

Abstract
YKL-40 is a secreted glycoprotein that can promote invasion, angiogenesis and inhibit apoptosis, and was highly expressed in a variety of tumours. In this paper, we investigated the impacts of YKL-40 on proliferation and invasion in HTR-8/SVneo cells during placenta accreta spectrum disorders (PAS) development. The levels of YKL-40 protein in late-pregnant placental tissue were detected using immunohistochemistry and Western blotting, and gene expression using reverse transcription-quantitative polymerase chain reaction (RT-qPCR). The proliferation, migration, invasion and apoptosis abilities of HTR-8/SVneo cells were detected by cell counting kit-8 (CCK-8), Transwell, scratch assay, and flow cytometry, respectively. Our current results showed that YKL-40 was significantly increased in the PAS group compared to the normal control group (P < 0.01). Biological function experiments showed that YKL-40 significantly promoted the proliferation, migration and invasion of HTR-8/SVneo cells, and inhibited cell apoptosis. Knockdown of YKL-40 inhibited the activation of Akt/MMP9 signalling in trophoblast cells. These data suggested that YKL-40 might be involved in the progression of PAS, which may be attributed to the regulation of Akt/MMP9 signalling pathway.
What is already known on this subject? YKL-40 is a secretory glycoprotein that can promote invasion, angiogenesis, and inhibit apoptosis and was highly expressed in a variety of tumours. Trophoblast cells resemble tumour cells in their migration and invasion.
What the results of this study add? YKL-40 expression was significantly up-regulated in PAS. CCK-8 assays showed that YKL-40 remarkably enhanced the viability of HTR-8/SVneo cells. Scratch and Transwell assays demonstrated that YKL-40 significantly promoted the migration and invasion of HTR-8/SVneo cells. Additionally, YKL-40 attenuated apoptosis in HTR-8/SVneo cells.
What the implications are of these findings for clinical practice and/or further research? Akt/MMP9 was involved in the regulation of YKL-40 on trophoblast invasion, which may provide theoretical basis and new ideas for the drug blocking intervention of placenta accreta.
IMPACT STATEMENT
Introduction
Placenta accreta spectrum (PAS) is the abnormal attachment and invasion of the chorionic villi in the myometrium. Pathologically, it can be classified into three categories: placenta accreta, increta, and percreta (Jauniaux et al. Citation2018). A recent study showed that the overall rate of PAS in the United States was 3.7‰ delivery-related discharges (Mogos et al. Citation2016). The primary cause of PAS is caesarean delivery. In recent years, the incidence of placenta previa and PAS has increased dramatically with the increase in caesarean delivery rates. In addition, postpartum haemorrhage and obstetric hysterectomy have increased rapidly due to the occurrence of placenta accreta and placenta previa (Pan et al. Citation2015, Weiniger Citation2020, OğlakÖlmez and Tunç Citation2022). The pathogenesis of PAS could be attributed to the defects in uterine metaplasm, excessive invasion of extravillous trophoblast cells, and abnormal angiogenesis (Long et al. Citation2020). Moreover, the proliferation, invasion and migration of trophoblast cells was similar to tumour cells (Zhang et al. Citation2020). Hence, further exploration of the pathogenesis of abnormal trophoblast invasion in PAS will provide more help for the diagnosis and treatment of PAS in the clinic.
Chitinase 3-like-1 (CHI3L1, also known as YKL-40 in humans and BRP-39 in mice), has a molecular weight of 40 kDa (Johansen et al. Citation1992). It is a glycoprotein secreted by chondroid, endothelial, tumour, and various inflammatory cells. It regulates vascular endothelial growth factor (VEGF) and plays a critical role in inflammation and angiogenesis (Chen et al. Citation2021). YKL-40 was overexpressed in tissues of patients with different types of tumours such as bladder cancer, colorectal cancer, glioma, ovarian cancer, and prostate cancer (Larionova et al. Citation2020, Hao et al. Citation2021, Liu et al. Citation2021, OhPyo and Son, Citation2021). It was also expressed abnormally as an inflammatory factor in asthma, rheumatoid arthritis, preeclampsia and kidney disease (Madazli et al. Citation2008, Yeo et al. Citation2019, Guvenir et al. Citation2021, Puthumana et al. Citation2021). Several studies have demonstrated that YKL-40 promoted cancer cell invasion, migration, and inhibition of apoptosis in various cancers (Hao et al. Citation2021). Currently, Gözükara et al. (Citation2017) concluded that YKL-40 is overexpressed in placental tissues with PAS. Nonetheless, its mechanism and biological function has not been clarified. Several studies have shown that PI3K/Akt was involved in multiple physiological and pathological processes, including cell proliferation, cell cycle, apoptosis, inflammation, chemical damage, and tumour progression (Wang et al. Citation2020). In addition, the epithelial-to-mesenchymal transition (EMT), migration, and invasion of trophoblasts can be promoted by activating the PI3K/Akt/Erk1/2 signalling pathway (Ding et al. Citation2021). Matrix metalloproteinases (MMPs) are a set of zinc endopeptidases vital for the decomposition of extracellular matrix (ECM) components and basement membrane (BM) (Jiang and Li Citation2021) and play a crucial role in various physiological and pathological processes, including morphogenesis, angiogenesis, inflammation, cancer invasion, and metastasis (Raeeszadeh-Sarmazdeh et al. Citation2020). Nevertheless, the potential relationship between YKL-40 and the Akt/MMP9 signalling pathway has not been explored in PAS.
In the present study, we found that YKL-40 mRNA and protein expression was up-regulated in placental tissues of PAS patients compared with the normal controls. The first extravillous cytotrophoblast (evCTB) cell line was established by Graham et al. (Citation1993) and named HTR-8/SVneo. It was generated by freshly isolated evCTB from the first-trimester placenta. HTR-8/SVneo cells are often used to detect the invasive and migratory ability of trophoblasts. To further elucidate the functional role of YKL-40 in the pathogenesis of PAS, we knocked down and overexpressed YKL-40 in vitro to investigate that YKL-40 plays a vital role in PAS by activating Akt/MMP9 pathways with human trophoblastic cell line HTR-8/SVneo.
Materials and methods
Patients
Placental tissues were obtained at the time of caesarean section from the Department of Obstetrics, Hebei General Hospital (Shijiazhuang, China) between June 2018 and December 2021. The study was approved by the Ethics Committee of the Hebei General Hospital, and all the pregnant women or their relatives provided written informed consent. In this study, a total of 25 cases of pregnant women (age 31.61 ± 3.37 years) with PAS, including 13 cases of placenta increta, and 12 of placenta percreta; meanwhile, 22 cases healthy pregnant women (age 32.14 ± 4.36 years) due to the caesarean section scar were included as the normal control group.
PAS was diagnosed during pregnancy by ultrasound and magnetic resonance imaging (MRI) and confirmed intraoperatively and pathologically. Inclusion conditions: 1) placenta increta or placenta percreta; 2) natural conception, singleton pregnancy; 3) gestational age ≥28 weeks. Exclusion criteria: 1) multifetal gestations; 2) gestational hypertension, gestational diabetes mellitus; 3) BMI ≥28; 4) other pregnancy complications.
Collection and preservation of placental samples
Placental tissues were collected within 3 min after caesarean sections, including the implanted portion of the uterus. It was divided into two parts: one was immediately fixed in formalin and embedded in paraffin after 24 h for immunohistochemical, while the other was flash-frozen in liquid nitrogen and stored at −80 °C refrigerator until further analysis.
Cell culture
Human HTR-8/SVneo trophoblast cell line was purchased from the ZhongQiao XinZhou Biotech (Shanghai, China). Cells were maintained in RPMI1640 medium (Gibco, USA) supplemented with 10% foetal bovine serum (Gibco, Australia) and 1% penicillin/streptomycin at 37 °C with 5% CO2 and 95% air.
Immunohistochemical staining of placental tissue
5-μm-thick sections were dewaxed in xylene (I and II) and gradient of anhydrous ethanol (100%,100%, 95%, 90%, 80%, and 70%). Then, the sections were placed in Tris/EDTA buffer pH 9.0 for 20mins for heat mediated antigen retrieval before incubation with the primary antibodies (YKL-40, 1:100, ab255297, Abcam) at 4 °C and overnight. Subsequently, the slides were incubated the membranes were incubated with horseradish peroxidase (HRP)-labeled secondary antibody (PV-6000, ZSGB-BIO, China) at room temperature for 1 h and visualised using the chromogenic 3,3′-diaminoenzidine, followed by re-staining with haematoxylin and dehydration with a gradient of anhydrous ethanol (70%, 80%, 90%, 95%, and 100%).
YKL-40 expression was measured by brown staining of the cell cytosol or cell membrane. Five fields (×200) were selected randomly to estimate the staining intensity grade: negative (−), weak-positive (+), medium-positive (++), and strong-positive (+++).
Total RNA extraction and RT-qPCR
Total RNA was extracted from the placental tissues and HTR-8/SVneo cells using the RNeasy Mini Kit (TIANGEN, Beijing, China). The concentration of RNA was measured on an Ultraviolet spectrophotometer (Thermo, USA). cDNA was transcribed from 2 μg RNA using the Fast Quant RT Kit (with gDNase) (TIANGEN, Beijing, China). RT-qPCR was carried out to evaluate the expression of YKL-40 using Super Real PreMix Plus (SYBR Green) (TIANGEN, Beijing, China) on an ABI PRISM 7500 sequence detector (Applied Biosystems, USA), according to the manufacturer’s instructions. GAPDH was used as an internal control to calculate the relative expression. The 2−ΔΔCt method was used to calculate the relative expression levels. Each sample was replicated at least three times.
All primers were synthesised by Youbao Biotech (Changsha, China), the sequences were as follows:
YKL-40 (CHI3L1): Forward: 5′-GTGAAGGCGTCTCAAACAGG-3′, Reverse: 5′-GAAGCGGTCAAGGGCATCT-3′.
GAPDH: Forward: 5′-GGAAGCTTGTCATCAATGGAAATC-3′, Reverse: 5′-TGATGACCCTTTTGGCTCC-3′
Extraction of total protein and Western blot
Total protein was extracted from the placental tissues and cells using RIPA buffer containing 50 μg/500 mL or each well of the 6‐well plate/200 μL protease inhibitors (Solarbio, Beijing China) according to the manufacturer’s instructions. Protein samples were resolved by 10% SDS-PAGE and transferred to polyvinylidene fluoride (PVDF) membrane. Then, the membranes were blocked with 5% skim milk in Tris-buffered saline with Tween 20 (TBST) at room temperature for 2 h before probing with anti-YKL-40 (1:700, Abcam) antibodies at 4 °C overnight, respectively. Then, the membranes were incubated with the secondary antibody for 1 h at room temperature. Finally, the blots were imaged on a hyper signal electrochemiluminescence (ECL) substrate. The gray values of the target bands were analysed by Image J software.
Transfections
HTR-8/SVneo cells were cultured in 6-well plate at a density of 5 × 105 cells/well for 24 h. Then, small interfering RNA for YKL-40 (si-YKL-40), negative control (si-NC), pcDNA3.1-Flag-YKL-40 (OE-YKL-40) and pcDNA3.1-Flag (OE-NC) were transfected into HTR-8/SVneo cells using a Lipofectamine 2000 Kit (Thermo Fisher Scientific, USA), respectively. After 24 h, fluorescence was detected under a fluorescence microscope, and the efficiency of transfection was determined by RT-qPCR. The cells were collected for protein extraction or follow-up with other biological function experiments at 48–96 h after transfection.
Cell proliferation
Briefly, transfected cell suspension (100 μL/well, 5 × 103 cells) was seeded into a 96-well plate in six replicates for each group at 37 °C and 5% CO2. The 10 μL of Cell Counting Kit-8 (Dojindo, Japan) reagent was added to the wells at 0 h, 24 h and 48 h respectively and sequentially incubated for 4 h. Then, the absorbance was measured at OD450 on an auto-microplate reader (Thermo, USA). Each sample was replicated at least three times.
Matrigel invasion and migration assay
This assay was performed as described previously (Wang et al. Citation2019b). For invasion assays, HTR-8/SVneo cells (1 × 105/well) in 100 μL serum-free RPMI1640 from the different treated groups were added in the upper part of a Transwell chamber (pre-covered with Matrigel). The medium with 20% FBS (700 µL) was added to the lower chamber. For migration assays, the same number of treated cells as the invasion assays was seeded in the upper chamber of each well, and 20% FBS (700 µL) was added to the lower chambers. After culturing at 37 °C for 26 h, the cells were fixed with methanol for 10 min before staining with haematoxylin for 8 min. Finally, the cells were counted from five fields at a × 200 magnification. Each experiment was repeated in triplicate.
Cell wound scratch assay
The scratch assay is used to determine the migratory ability of YKL-40 of HTR-8/SVneo cells. The cells transfected with siRNA or pcDNA plasmid were added into 6-well plate. After 24 h, a linear vertical scratch was drawn with a sterile 200 μL pipette tip. Images were captured under a microscope at 0 h and 24 h, and the wounds were selected at the same points. Wound areas (μm2) were measured using ImageJ, and the cell migration rate was calculated as follows: (area difference between 0 h and 24 h)/area of 0 h × 100%. The assay was repeated at least three times independently.
Cell apoptosis analysis
The cell apoptosis was measured by flow cytometry. The collected cells were stained with Annexin V-FITC/PI Apoptosis Detection Kit (MultiSciences, China) and incubated in the dark for 15 min. Finally, cell apoptosis was assessed on a FACS scan flow cytometer (Epics XL, USA) to calculate the percentages of necrotic and apoptotic cells in 10,000 events. The assay was performed in triplicates.
Statistical analysis
All statistical analyses were conducted using IBM SPSS 25.0 (IBM Corp, Armonk, NY, USA), and the graphs were plotted using GraphPad Prism 7.04 software (GraphPad, San Diego, CA, USA). For quantitative data, normal distribution was expressed by Student’s t-test with mean ± standard deviation (SD), while non-normal distribution was expressed by non-parametric test with percentile expression, and chi-square test was used for the count data. p < 0.05 was considered statistically significant.
Results
Clinical and demographic characteristics of the subjects
No statistically significant differences were detected between PAS and control groups according to demographic data, including age and Apgar scores, but the gestational age (p < 0.01) was significantly lower and caesarean hysterectomy was higher (p < 0.01) in the PAS group than the control group ().
Table 1. Demographic Characteristics in Two Groups.
Expression of YKL-40 in PAS and normal pregnant placental tissues
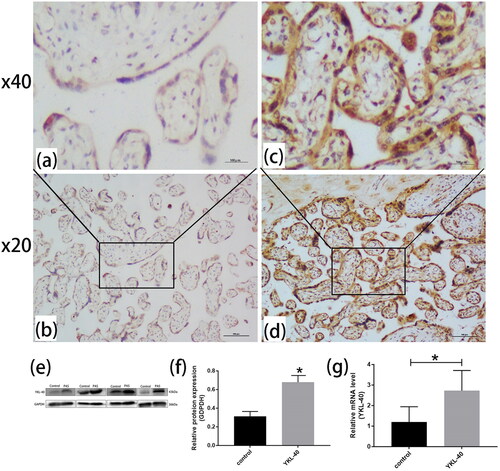
Immunohistochemistry showed that the YKL-40 level was significantly higher in the PAS group (0.484 ± 0.002) than in the control group (0.323 ± 0.003) (p < 0.01). This finding was consistent with the Western blot and RT-qPCR results ().
Figure 1. YKL-40 expression was higher in PAS than in control group (PAS: n = 25, control group: n = 22). Immunohistochemistry showed that YKL-40 was mainly expressed in the cell membrane and cytoplasm of placental villi. (a, b) Control group placental tissues were stained light brown, nevertheless, (c, d) PAS placental tissues were stained dark brown (magnification, ×200 and ×400, p < 0.01). In addition, (e, f) Western blotting showed YKL-40 was highly expressed in placental tissues in PAS patients than control groups (p < 0.05). (g) RT-qPCR was used to detect the expression of YKL-40 in placental tissues of PAS and control groups (P < 0.05).
The expression of YKL-40 mRNA and proteins by knockdown or overexpression of YKL-40 in human trophoblastic HTR-8/SVneo
Transfection efficiency was measured by RT-qPCR and Western blot ().
Figure 2. RT-qPCR and Western blot determined the expression of YKL-40 mRNA and proteins in HTR-8/SVneo after YKL-40 was knocked down or overexpressed, respectively (**P < 0.01). (a) RT-qPCR showed the mRNA levels of knocked out YKL-40 (**P < 0.01, si-NC vs. si-YKL-40). (c) Western blot indicated protein levels of knocked out YKL-40 (**P < 0.01, si-NC vs. si-YKL-40). (b) RT-qPCR displayed mRNA levels of overexpression of YKL-40 (**P < 0.01, OE-NC vs. OE-YKL-40). (d) Western blot demonstrated protein levels of overexpression YKL-40 (**P < 0.01, OE-NC vs. OE-YKL-40).
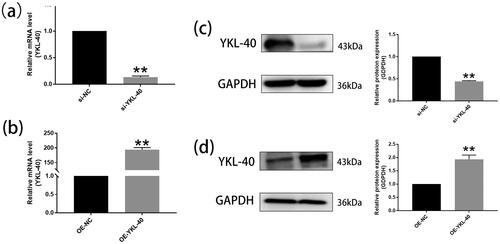
After YKL-40 was knocked down in HTR-8/SVneo for 24 h, the mRNA levels () were distinctly lower than controls with transfected blank (p < 0.01). The level of YKL-40 proteins () in the silencing group was significantly decreased compared to the si-NC group after cells were transfected for 48 h (p < 0.01). Similarly, the overexpressed YKL-40 mRNA and proteins () were significantly higher than in the OE-NC group (p < 0.01).
YKL-40 knockdown inhibited HTR-8/svneo cell proliferation, invasion, and migration and promoted cell apoptosis
The CCK-8 assay explored the proliferation of HTR-8/SVneo cells by knockdown of the YKL-40 gene, as shown in . We found that HTR-8/Svneo cells growth was inhibited significantly after transfection with si-YKL-40 for 24 and 48 h compared to the negative control (p < 0.05).
Figure 3. YKL-40 gene knockout inhibited the proliferation, invasion and migration of HTR-8/SVneo cells, and promoted apoptosis (*P < 0.05, **P < 0.01, magnification ×200; and f magnification ×200). (a) CCK-8 assays show that the proliferation of si-YKL-40 in cells is lower than in the negative control (24 h: P < 0.01; 48 h: P < 0.05). (b, f) Transwell and wound scratch assays were used to assess the migration of trophoblast cells. The migration viability of cells was significantly reduced after YKL-40 knockdown (**P < 0.01). (e) Transwell assays were performed to detect cell invasion. The invasion ability with silenced YKL-40 was weaker than in the si-NC group (**P < 0.01). (c, d) The apoptotic rates of HTR-8/SVneo cells were observed by flow cytometric assays (*P < 0.05).
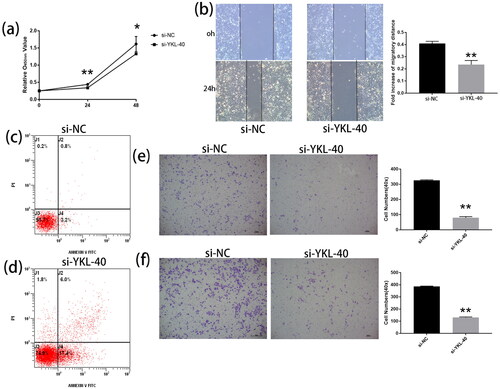
Wound scratch and Transwell assays showed that the migratory viability of silenced YKL-40 was significantly lower than that of the negative control (, p < 0.01). Subsequently, we used a Transwell assay and observed that the invasion ability of HTR-8/Svneo cells was weaker than that of the si-NC group after YKL-40 knockdown (, p < 0.01).
After the addition of the transfection reagent for 48 h, flow cytometry assessed the apoptosis rate of HTR-8/Svneo cells. The apoptosis rate of si-YKL-40 group was significantly higher compared to the negative control (p < 0.05). These results indicated that YKL-40 knockdown promoted apoptosis in HTR-8/Svneo cells (si-NC vs. si-YKL-40, 48 h, 4.0% vs. 23.4%), as shown in .
Overexpression of YKL-40 increased the proliferation, invasion and migration and decreased apoptosis of HTR-8/svneo cells
Transfection efficiency was measured by RT-qPCR 24 h after transfection (, p < 0.01).
Cell proliferation assay indicated that HTR-8/Svneo cells growth was significantly enhanced after transfection with OE-YKL-40 for 24 and 48 hours compared with OE-NC (Figure S1(a), p < 0.01).
Wound scratch and Transwell assays showed that the migration viability of HTR-8/SVneo was significantly increased after YKL-40 overexpression (Figure S1(b) and e, p < 0.01). Transwell invasion assay also showed that the invasive activity of HTR-8/Svneo cells was enhanced after YKL-40 overexpression compared with OE-NC group (Figure S1(f), p < 0.01).
Similarly, the apoptosis of HTR-8/Svneo cells in the OE-YKL-40 group was significantly lower than in the OE-control group (p < 0.05). The results indicated that overexpressed YKL-40 repressed the apoptosis of HTR-8/Svneo cells (Figure S1(c,d), OE-control vs OE-YKL-40, 48 h,7.8% vs. 2.9%).
(a) CCK-8 assay showed that the proliferation activity of OE-YKL-40 cells at 24h and 48h was significantly higher than that of over control cells (p < 0.05). (b, e) Wound scratch and Transwell assays were used to assess trophoblast cell migration. After YKL-40 overexpression, the cell migration activity was significantly increased (p < 0.01, magnification ×200). (f) In addition, Transwell assays were performed to examine cell invasion ability. The invasion ability with overexpressed YKL-40 was increased significantly than the OE-NC group (p < 0.01, magnification ×200). (c, d) The apoptotic rates of HTR-8/SVneo cells were measured by flow cytometric assays (p < 0.05).
YKL-40 regulated AKT/MMP9 signalling pathway in HTR8/SVneo cells
Previous studies have shown that YKL-40 regulated cell proliferation and motility, including tumour cell migration and invasion. Hence, we verified the related proteins, MMP9 and vital proteins involved in the PI3K/AKT downstream signalling pathway, including AKT phosphorylation (PAKT), AKT, after knockdown or overexpression of YKL-40 genes.
Western blotting showed that knockdown of YKL-40 (Figure S2(a)) significantly reduced the protein expression of PAKT and MMP9 (p < 0.01), while overexpression of YKL-40 (Figure S2(b)) activated the protein expression of PAKT and MMP9 (p < 0.01). In addition, AKT (Figure S2(a,b)) did not change significantly in each group (p > 0.05).
Discussion
In normal, uncomplicated pregnancies, extravillous trophoblasts invade through the uterine interstitium (interstitial trophoblasts) into spiral arteries, then line and remodel them (endovascular trophoblasts). Therefore, they are responsible for successfully establishing maternal blood flow into the placenta (Weiss et al. Citation2016, Xu et al. Citation2019a). Some evidences indicated that overexpressed YKL-40 in the serum/plasma has a high correlation with low survival in patients who have solid tumours (Bian et al. Citation2019). The function of YKL-40, a factor expressed in almost all tumour tissues (Böckelmann et al. Citation2021), promoted cell proliferation and invasion in tumours, but its role in the development of PAS has not been elucidated.
PAS is mainly caused by factors such as decreased, absent or abnormal decidua and/or defects in decidualization, which lead to increased penetration depth of (normal) extravillous trophoblast (Illsley et al. Citation2020). In addition, trophoblast and tumour metastasis are often similar in migration and invasion because they have similar molecular mechanisms (Kohan-Ghadr et al. Citation2016). Accumulating evidence demonstrated that YKL-40 knockdown or overexpression inhibited or promoted tumour cell migration and invasion (Jefri et al. Citation2015).
In the present study, 47 maternal cases were included, and the gestational age in the PAS group was lower and caesarean hysterectomy was higher than that in normal control group. The timing of delivery should be individualised. Hysterectomy has become the cornerstone of treatment in patients with PAS and postpartum haemorrhage. As for the termination time of PAS, when the mother and foetus are stable, the planned termination time varies according to different associations, and the American College of Obstetricians and Gynaecologists (ACOG) considered the best termination time to be 34–35+6 weeks (Allen et al. Citation2018). In addition, elective surgery under the condition of adequate preparation significantly reduced the risk of bleeding than emergency surgery, and also reduced the occurrence of many related complications (Miller et al. Citation2020). Therefore, the gestational age at termination of pregnancy in the PAS group was shorter than that in the control group, and no normal control pregnant women matched with the gestational age of PAS were obtained, which was also the limitation of this study. However, both groups were pregnant women in late pregnancy, which had limited impact on the study results. Among the 25 PAS cases, there was no significant difference in YKL-40 expression between 13 cases of placenta increta and 12 cases of placenta percreta (P > 0.05), which may be related to insufficient sample size and individual differences.
Hence, in this study, the expression and biological function of YKL-40 in the progression of PAS were investigated. Immunohistochemistry, RT-qPCR, and Western blot analysis indicated the up-regulation of both YKL-40 mRNA and protein in placental tissues of the pregnant woman with PAS. YKL-40 was consistent with the participation of Cripto-1 in controlling angiogenesis, a recent study concluded that serum Cripto-1 levels were significantly increased levels in pregnant women suffering from PAS than in pregnant women with PP and uncomplicated healthy pregnancies (Ozkose et al. Citation2022). Besides, further biological functions suggested that knockdown of YKL-40 inhibited cell proliferation, migration, invasion, and promoted cell apoptosis. Conversely, overexpression of YKL-40 promoted cell proliferation, invasion, migration, and hindered cell apoptosis. These findings are in line with previous studies suggesting a role for YKL-40 in promoting invasion of malignant tumour cells (Li et al. Citation2016, Chen et al. Citation2021) and that the up-regulated expression of YKL-40 may play a crucial role in PAS progression. The PI3K/Akt signalling pathway is fundamental in mediating the growth-factor-dependent regulation of trophoblast growth, migration, invasion, placental development, and foetal growth (Xu et al. Citation2019a, Xu et al. Citation2019b). A large number of studies suggested that the invasion and migration capability of HTR-8/SVneo cells and the expression of key kinases in PI3K/AKT/mTOR pathways were decreased when PI3K was inhibited (Zhu et al. Citation2016, Wang et al. Citation2019a). In addition, PI3K/AKT regulated trophoblast invasion and migration in human extravillous trophoblasts (EVTs) via the upregulation of MMP9 expression by epidermal growth factor (Liu et al. Citation2020). Another study indicated YKL-40 might affect the treatment efficacy of PI3K/AKT-based pathway inhibitors in glioblastoma (Wang et al. Citation2018).
The present results showed that the protein expression of YKL-40 was up-regulated in placental tissues from PAS. In addition, the Akt/MMP9 signalling pathway was activated by YKL-40 overexpression but inhibited by YKL-40 knockdown. Therefore, YKL-40 may play a role in the pathogenesis of PAS through Akt/MMP9 signalling pathway. Human trophoblastic cell invasion during placentation was regarded as limited to the invasion into uterine connective tissues and uterine spiral arteries (Moser et al. Citation2018). Although some studies have shown that trophoblastic cells are similar to tumour cell migration and invasion, YKL-40 can promote these phenomena, which are different from those in tumours. The invasion of trophoblastic cells during implantation and placentation is strictly restricted in both space and time (Kohan-Ghadr et al. Citation2016). Therefore, the mechanism by which YKL-40 promotes trophoblast invasion should be under intensive focus. The strength of this study is that we have demonstrated for the first time that YKL-40 may regulate PAS invasion through AKT/MMP9 signalling. Nevertheless, the present study has some limitations. We had not used inhibitors of the PI3K/AKT signalling pathway to study the effect of YKL-40 protein on trophoblast function through this pathway. And the biological function of YKL-40 was only verified using HTR-8/SVneo cells, which could be verified further with other pregnancy-related or primary cells.
Conclusion
In this study, we demonstrated that the expression and biological function of YKL-40 in placental tissues, and the expression was up-regulated in PAS. YKL-40 promoted HTR-8/SVneo cell proliferation, invasion, migration and inhibited apoptosis via Akt/MMP9 pathway.
Authors’ contributions
Weifang Liu and Runfang Wang conceived of the study. Suxin Liu and Xiaoqian Yin collected human tissue samples. Weifang Liu, Shengxian Li and Runfang Wang performed related experiments. Weifang Liu, Ruiling Zhang and Jia Li analysed the data and made figures. Weifang Liu wrote the first draft of the manuscript and Yan Huo performed revisions. All authors read and approved the final manuscript.
Supplemental Material
Download Zip (6.7 MB)Acknowledgements
The authors thank the obstetric medical staff and patients who participated in the study.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Data availability statement
The data underlying this article will be shared on reasonable request to the corresponding author.
Additional information
Funding
References
- Allen, L., et al., 2018. FIGO consensus guidelines on placenta accreta spectrum disorders: Nonconservative surgical management. International Journal of Gynaecology and Obstetrics: The Official Organ of the International Federation of Gynaecology and Obstetrics, 140 (3), 281–290.
- Bian, B., et al., 2019. Prognostic value of YKL-40 in solid tumors: a meta-analysis of 41 cohort studies. Cancer Cell International, 19, 259.
- Böckelmann, L.C., et al., 2021. YKL-40 protein expression in human tumor samples and human tumor cell line xenografts: implications for its use in tumor models. Cellular Oncology, 44 (5), 1183–1195.
- Chen, H.Y., et al., 2021. Knockdown of YKL-40 inhibits angiogenesis through regulation of VEGF/VEGFR2 and ERK1/2 signaling in endometrial cancer. Cell Biology International, 45 (12), 2557–2566.
- Ding, J., et al., 2021. M2 macrophage-derived G-CSF promotes trophoblasts EMT, invasion and migration via activating PI3K/Akt/Erk1/2 pathway to mediate normal pregnancy. Journal of Cellular and Molecular Medicine, 25 (4), 2136–2147.
- Gözükara, İ., et al., 2017. YKL-40 expression in abnormal invasive placenta cases. Journal of Perinatal Medicine, 45, 571–575.
- Graham, C.H., et al., 1993. Establishment and characterization of first trimester human trophoblast cells with extended lifespan. Experimental Cell Research, 206 (2), 204–211.
- Guvenir, H., et al., 2021. Can serum periostin, YKL-40, and osteopontin levels in pre-school children with recurrent wheezing predict later development of asthma? Pediatric Allergy and Immunology : official Publication of the European Society of Pediatric Allergy and Immunology, 32 (1), 77–85.
- Hao, H., et al., 2021. YKL-40 promotes invasion and metastasis of bladder cancer by regulating epithelial mesenchymal transition. Ann Med, 53, 1170–1178.
- Illsley, N.P., et al., 2020. Trophoblast invasion: lessons from abnormally invasive placenta (placenta accreta). Placenta, 102, 61–66.
- Jauniaux, E., Collins, S. and Burton, G.J., 2018. Placenta accreta spectrum: pathophysiology and evidence-based anatomy for prenatal ultrasound imaging. American Journal of Obstetrics and Gynecology, 218 (1), 75–87.
- Jefri, M., et al., 2015. YKL-40 regulated epithelial-mesenchymal transition and migration/invasion enhancement in non-small cell lung cancer. BMC Cancer, 15, 590.
- Jiang, H. and Li, H., 2021. Prognostic values of tumoral MMP2 and MMP9 overexpression in breast cancer: a systematic review and meta-analysis. BMC Cancer, 21 (1), 149.
- Johansen, J.S., et al., 1992. Identification of proteins secreted by human osteoblastic cells in culture. Journal of Bone and Mineral Research: The Official Journal of the American Society for Bone and Mineral Research, 7 (5), 501–512.
- Kohan-Ghadr, H.R., et al., 2016. Potential role of epigenetic mechanisms in regulation of trophoblast differentiation, migration, and invasion in the human placenta. Cell Adhesion & Migration, 10 (1-2), 126–135.
- Larionova, I., et al., 2020. Tumor-associated macrophages in human breast, colorectal, lung, ovarian and prostate cancers. Frontiers in Oncology, 10, 566511.
- Li, L.L., et al., 2016. Effects of a small interfering RNA targeting YKL-40 gene on the proliferation and invasion of endometrial cancer HEC-1A cells. International Journal of Gynecological Cancer : official Journal of the International Gynecological Cancer Society, 26 (7), 1190–1195.
- Liu, L., et al., 2021. CHI3L2 Is a novel prognostic biomarker and correlated with immune infiltrates in gliomas. Frontiers in Oncology, 11, 611038.
- Liu, Y., et al., 2020. Knockdown of activated Cdc42-associated kinase inhibits human extravillous trophoblast migration and invasion and decreases protein expression of pho-Akt and matrix metalloproteinase. The Journal of Maternal-Fetal & Neonatal Medicine, 33 (7), 1125–1133.
- Long, Y., et al., 2020. The expression and biological function of chemokine CXCL12 and receptor CXCR4/CXCR7 in placenta accreta spectrum disorders. Journal of Cellular and Molecular Medicine, 24 (5), 3167–3182.
- Madazli, R., et al., 2008. Chitotriosidase and YKL-40 in normal and pre-eclamptic pregnancies. International Journal of Gynaecology and Obstetrics: the Official Organ of the International Federation of Gynaecology and Obstetrics, 100 (3), 239–243.
- Miller, R., Timor-Tritsch, I.E. and Gyamfi-Bannerman, C, 2020. Society for maternal-fetal medicine (SMFM) consult series #49: Cesarean scar pregnancy. American Journal of Obstetrics and Gynecology, 222 (5), B2–b14.
- Mogos, M.F., et al., 2016. Recent trends in placenta accreta in the United States and its impact on maternal-fetal morbidity and healthcare-associated costs, 1998–2011. The Journal of Maternal-Fetal & Neonatal Medicine, 29 (7), 1077–1082.
- Moser, G., et al., 2018. Human trophoblast invasion: new and unexpected routes and functions. Histochemistry and Cell Biology, 150 (4), 361–370.
- Oğlak, S.C., Ölmez, F. and Tunç, Ş., 2022. Evaluation of antepartum factors for predicting the risk of emergency cesarean delivery in pregnancies complicated with placenta previa. The Ochsner Journal, 22 (2), 146–153.
- Oh, I.H., Pyo, J.S. and Son, B.K., 2021. Prognostic impact of YKL-40 immunohistochemical expression in patients with colorectal cancer. Current Oncology (Toronto, Ont.), 28 (4), 3139–3149.
- Ozkose, Z.G., et al., 2022. Maternal serum cripto-1 levels in pregnancies complicated with placenta previa and placenta accreta spectrum (PAS). J Coll Physicians Surg Pak, 32, 1570–1575.
- Pan, X.Y., et al., 2015. A marked increase in obstetric hysterectomy for placenta accreta. Chinese Medical Journal, 128 (16), 2189–2193.
- Puthumana, J., et al., 2021. Biomarkers of inflammation and repair in kidney disease progression. J Clin Invest, 131 (3), e139927.
- Raeeszadeh-Sarmazdeh, M., Do, L.D. and Hritz, B.G., 2020. Metalloproteinases and their inhibitors: potential for the development of new therapeutics. Cells, 9 (5), 1313.
- Wang, L., et al., 2019a. Reduced ELABELA expression attenuates trophoblast invasion through the PI3K/AKT/mTOR pathway in early onset preeclampsia. Placenta, 87, 38–45.
- Wang, N., et al., 2020. Combination of tanshinone IIA and astragaloside IV attenuate atherosclerotic plaque vulnerability in ApoE(-/-) mice by activating PI3K/AKT signaling and suppressing TRL4/NF-κB signaling. Biomedicine & Pharmacotherapy, 123, 109729.
- Wang, Q., et al., 2019b. Down-regulated long non-coding RNA PVT1 contributes to gestational diabetes mellitus and preeclampsia via regulation of human trophoblast cells. Biomedicine & Pharmacotherapy, 120, 109501.
- Wang, Y., et al., 2018. Differential regulation of the pro-inflammatory biomarker, YKL-40/CHI3L1, by PTEN/Phosphoinositide 3-kinase and JAK2/STAT3 pathways in glioblastoma. Cancer Letters, 429, 54–65.
- Weiniger, C.F., 2020. What’s new in obstetric anesthesia in 2018? International Journal of Obstetric Anesthesia, 42, 99–108.
- Weiss, G., et al., 2016. Arterial endothelial cytokines guide extravillous trophoblast invasion towards spiral arteries; an in-vitro study with the trophoblast cell line ACH-3P and female non-uterine endothelial cells. Placenta, 38, 49–56.
- Xu, B., et al., 2019a. FCPR03, a novel phosphodiesterase 4 inhibitor, alleviates cerebral ischemia/reperfusion injury through activation of the AKT/GSK3β/β-catenin signaling pathway. Biochemical Pharmacology, 163, 234–249.
- Xu, Y., et al., 2019b. ANXA4 promotes trophoblast invasion via the PI3K/Akt/eNOS pathway in preeclampsia. American Journal of Physiology. Cell Physiology, 316 (4), C481–c491.
- Yeo, I.J., et al., 2019. Roles of chitinase 3-like 1 in the development of cancer, neurodegenerative diseases, and inflammatory diseases. Pharmacology & Therapeutics, 203, 107394.
- Zhang, B., et al., 2020. Identification of prognostic biomarkers for multiple solid tumors using a human villi development model. Frontiers in Cell and Developmental Biology, 8, 492.
- Zhu, X., et al., 2016. Knockdown of TACC3 inhibits trophoblast cell migration and invasion through the PI3K/Akt signaling pathway. Molecular Medicine Reports, 14 (4), 3437–3442.