, histological grade, histological subtype, and TP53 mutation were analyzed by UALCAN (E). There was a positive correlation between BTG4 mRNA expression and overall relapse-free survival rate of the patients with endometrial cancer (H, p <.05). Note: N: normal tissue; T: tumor; HR: hazard ratio.
2. Material and methods
2.1. Oncomine database analysis
The gene expression level of BTG4 was analyzed using OncomineTM 4.5 (www.oncomine.org), a cancer microarray database and web-based data mining platform for making new discoveries from genome-wide expression analyses. We analyzed the differences in BTG4 mRNA levels between normal and cancer tissues. All data were log-transformed, the median was centered per array, and the standard deviation was normalized to one per array.
2.2. TCGA database analysis
The expression data (RNA-seqV2) and clinicopathological data of breast cancer patients (n = 1093) were downloaded from the Cancer Genome Atlas (TCGA, https://cancergenome.nih.gov/) database by TCGA-assembler in R software (Szász et al. Citation2016). We integrated the raw data, analyzed BTG4 mRNA expression in the cancer, and compared it with clinicopathological and prognostic data of the cancer patients. The means were compared using Student’s t-test. Kaplan–Meier survival plots were generated with survival curves compared by log-rank statistics. Cox’s proportional hazards model was employed for multivariate analysis. Two-sided p < .05 was considered statistically significant. SPSS 17.0 software was employed to analyze all of the data.
2.3. KM plotter analysis
The prognostic significance of BTG4 mRNA (ID: 220766_at) was also analyzed in breast, cervical, endometrial and ovarian cancers using Kaplan–Meier plotter (http://kmplot.com) (Zhu et al. Citation2014).
2.4. UALCAN analysis
The expression and methylation of the BTG4 gene were analyzed using the UALCAN database (http://ualcan.path.uab.edu). They were also compared with the clinicopathological and prognostic parameters of breast, cervical, endometrial and ovarian cancers.
2.5. Xiantao analysis
The expression and methylation of the BTG4 gene were analyzed using the Xiantao platform (https://www.xiantao.love/). Additionally, we found the differential and related genes of BTG4 using Xiantao platform. The differentially expressed genes were used to construct a PPI network and the important hub genes were selected out. These genes were subjected to KEGG analysis in order to construct signal pathways.
3. Results
3.1. Clinicopathological and prognostic significance of BTG4 mRNA expression in breast cancer
According to the Xiantao (Supplementary Figure 2A) and UALCAN databases (Supplementary Figure 2B), BTG4 expression was lower in breast cancer than in normal tissues (p < .05). TCGA database showed that BTG4 expression was negatively correlated with T staging and distant metastasis of breast cancer patients (Supplementary Figure 2C, p < .05). In UALCAN, luminal subtype showed higher BTG4 expression than triple-negative breast cancer (TNBC, Supplementary Figure 2D, p < .05). There was also a negative correlation between BTG4 mRNA and methylation (cg09170311, cg18515591, and cg22806002) according to Xiantao (Supplementary Figure 2E, p < .05). Moreover, BTG4 methylation was higher in breast cancer than in normal tissues (Supplementary Figure 2F, p < .05), higher in N2 than in N0, higher in stages 2 and 3 than in stage 1, and higher in stage 3 than in stage 2 cancer patients (Supplementary Figure 2G, p < .05).
Our findings obtained using Kaplan–Meier plotter revealed that all cancer patients with high BTG4 expression showed longer relapse-free survival than those with its low expression (Supplementary Figure 1H, p < .05). There appeared to be positive relationships between BTG4 expression and the relapse-free survival rate of patients with luminal-A, or luminal-B and basic-like1 cancers (Supplementary Figure 1H, p < .05).
3.2. Clinicopathological and prognostic significance of BTG4 mRNA expression in cervical cancer
We used the Xiantao database to perform bioinformatic analysis and found that BTG4 expression was higher in cervical cancer than in normal tissue (Supplementary Figure 3A, p < .05). As shown in , BTG4 mRNA expression was positively associated with T stage squamous cell carcinoma, and non-keratinizing squamous cell carcinoma (p < .05). The Xiantao database showed that BTG4 mRNA expression was negatively associated with the methylation of BTG4 promoter sites (TS-3, −208, 435, −523, −588, −809, −968, −1143, −2579, and −3189; Supplementary Figure 3B, p < .05). BTG4 methylation was also lower in cervical cancer than in normal tissue (Supplementary Figure 3C, p < .05), in squamous carcinoma than in mucinous and endocervical adenocarcinoma, in adenosquamous than in endocervical carcinoma, and in African-American than in Caucasian cervical cancer patients (Supplementary Figure 3D, p < .05). Our findings obtained using Kaplan–Meier plotter revealed that higher BTG4 expression was positively correlated with overall and relapse-free survival rates of Grade 3 cancer patients (Supplementary Figure 3E, p < .05).
Figure 2. Clinicopathological and prognostic significance of BTG4 mRNA expression in ovarian cancer. Hendrix’s datasets were employed for bioinformatic analysis to observe BTG4 mRNA expression in ovarian cancer. BTG4 mRNA expression was lower in ovary than in clear cell adenocarcinoma (CCA), endometrioid (Endo), mucinous (Muc), and serous (Sero) adenocarcinoma (A, p <.05), in agreement with the findings from Xiantao (B, p <.05). The correlation was analyzed between BTG4 mRNA expression and overall or post-progression survival rate of the patients with breast cancer, even stratified by different clinicopathological parameters (C, p <.05). Note: N: normal tissue; T: tumor; HR: hazard ratio.
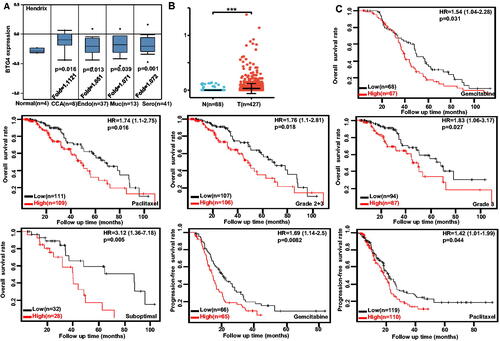
Table 1. Relationship between BTG4 expression and clinicopathological parameters of cervical cancer.
3.3. Clinicopathological and prognostic significance of BTG4 mRNA expression in endometrial cancer
According to the Xiantao database, BTG4 expression was higher in endometrial cancer than in normal tissues (, p < .05). BTG4 expression was also higher in endometrioid carcinoma than in serous adenocarcinoma, in non-TP53-mutant than in TP53-mutant cancer patients, and in stage 1 and stage 3 than in stage 4 cancer patients (, p < .05). As summarized in , BTG4 mRNA expression was negatively associated with tumor invasion, clinical stage, low weight and BMI, serous subtype, low histological grade, and no diabetes (p < .05). There was also a negative correlation between BTG4 mRNA and its methylation (TS-435, −496, −523, −544, −588, −809, −968, −1143, −2698, and −3189), as revealed by Xiantao (, p < .05). BTG4 methylation was higher in endometrial normal tissue than in cancer tissue (, p < .05); in stage 1 than in stage 3 and stage 4 cancer patients; in stage 2 than in stage 4 cancer patients; in Grade 1, Grade 2, and Grade 3 cancer patients than in Grade 4 ones; in grade 2 than in grade 3 cancer patients; in endometrioid and mixed serous and endometrioid carcinoma than in serous carcinoma; and in non-TP53-mutant than in TP53-mutant cancer patients (, p < .05). According to Kaplan–Meier plotter, we found that higher BTG4 expression was positively correlated with overall and relapse-free survival rates of all cancer patients (, p < .05).
Table 2. Relationship between BTG4 expression and clinicopathological parameters of endometrial cancer.
3.4. Clinicopathological and prognostic significance of BTG4 mRNA expression in ovarian cancer
We performed a bioinformatic analysis of BTG4 expression in ovarian cancer using Hendrix’s datasets. BTG4 had lower expression in normal ovary than in clear cell, endometrioid, mucinous, and serous adenocarcinomas (, p < .05), consistent with the finding from Xiantao (, p < .05). The data from Kaplan–Meier plotter showed a negative relationship between BTG4 expression and either overall or progression-free survival rate of ovarian cancer patients, treated with paclitaxel or gemcitabine (, p < .05). A negative correlation between BTG4 expression and overall survival rate was observed in the patients with grade 2 + 3, grade 2, or suboptimal ovarian cancer (, p < .05).
3.5. Relationship between BTG4 mRNA expression and infiltrating immune cells in gynecological cancers
According to Xiantao, BTG4 mRNA expression was positively related to the infiltration of eosinophils, mast cells, and NK CD56bright cells in breast cancer (Supplementary Figure 4A, p < .05). Meanwhile, it was negatively related to the infiltration of T cells, NK CD56bright cells, TFH, CD8+ T cells, cytotoxic cells, and Th17 cells in cervical cancer (Supplementary Figure 4B, p < .05). It was also positively related to the infiltration of NK CD56bright cells, eosinophils, and mast cells, but negatively related to macrophages, NK CD56dim, Treg, Tem, Tgd, Th1, aDC, DC, and Th1 cells in endometrial cancer (Supplementary Figure 4C, p < .05). Moreover, it was positively related to the infiltration of NK CD56bright cells, but negatively related to Th1 cells, macrophages, cytotoxic cells, NK CD56dim cells, T cells, Tem, mast cells, B cells, CD8 T cells, and Idc in ovarian cancer (Supplementary Figure 4D, p < .05).
3.6. Btg4-related genes and pathways in gynecological cancers
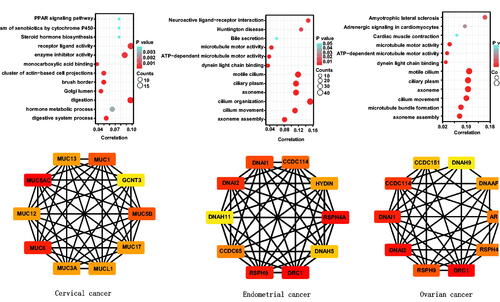
On the Xiantao platform, we discovered the genes that differed in expression between the groups with low and high expression of BTG4 mRNA in gynecological cancers. KEGG analysis showed that the top signal pathways related to BTG4 included hormone metabolism and biosynthesis, PPAR signal pathway, xenobiotic metabolism, receptor ligand activity, enzyme inhibitor activity, monocarboxylic acid binding, brush border, and Golgi lumen in cervical cancer; and ligand-receptor interaction, microtubule motor activity, dynein light chain binding, and cilium organization and movement in endometrial and ovarian cancers (). In addition, STRING was used to identify the PPI pairs and Cytoscape was used to identify the top 10 nodes ranked by the level of connectivity (). The top hub genes mainly included mucins in cervical cancer; and outer dynein arm docking complex, dynein axonemal intermediate or heavy chain, radial spoke head component, and dynein regulatory complex in endometrial and ovarian cancers.
The genes shown to be correlated with BTG4 in cancers by the Xiantao database were analyzed and subjected to KEGG analysis (Supplementary Figure 5). The BTG4-correlated genes were found to be involved in chemical carcinogenesis, drug metabolism by cytochrome P450, growth factor binding, collagen-containing extracellular matrix, cornified envelope, basement membrane, and keratinocyte differentiation for cervical cancer; and microtubule motor activity, dynein light chain binding, and ciliary part, as well as cilium organization, assembly, and movement for endometrial and ovarian cancers.
Figure 3. Differentially-expressed genes and signal pathways related to BTG4 in gynecological cancers. The genes differentially expressed in association with BTG4 were subjected to signal pathway analysis using KEGG (A). STRING was used to construct a protein-protein interaction network of genes differentially expressed in association with BTG4 in cancers, and Cytoscape was employed to identify the top 10 hub nodes ranked by the level of connectivity (B).
4. Discussion
Räty et al. (Citation2011) identified BTG4 as a potential candidate for oocyte developmental competence, as a secreted acidic protein rich in cysteine for cumulus cell expansion. Moreover, Liu et al. (Citation2021) reported that a novel homozygous missense variant in BTG4 causes zygotic cleavage failure and female infertility. Meanwhile, Chu et al. (Citation2012) found that BTG4 had a higher mRNA level in oocytes treated with FSH than in those produced through the natural cycle. Furthermore, Pasternak et al. (Citation2016) demonstrated that the BTG4-CAF1 complex safeguarded metaphase II arrest in mammalian eggs by deadenylating maternal mRNAs. BTG4-depleted eggs progressed into anaphase II spontaneously before fertilization. BTG4 prevented the progression into anaphase by ensuring that the anaphase-promoting complex/cyclosome (APC/C) was completely inhibited during the arrest. These findings indicate that BTG4 may block the cycle progression of oocytes or eggs.
CpG island methylation was reported to be associated with transcriptional silencing of BTG4, and BTG4 overexpression suppressed colony formation of colorectal cancer cells (Toyota et al. Citation2008). BTG4 could also contribute to the pathogenesis of chronic lymphocytic leukemia following inactivation by haploinsufficiency (Auer et al.Citation2005). In the present study, BTG4 mRNA expression was downregulated in breast cancers compared with that in normal tissue, but upregulated in cervical, endometrial and ovarian cancers, indicating that aberrant expression of BTG4 mRNA is closely linked to carcinogenesis. These conflicting findings might be due to tissue specificity or the difference in embryonic layers from which the cancerous tissue was derived (breast from ectoderm; ovary, cervix, and uterus from entoderm). Moreover, a negative correlation was identified between mRNA expression and promoter methylation of BTG4 in breast, cervical, and endometrial cancers, indicating the aberrant BTG4 expression might be attributable to its promoter methylation, which will be analyzed in future study.
BTG4 mRNA expression was negatively correlated with T staging and distant metastasis of breast cancer patients; and with tumor invasion, clinical stage, low weight and BMI, low histological grade, and no diabetes in endometrial cancer. However, it was positively correlated with T stage squamous cell carcinoma, and non-keratinizing squamous cell carcinoma in endometrial cancer, indicating that it is a potential marker of aggressiveness of gynecological cancers. In addition, BTG4 expression was highest in endometrioid adenocarcinoma and cervical endocervical squamous cell carcinoma, suggesting the histogenic role of BTG4 in cervical and endometrial cancers. Finally, BTG4 expression was positively related to survival rates of breast, cervical, and endometrial cancer, but was negatively related to ovarian cancer survival. Therefore, caution should be taken when employing BTG4 expression as a prognostic marker of cancers.
BTG4 has been reported to arrest egg cells in metaphase II and to suppress the proliferation of colorectal cancer cells (Toyota et al. Citation2008, Pasternak et al. Citation2016). Our bioinformatic analysis showed that BTG4 might be involved in hormone metabolism and biosynthesis, the PPAR signal pathway, drug metabolism, receptor ligand activity, collagen-containing extracellular matrix, basement membrane, monocarboxylic acid binding, brush border, and Golgi lumen in cervical cancer; along with ligand-receptor interaction, microtubule motor activity, dynein light chain binding, and cilium organization, assembly, and movement in endometrial and ovarian cancers. These findings provide novel clues about the roles and molecular mechanisms by which BTG4 acts in the tumorigenesis and progression of gynecological cancers, which should be investigated in future work.
Neoadjuvant chemotherapy and primary debulking surgery reportedly have similar efficacy for ovarian cancer, but they differ in their adverse effects (Fagotti et al. Citation2020). The majority of ovarian cancer patients achieved complete remission after the first cycle of chemotherapy, and subsequently presented a relapse due to remaining drug-resistant cells with genetic and epigenetic alterations (Laganà et al. Citation2015). Additionally, the suppression of either immune activation or surveillance has been found to correlate with the promotion of drug resistance (Laganà et al. Citation2016, Li et al. Citation2022). In the present study, we found that BTG4 mRNA expression was closely linked to the infiltration of immune cells, indicating BTG4’s potential role in immune surveillance and the response to chemotherapy of gynecological cancers, which will be further investigated in future work.
BTG4 exerts anti-proliferative effects by arresting the cell cycle and can inhibit oocyte and embryonic development and carcinogenesis. To improve our understanding of the anti-tumor effect of oocyte-arresting BTG4 and identify new cancer diagnostic biomarkers, we firstly employed bioinformatics to clarify the clinicopathological and prognostic significance of BTG4 mRNA expression in gynecological cancers and its related signal pathways. However, this study had the limitation that the results from the Oncomine, TCGA, Xiantao, UALCAN, and Kaplan–Meier plotter datasets were not validated using real-time RT-PCR, even after laser capture dissection, because of insufficient specificity of cDNA chip or RNA sequencing. The association between BTG4 mRNA expression and promoter methylation, chemoresistance, or immune surveillance should thus be studied in future work.
In summary, aberrant BTG4 mRNA expression is closely linked to carcinogenesis and can be employed as a marker of the aggressiveness of breast cancer and histogenesis in gynecological cancers. BTG4 mRNA expression level in cancer and its prognostic significance vary depending on the cancer type.
Author contributions
All authors approved the final manuscript as submitted and agree to be accountable for all aspects of the work. Hua-chuan Zheng: Project development, Study design, Manuscript writing, Data analysis and Interpreted the data. Hang Xue: Revised the manuscript and Interpreted the data. Cong-yu Zhang: Project development, Revised the manuscript and Interpreted the data. Rui Zhang: Critically reviewed the manuscript.
Supplemental Material
Download Zip (1.8 MB)Disclosure statement
No potential conflict of interest was reported by the author(s).
Data availability statement
The datasets generated and analyzed in the present study are available from the corresponding author on reasonable request.
Additional information
Funding
References
- Alves Pinto de Andrade, D., et al., 2022. Reassessment of low- and intermediate-risk endometrial cancer reports by gynecological pathologists increases risk classification without impacting outcome. The European Journal of Obstetrics & Gynecology and Reproductive Biology, 278, 172–176.
- Auer, R.L., et al., 2005. Identification of a potential role for POU2AF1 and BTG4 in the deletion of 11q23 in chronic lymphocytic leukemia. Genes, Chromosomes & Cancer, 43 (1), 1–10.
- Börner, C., et al., 2018. Pain mechanisms in peritoneal diseases might be partially regulated by estrogen. Reproductive Sciences, 25 (3), 424–434.
- Buanne, P., et al., 2000. Cloning of PC3B, a novel member of the PC3/BTG/TOB family of growth inhibitory genes, highly expressed in the olfactory epithelium. Genomics, 68 (3), 253–263.
- Chu, T., et al., 2012. Effect of ovarian stimulation on oocyte gene expression in cattle. Theriogenology, 77 (9), 1928–1938.
- Dellino, M., et al., 2021. A strengthening the reporting of observational studies in epidemiology (STROBE): are HE4 and CA 125 suitable to detect a Paget disease of the vulva? Medicine, 100 (5), e24485.
- Dong, W., et al., 2009. Frequent promoter hypermethylation and transcriptional downregulation of BTG4 gene in gastric cancer. Biochemical and Biophysical Research Communications, 387 (1), 132–138.
- Elis, S., et al., 2008. Search for the genes involved in oocyte maturation and early embryo development in the hen. BMC Genomics., 9, 110.
- Fagotti, A., et al., 2020. Randomized trial of primary debulking surgery versus neoadjuvant chemotherapy for advanced epithelial ovarian cancer (SCORPION-NCT01461850). International Journal of Gynecological Cancer: Official Journal of the International Gynecological Cancer Society, 30 (11), 1657–1664.
- Giannini, A., et al., 2022. Advances on prevention and screening of gynecologic tumors, are we stepping forward? Healthcare, 10 (9), 1605.
- Khoramdad, M., et al., 2022. Breast cancer risk factors in Iranian women: a systematic review and meta-analysis of matched case-control studies. European Journal of Medical Research, 27 (1), 311.
- Králíčková, M., et al., 2020. Endometriosis and risk of ovarian cancer: what do we know? Archives of Gynecology and Obstetrics, 301 (1), 1–10.
- Laganà, A.S., et al., 2015. Cytogenetic analysis of epithelial ovarian cancer’s stem cells: an overview on new diagnostic and therapeutic perspectives. European Journal of Gynaecological Oncology, 36 (5), 495–505.
- Laganà, A.S., et al., 2016. Epithelial ovarian cancer inherent resistance: may the pleiotropic interaction between reduced immunosurveillance and drug-resistant cells play a key role? Gynecologic Oncology Reports, 18, 57–58.
- Li, Y., et al., 2022. MSLN correlates with immune infiltration and chemoresistance as a prognostic biomarker in ovarian cancer. Frontiers in Oncology, 12, 830570.
- Liu, R., et al., 2021. A novel homozygous missense variant in BTG4 causes zygotic cleavage failure and female infertility. Journal of Assisted Reproduction and Genetics, 38 (12), 3261–3266.
- Liu, Y., et al., 2016. BTG4 is a key regulator for maternal mRNA clearance during mouse early embryogenesis. Journal of Molecular Cell Biology, 8 (4), 366–368.
- Mano, H., et al., 2015. Age-related decrease of IF5/BTG4 in oral and respiratory cavities in mice. Bioscience, Biotechnology, and Biochemistry, 79 (6), 960–968.
- Mori, Y., et al., 2011. Novel candidate colorectal cancer biomarkers identified by methylation microarray-based scanning. Endocrine-Related Cancer, 18 (4), 465–478.
- Mulongo, M., et al., 2022. Prevention of cervical cancer in low-resource African settings. Obstetrics and Gynecology Clinics of North America, 49 (4), 771–781.
- Pasternak, M., et al., 2016. The BTG4 and CAF1 complex prevents the spontaneous activation of eggs by deadenylating maternal mRNAs. Open Biology, 6 (9), 160184.
- Pennetier, S., et al., 2005. Genes preferentially expressed in bovine oocytes revealed by subtractive and suppressive hybridization. Biology of Reproduction, 73 (4), 713–720.
- Räty, M., et al., 2011. In vitro maturation supplements affect developmental competence of bovine cumulus-oocyte complexes and embryo quality after vitrification. Cryobiology, 63 (3), 245–255.
- Seo, J.S., et al., 2017. Hinokitiol induces DNA demethylation via DNMT1 and UHRF1 inhibition in colon cancer cells. BMC Molecular and Cell Biology, 18 (1), 14.
- Szász, A.M., et al., 2016. Cross-validation of survival associated biomarkers in gastric cancer using transcriptomic data of 1,065 patients. Oncotarget, 7 (31), 49322–49333.
- Toyota, M., et al., 2008. Epigenetic silencing of microRNA-34b/c and B-cell translocation gene 4 is associated with CpG island methylation in colorectal cancer. Cancer Research, 68 (11), 4123–4132.
- Winkler, G.S., 2010. The mammalian anti-proliferative BTG/Tob protein family. Journal of Cellular Physiology, 222 (1), 66–72.
- Yu, C., et al., 2016. BTG4 is a meiotic cell cycle-coupled maternal-zygotic-transition licensing factor in oocytes. Nature Structural & Molecular Biology, 23 (5), 387–394.
- Zhang, N., et al., 2020. BTG4 is a novel p53 target gene that inhibits cell growth and induces apoptosis. Genes (Basel), 11 (2), 217.
- Zheng, W., et al., 2020. Homozygous mutations in BTG4 cause zygotic cleavage failure and female infertility. American Journal of Human Genetics, 107 (1), 24–33.
- Zhu, Y., et al., 2014. TCGA-assembler: open-source software for retrieving and processing TCGA data. Nature Methods, 11 (6), 599–600.