

Abstract
Background
Despite the strong evidence concerning carcinogenic roles of glucagon-like peptide 1 receptor (GLP1R), the role of this gene in endometrial cancer (EC) remains elusive. This study investigated the properties of GLP1R on EC in vitro.
Methods
The expression of GLP1R in EC was detected by RT-qPCR, immunohistochemistry, and western blotting. Cell viability, cell cycle, apoptosis, migration, invasion and ferroptosis were assessed through CCK-8, flow cytometry, wound healing, transwell, DCFH-DA and western blotting, respectively.
Results
We found that GLP1R was up-regulated in EC than normal specimens. It had the highest expression in AN3CA cells. Cell viability, migration and invasion were significantly reduced, while cell cycle arrest and apoptosis were induced following GLP1R knockdown. The malignant biological behaviours of AN3CA cells were investigated when treated with exendin-4 (GLP1R agonist). Moreover, GLP1R lowered intracellular ROS level and expression of SLC7A11, and FTH1, but mitigated GPX4 expression in AN3CA cells.
Conclusion
In a word, GLP1R was up-regulated in EC and its up-regulation facilitated the proliferative and metastatic potentials, and protected cells from ferroptosis, thereby accelerating EC progression. These data emphasised the potency of GLP1R as a therapeutic agent against EC.
PLAIN LANGUAGE SUMMARY
Endometrial cancer (EC) is the second most common form of gynaecologic malignancy, with over 189,000 new cases and about 45,000 deaths worldwide per annum. The effects of glucagon-like peptide 1 receptor (GLP1R) in cancers such as colon and pancreatic cancers have been uncovered. However, whether GLP1R affects EC progression especially ferroptosis process remains elusive. In this study, up-regulation of GLP1R promotes the proliferative and metastatic potentials of EC cells, and protects EC cells from ferroptosis. The opposite results are observed in GLP1R knocking-down. Our study found that GLP1R may exert an oncogene function in EC cells, which can affect proliferative, migrated as well as invasive capacities of EC cells. Moreover, it protected EC cells from ferroptosis. Thus, our results expanded the understanding of the function of GLP1R protein and offered insights into the targeted treatment strategies against EC.
Keywords:
Introduction
Endometrial cancer (EC) ranks the second most commonly diagnosed gynaecologic malignancy following ovarian carcinoma (Lees et al. Citation2021). The global incidence rate of EC has been steadily increasing in recent years, accompanied by a notable trend of earlier onset, particularly among women in their 30s (Matsuo et al. Citation2021). EC progression is a complex process with multiple factors and steps. Long-term oestrogen stimulation of the endometrium and/or absence of progesterone antagonism is widely acknowledged as the primary determinant leading to endometrial hyperplasia and cancer (Zhou et al. Citation2019). Furthermore, there are various risk factors that induce EC, like hypertension, adiposity, and diabetes (Carugno et al. Citation2021). The current clinical management of EC primarily relies on surgical intervention in conjunction with adjuvant therapies such as chemotherapy and endocrine therapy (Forsse et al. Citation2020, Matsuo et al. Citation2021). However, the survival rates are still very dismal. Furthermore, advanced or recurrent patients present a more unfavourable clinical outcome due to chemoresistance (Xiao et al. Citation2021). The identification and validation of novel, efficient biomarkers for molecular diagnostics, targeted therapies, and prognostic monitoring in EC pose a challenging task that necessitates thorough exploration and resolution within the realms of current basic research, clinical diagnosis, precision treatment, and prognosis evaluation (Arend et al. Citation2018, Murali et al. Citation2018, Sobecki-Rausch and Barroilhet Citation2019). Hence, it is mperative to investigate and elucidate the pathogenesis of EC, as well as clarify the biological behaviours of EC cells in malignant transformation processes.
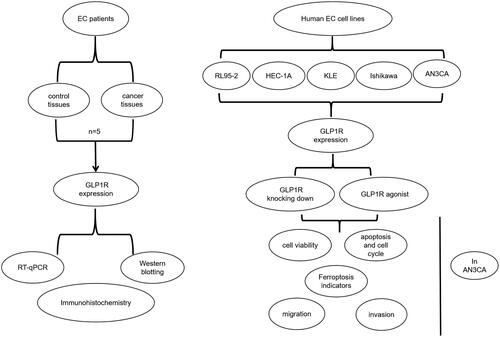
Ferroptotic cell death is a newly discovered type to modulate cell death (Liang et al. Citation2019). Accumulated evidence demonstrates the potential of inducing ferroptosis for cancer treatment, especially for eradication of cancers that display the resistance to traditional treatment (Xing et al. Citation2022). A seven-transmembrane protein encoded by GLP1R gene, a receptor of GLP1, can stimulate insulin secretion induced by glucose and pancreatic β-cell proliferation (Jones et al. Citation2018). Polymorphisms of GLP1R gene exhibit associations with diabetes (Suzuki et al. Citation2019). GLP1R-based therapy provides a basis for treating diabetes. In reproduction, it is involved in energy metabolism (Timper et al. Citation2020), G protein-coupled receptor pathway (Ast et al. Citation2020) and the like. Recently, carcinogenic roles of GLP1R have garnered significant attention. As an example, Toru Shigeoka et al. GLP1R up-regulation may attenuate prostate carcinoma growth via restraining cell cycle development (Shigeoka et al. Citation2020). However, whether GLP1R affects EC progression via ferroptosis signalling remains elusive. In this study, GLP1R up-regulation was detected in EC specimens. Silencing of GLP1R significantly inhibited the proliferative, migrated as well as invasive capacities of AN3CA cells. Also, its knockdown led to cell cycle disorders and induced cell apoptosis and ferroptosis. These data were suggestive of the functions of GLP1R on endometrial carcinoma at the cellular level. The flow chart of this study was shown in .
Figure 1. A flow chart of this study. EC: endometrial cancer; GLP1R: glucagon-like peptide 1 receptor; RT-qPCR: quantitative reverse transcription PCR. RL95-2, HEC-1A, KLE, Ishikawa and AN3CA are human EC cell lines.
Methods
Quantitative reverse transcription PCR (RT-qPCR)
Tissue or cell samples were homogenised and broken, and then added to 1 mL TRIzol™ reagent (15596018; Invitrogen, Carlsbad, CA). A total of 1 μL RNA was used to calculate the purity, followed by reversible transcription into cDNA productions. Relevant procedures were listed as follows: 37 °C for 10 min; then 85 °C for 5 s; and final 4 °C for hold. Primer used in this research were as follows: GLP1R, 5′-TCGCTGTGAAAATGAGGAGG-3′ (forward), 5′-TTGGCTGAGGTTAGAAGAGCC-3′(reverse); GAPDH, 5′-TCAAGAAGGTGGTGAAGCAGG-3′(forward), 5’TCAAAGGTGGAGGAGTGGGT-3′ (reverse). GAPDH was used for normalisation and GLP1R amounts were reckoned with 2−ΔΔCt approach.
Western blotting
After being lysed, the supernatant was collected for protein extraction. The protein concentration was determined using the BCA method (P0009; Beyotime, Beijing, China). After separation by 10% SDS-PAGE electrophoresis, the samples were subsequently transferred onto the PVDF membrane and then sealed with 5% milk. The membrane had the incubation with primary antibodies against GLP1R (1:600), GAPDH (1:5000;), SLC7A11 (1:1000), FTH1 (1:1000), and GPX4 (1:1000; P36969) at 4 °C overnight. Then, the PVDF membrane had the incubation with the horseradish peroxidase-labeled secondary antibody (1:5000). All the antibodies were got from RayBiotech (Shanghai, China). Protein bands were observed with a gel imaging system.
Immunohistochemistry
The sections were embedded in paraffin, deparaffinized and subsequently incubated in a 3% H2O2 solution under dark conditions. Afterwards, they were blocked with 3% BSA (A8020; Solarbio, Beijing, China) at room temperature for 30 min. During this step, the primary antibody was added and incubated overnight at 4 °C. The slices were washed three times, with each time for 5 min, followed by mixture with secondary antibody (Proteintech) at 20 °C for 1 h. After DAB (K5007; DAKO, Shanghai, China) development, the slices were redyed 3 min by Harris haematoxylin. Subsequently, dehydration was performed followed by fixation with neutral gum. Finally, image acquisition and analysis were conducted.
Cell culture
Human EC cells including RL95-2, HEC-1A, KLE, Ishikawa and AN3CA were cultured in DMEM medium (Hyclone, Beijing, China) that contained 10% FBS (SH30084.03; Hyclone, Beijing, China) and 1% streptomycin/penicillin, with the conditions of 5% CO2 at 37 °C.
Transfection and treatment
Following aspirating the original culture medium, AN3CA cells were added to PBS and digested by trypsin for 2 min. Then, cell suspension was plated into 6-well plates (3 × 105 cells/well) and cultured overnight. Three siRNAs targeting GLP1R (20 μM) were incubated by 100 μL opti-MEM for 5 min. Then, siRNA mix was incubated by Lipofectamine mix for 5 min. Furthermore, AN3CA cells were treated with 100 nmol/L exendin-4 (GLP1R agonist). To inhibit ferroptosis, AN3CA cells were exposed to 60 nmol/L ferrostatin-1.
Cell counting kit-8 (CCK-8) tests
A CCK-8 kit from Dojindo (Kumamoto, Japan) was used for the measurement of cell viability. A 100 μL cell suspension () was dispensed into each well of a 96-well culture plate, with a seeding density of 4500 cells per well, for subsequent culturing. Cell survival was assessed at the respective time points. Then, cell viability was assessed via a microplate detector after CCK-8 solution addition into the well.
Flow cytometry analysis for cell cycle and apoptosis
Cell samples were centrifugated, and the cell suspension was collected and added to pre-cooled ethanol at −20 °C for overnight fixation at 4 °C. After ethanol removal, the cells were incubated with 300 μL of DNA staining solution in the dark. For apoptosis assessment, cells were resuspended in 100 μL of Annevix binding buffer after centrifugation, followed by the addition of 5 μL Annevix FITC and 5 μL PI in the dark. With the aid of flow cytometry system from Beckman Coulter (USA), cell cycle and apoptosis were assessed using a flow cytometry system provided by Beckman Coulter (USA).
Wound healing experiments
Cell suspension was seeded onto the six well plate, and once the cells reached 100% confluence a 10 μL pipette tip was utilised to create a scratch. Following aspirating the cell culture fluid, the plate was rinsed with PBS three times. Subsequently, DMEM was added once the generated cell debris had been washed away by PBS. The cells were cultivated continuously and images were captured at corresponding time points.
Transwell assay
The invasive capacity was monitored assessed using the transwell assay. A total of 50 μL Matrigel (356234; BD Biocoat, USA) diluted five-fold with serum-free medium was used for pre-coating the upper chamber. The transfected cells were digested for 24 h. After terminating the digestion, the culture medium was removed by centrifugation. After being washed with PBS, the resuspended cells (1 × 105) were cultured in the transwell chamber and incubated for 24 h. Afterwards, cells were fixed, washed and then stained with crystal violet for 15 min. The invaded cells were observed using a microscope, and three fields of view were randomly selected for image acquisition.
Intracellular reactive oxygen species (ROS) productions
We quantified levels of ROS using a ROS detection kit from Beyotime (Shanghai, China). Cell samples were incubated with DCFH-DA (10 μmol/L) for 20 min, followed by rinsing with serum-free DMEM. Subsequently, images were captured using a laser confocal microscope.
Statistical analysis
Data analysis was done in GraphPad Prism 8.0 and the values were presented in the form of mean ± standard deviation. Each experiment was repeated three times. Student’s t test or one-way ANOVA was adopted to gauge the differences across groups. p < 0.05 means statistical difference.
Patients and specimens
From 2019 to 2020, five pairs of fresh EC and normal tissues were obtained in our hospital. This research was abided by the Declaration of Helsinki, conformed to the Enhancing the Quality and Transparency Of health Research (EQUATOR) network guidelines and supported by our hospital’s Ethics Committee (approval number: 2019038). All participants provided written informed consent.
Results
Up-regulation of GLP1R is observed in EC tissues and cell lines
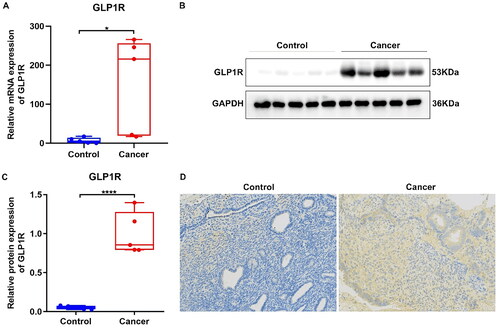
We evaluated GLP1R expression in five paired EC and normal tissue specimens. Compared to normal tissues, there was a significant increase in GLP1R mRNA levels observed in EC tissues (, p < 0.05). Meanwhile, an up-regulation of GLP1R was also observed at the protein level in tumour tissues relative to normal tissues (, p < 0.0001). In , we performed immunohistochemistry and confirmed that the expression of GLP1R protein was predominantly localised to the plasma membrane of tumour cells. Additionally, GLP1R amounts were also scrutinised in five EC cell lines (RL95-2, HEC-1A, KLE, Ishikawa and AN3CA). In Supplementary Figure 1(A), the expression of GLP1R was detected in these cell lines, with AN3CA cells exhibiting the highest mRNA expression level (p < 0.0001). Furthermore, western blotting confirmed its highest protein expression in AN3CA cells (Supplementary Figure 1(B,C), p < 0.0001). Hence, AN3CA cells were chosen for the succeeding assays. Three siRNAs against GLP1R were transfected into AN3CA cells. RT-qPCR verified that GLP1R exhibited relatively low expression in AN3CA cells transfected with siRNA-1 (Supplementary Figure 1(D), p < 0.0001). To activate GLP1R expression, AN3CA cells were treated by 100 nmol/L exendin-4. The results of RT-qPCR analysis demonstrated a distinct upregulation of GLP1R expression in response to exendin-4 treatment compared to the control group (Supplementary Figure 1(E), p < 0.01).
Figure 2. Up-regulation of GLP1R in EC tissue specimens. (A) RT-qPCR for GLP1R mRNA expression between tumour and normal samples. (B,C) Western blotting for GLP1R protein between tumour and normal samples. (D) Immunohistochemistry of GLP1R protein in tumour and normal samples. Positive cells are stained by DAB and appear as brown, and cell nucleus is stained by haematoxylin as blue. *p < 0.05 and ****p < 0.0001.
GLP1R up-regulation facilitates EC cell viability
After treatment with siGLP1R or a GLP-1 agonist, we assessed the viability of AN3CA cells. At 0 h, no discernible changes were observed following transfection with siGLP1R or GLP-1 agonist (Supplementary Figure 2(A)). At 24 h, GLP-1 agonist treatment significantly increased survival rate by contrast to controls (Supplementary Figure 2(B), p < 0.05). At 36 h, compared to si-negative control (NC) group, survival rate was markedly lowered in siGLP1R group (Supplementary Figure 2(C), p < 0.01). At 48 and 72 h, lower survival rate was found in siGLP1R group compared with siNC group (Supplementary Figure 2(D,E), p < 0.001). Meanwhile, compared to control, GLP-1 significantly enhanced survival rate of AN3CA cells (p < 0.001). The above data suggests that knockdown of GLP1R results in a significant decrease in cell viability, while up-regulation of GLP1R promotes EC cell viability, in a time dependent manner.
GLP1R knockdown induces cell cycle arrest EC cells
As manifested in Supplementary Figure 3(A,B), number of cells in the G1 and G0 phases was significantly increased in the siGLP1R group compared to the control group (p < 0.01), while there was a significant decrease in cells in the S phage (p < 0.05). Following activation of the GLP1R, there was a distinct reduction in cells in the G1 and G0 phases (p < 0.001), while the percentage of cells in the S phage showed a significant up-regulation (p < 0.01). Thus, knockdown of GLP1R induced G0/G1 arrest, while its activation suppressed such arrest in EC cells.
GLP1R up-regulation exhibits the inhibitory effect on apoptosis of EC cells
The apoptosis of AN3CA cells was investigated using flow cytometry (Supplementary Figure 4(A,B)). The apoptotic rate of AN3CA cells in siGLP1R group was significantly higher than the siNC group (p < 0.0001). In comparison to control group, lower apoptotic rate was observed in AN3CA cells treated with GLP-1 agonist (p < 0.0001). Hence, knockdown of GLP1R induced apoptosis and its up-regulation suppressed apoptosis in AN3CA cells.
GLP1R up-regulation heightens the migrated and invasive capacities of EC cells
The migratory capacity of AN3CA cells was assessed using the wound healing assay. Following a 24 h treatment with GLP-1 agonist, there was a significant increase in the migrated distance compared to the control group (Supplementary Figure 5(A,B), p < 0.01). Interestingly, 48 h after transfection with siGLP1R, the migrated distance was reduced compared with siNC (p < 0.01). Meanwhile, GLP-1 agonist markedly extended the migrated distance by contrast to controls (p < 0.05). Additionally, the invasive ability of AN3CA cells was evaluated through transwell assay. As illustrated in Supplementary Figure 6(A,B), compared to siNC group, the number of invasive cells was reduced in siGLP1R group (p < 0.05). Additionally, the GLP-1 agonist significantly increased the number of invasive cells (p < 0.001). These findings suggested that up-regulation of GLP1R plays a stimulatory role in promoting migration and invasion capabilities of EC cells.
GLP1R protects EC cells from ferroptosis
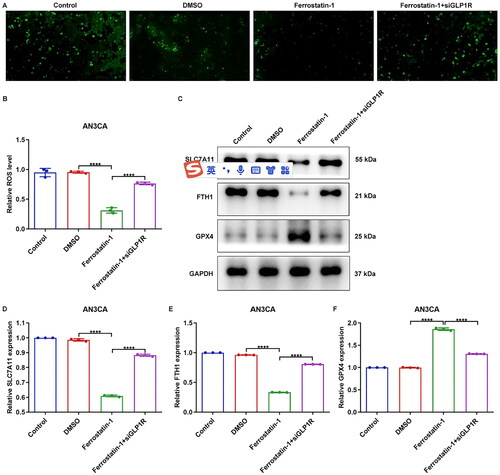
The levels of ferroptosis indicators, including intracellular ROS, SLC7A11, FTH1 and GPX4, were quantified in AN3CA cells. As manifested in , in comparison to siNC group, ROS amounts were elevated in siGLP1R group (p < 0.0001). However, lower ROS expression was captured in GLP-1 agonist-treated cells (p < 0.0001). Furthermore, our findings revealed that the siGLP1R group exhibited significantly upregulated expression of SLC7A11 and FTH1, while displaying downregulated expression of GPX4 compared to the siNC group (, p < 0.0001). Interestingly, GLP-1 agonist notably attenuated expression of SLC7A11 and FTH1, but elevated GPX4 expression in AN3CA cells (p < 0.0001). In addition, we further indicated that the ferroptosis inhibitor, ferrostatin-1, effectively attenuated the intracellular ROS level in AN3CA cells (, p < 0.0001). Interestingly, siGLP1R ameliorated the inhibitory effects of ferrostatin-1 on intracellular ROS level (p < 0.0001). At the same time, the expression levels of SLC7A11, and FTH1 were downregulated, while GPX4 expression was elevated in AN3CA cells treated with ferrostatin-1 (, p < 0.0001). However, these effects were reversed upon siGLP1R treatment (p < 0.0001). Altogether, GLP1R upregulation may protect EC cells from ferroptotic cell death.
Figure 3. GLP1R protects EC cells from ferroptosis. (A,B) Intracellular ROS level ofAN3CA cells treated with siGLP1R or GLP-1 agonist. (C–F) Expression of SLC7A11, FTH1 and GPX4 in AN3CA cells treated with siGLP1R or GLP-1 agonist. ****p < 0.001.
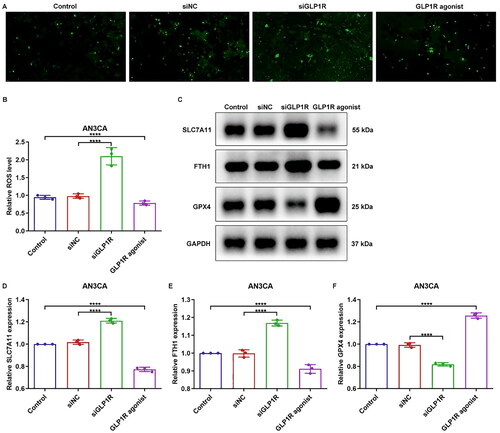
Figure 4. Suppression of GLP1R attenuates the inhibitory effects of ferrostatin-1 on ferroptosis of EC cells. (A,B) Intracellular ROS level of AN3CA cells treated with ferrostatin-1 and/or siGLP1R. (C–F) Expression of SLC7A11, FTH1 and GPX4 in AN3CA cells treated with ferrostatin-1 and/or siGLP1R. ****p < 0.001.
Discussion
Currently, the pathogenesis of EC remains poorly understood. Therefore, it is crucial to explore novel therapeutic strategies for EC (Vetter et al. Citation2019). Our data unequivocally validated the up-regulation of GLP1R in EC specimens. The up-regulation exerted the carcinogenic roles in EC. Knockdown of GLP1R effectively attenuated the proliferative, migratory and invasive capacities of AN3CA cells. These findings to some extent showed the potential potency of GLP1R as a promising molecular agent for EC treatment.
We observed a significant up-regulation of GLP1R in EC tissue specimens compared to normal samples, indicating its potential carcinogenic role in EC. GLP1R overexpression has been discovered in prostate carcinoma (Zhao et al. Citation2020), breast carcinoma (Chikayo et al. Citation2017) and other malignancies. Consistently, GLP1R was primarily distributed in cytomembrane of EC cells (Kanda et al. Citation2018). The function of GLP1R in AN3CA cells was further investigated. The immortal proliferation is a characteristic of EC cells. The up-regulation of pro-proliferation factors and the down-regulation of anti-proliferation factors can both lead to uncontrolled cell proliferation, resulting in unlimited growth potential. Silencing GLP1R inhibited cellular proliferative ability of AN3CA cells. Conversely, the GLP-1 agonist exendin-4 elicited a stimulatory effect on AN3CA cell proliferation. Previously, GLP1R activation lessened proliferative capacities of prostate carcinoma cells (Li et al. Citation2017) and ovarian cancer cells (He et al. Citation2016). Cell cycle changes are a key factor affecting tumour progression. Cell cycle arrest leads to a prolonged duration of the cell cycle, thereby impacting the proliferative capacities of neoplastic cells. We found that after disrupting GLP1R, the cell cycle was mainly blocked in G0/G1 phase. Previously, exendin-4 heightened apoptosis of ovarian carcinoma (He et al. Citation2016), colon carcinoma (Koehler et al. Citation2011) and pancreatic carcinoma cells (Zhao et al. Citation2014). "In this study, knockdown of GLP1R significantly enhanced apoptotic levels in AN3CA cells, whereas exendin-4 attenuated apoptotic levels in AN3CA cells.
The invasion of EC heightens the risk of tumour progression and patient mortality (Bogani et al. Citation2019, Moro et al. Citation2019). Some patients have metastases when diagnosed, and clinically stage III or IV EC patients account for about 30% of all cases (Bakkum-Gamez et al. Citation2014, Kito et al. Citation2015). The metastasis of EC involves complex series of stages, beginning with weakened cell adhesion leading to detachment from the primary site and acquisition of migratory and invasive capabilities, followed by local infiltration into blood vessels and lymphatic vessels, ultimately resulting in distant metastasis (Zhu et al. Citation2019). Finally, shed tumour cells establish secondary growth in new environments. Therefore, it is imperative to investigate the mechanism underlying invasion and metastasis in EC to develop effective anti-tumour strategies that can prolong patient survival (Mahecha et al. 2017, Mali et al. Citation2019; Casey and Singh Citation2020). Herein, migrated and invasive abilities were suppressed by silencing GLP1R, which were activated by exendin-4. Nie et al. reported that exendin-4 inhibited glioma cell migrated as well as invasive phenotypes by GLP1R/sirt3 pathway (Nie et al. Citation2018). Moreover, Luciani et al. confirmed that exendin-4 induced invasive potential of neuroblastoma cells (Paola et al. Citation2013). Ana Ines Cases et al. emphasised the auxo-action of GLP1R on migration as well as invasion of pancreatic ductal adenocarcinoma cells (Cases et al. Citation2015).
Ferroptosis is triggered by oxidative perturbation of the intracellular microenvironment, which is constitutively regulated by GPX4 (Chen et al. Citation2021). The iron-dependent accumulation of ROS and lipid peroxidation remain pivotal in the induction of ferroptotic cell death, which can be attenuated by iron chelators and lipophilic antioxidants (Su et al. Citation2020). Previous data supports the crucial role of ferroptosis in attenuating the growth and survival of EC cells (Wang et al. Citation2021). The current study demonstrated that GLP1R effectively reduced intracellular ROS level and downregulated the expression of SLC7A11 and FTH1, while simultaneously upregulating GPX4 expression in AN3CA cells, thereby conferring protection against ferroptotic cell death. Our findings suggested that GLP1R, as an oncogene, plays a crucial role in sustaining the continuous proliferation and self-renewal of the malignant phenotype in EC cells. Consequently, targeting GLP1R may hold promise as a potential therapeutic strategy for treating EC.
Conclusion
Collectively, our study reveals that GLP1R may possess an oncogene role in EC cells, influencing their proliferative, migratory and invasive capacities. Moreover, it confers protection against ferroptosis in EC cells. Consequently, our findings enhance the comprehension of GLP1R protein function and provide valuable insights for targeted therapeutic strategies against EC.
Authors’ contributions
Xinmei Zhang made substantial contributions to the conception and design of the work. Wu Li made substantial contributions to the acquisition, analysis and interpretation of data for the work, and drafted the manuscript. Wen Lyu, Songjun Liu and Fan Ruan revised the manuscript critically for important intellectual content. All authors have read the manuscript, agreed to be accountable for all aspects of the work in ensuring that questions related to the accuracy and agreed final approval of the version to be published.
Ethial approval
This research was abided by the Declaration of Helsinki and supported by Tongde Hospital of Zhejiang Province’s Ethics Committee (approval number: 2019038). All participants provided written informed consent.
Supplemental Material
Download Zip (2.1 MB)Disclosure statement
No potential conflict of interest was reported by the author(s).
Data availability statement
The datasets analysed during the current study are available from the corresponding author on reasonable request.
Additional information
Funding
References
- Arend, R. C., et al., 2018. Endometrial cancer: Molecular markers and management of advanced stage disease. Gynecologic Oncology, 150 (3), 569–580.
- Ast, J., et al., 2020. Super-resolution microscopy compatible fluorescent probes reveal endogenous glucagon-like peptide-1 receptor distribution and dynamics. Nature Communications, 11 (1), 5160.
- Bakkum-Gamez, J. N., et al., 2014. Efficacy of contemporary chemotherapy in stage IIIC endometrial cancer: A histologic dichotomy. Gynecologic Oncology, 132 (3), 578–584.
- Bogani, G., et al., 2019. Low-volume disease in endometrial cancer: the role of micrometastasis and isolated tumor cells. Gynecologic Oncology, 153 (3), 670–675.
- Carugno, J., et al., 2021. New development on hysteroscopy for endometrial cancer diagnosis: State of the art. Minerva Medica, 112 (1), 12–19.
- Cases, A. I., et al., 2015. Significance of expression of glucagon-like peptide 1 receptor in pancreatic cancer. Oncology Reports, 34 (4), 1717–1725.
- Casey, L. and Singh, N., 2020. Metastases to the ovary arising from endometrial, cervical and fallopian tube cancer: recent advances. Histopathology, 76 (1), 37–51.
- Chen, L., et al., 2021. Characterization of interplay between autophagy and ferroptosis and their synergistical roles on manipulating immunological tumor microenvironment in squamous cell carcinomas. Frontiers in Immunology, 12, 739039.
- Chikayo, I., et al., 2017. Exendin-4, a glucagon-like peptide-1 receptor agonist, attenuates breast cancer growth by inhibiting NF-κB activation. Endocrinology, (12), 4218.
- Forsse, D., et al., 2020. Maintained survival outcome after reducing lymphadenectomy rates and optimizing adjuvant treatment in endometrial cancer. Gynecologic Oncology, 160 (2), 396–404.
- He, W., et al., 2016. Exendin-4 inhibits growth and augments apoptosis of ovarian cancer cells. Molecular and Cellular Endocrinology, 436, 240–249.
- Jones, B., et al., 2018. Targeting GLP-1 receptor trafficking to improve agonist efficacy. Nature Communications, 9 (1), 1602.
- Kanda, R., et al., 2018. Expression of the glucagon-like peptide-1 receptor and its role in regulating autophagy in endometrial cancer. BMC Cancer, 18 (1), 657.
- Koehler, J. A., Kain, T. and Drucker, D. J., 2011. Glucagon-like peptide-1 receptor activation inhibits growth and augments apoptosis in murine CT26 colon cancer cells. Endocrinology, 152 (9), 3362–3372.
- Lees, B., et al., 2021. A population-based study of causes of death after endometrial cancer according to major risk factors. Gynecologic Oncology, 160 (3), 655–659.
- Li, X. N., et al., 2017. Glucagon-like peptide-1 analogues inhibit proliferation and increase apoptosis of human prostate cancer cells in vitro. Experimental and Clinical Endocrinology & Diabetes: Official Journal, German Society of Endocrinology [and] German Diabetes Association, 125 (2), 91–97.
- Liang, C., et al., 2019. Recent progress in ferroptosis inducers for cancer therapy. Advanced Materials, 31 (51), e1904197.
- Mahecha, A. M., and Wang, H., 2017. The influence of vascular endothelial growth factor-A and matrix metalloproteinase-2 and -9 in angiogenesis, metastasis, and prognosis of endometrial cancer. OncoTargets & Therapy, 10, 4617–4624.
- Mali, A. V., et al., 2019. Anticancer and antimetastatic potential of enterolactone: clinical, preclinical and mechanistic perspectives. European Journal of Pharmacology, 852, 107–124.
- Matsuo, K., et al., 2021. Ovarian conservation for young women with early-stage, low-grade endometrial cancer: a 2-step schema. American Journal of Obstetrics and Gynecology, 224 (6), 574–584.
- Moro, F., et al., 2019. Synchronous primary cancers of endometrium and ovary vs endometrial cancer with ovarian metastasis: an observational study. Ultrasound in Obstetrics & Gynecology, 53 (6), 827–835.
- Murali, R., et al., 2018. Evolving roles of histologic evaluation and molecular/genomic profiling in the management of endometrial cancer. Journal of the National Comprehensive Cancer Network: Jnccn, 16 (2), 201–209.
- Nie, Z. J., et al., 2018. Exendin-4 inhibits glioma cell migration, invasion and epithelial-to-mesenchymal transition through GLP-1R/sirt3 pathway. Biomedicine and Biomedicine & Pharmacotherapy = Biomedecine & Pharmacotherapie, 106, 1364–1369.
- Paola, L., et al., 2013. Exendin-4 induces cell adhesion and differentiation and counteracts the invasive potential of human neuroblastoma cells. Plos One, 8 (8), 903–906.
- Kito, M., et al., 2015. CRP 1846C > T genetic polymorphism is associated with lymph node metastasis and/or severe lymphatic invasion in endometrial cancer. The Tohoku Journal of Experimental Medicine, 237, 25–30.
- Shigeoka, T., et al., 2020. Activation of overexpressed GLP attenuates prostate cancer growth by inhibiting cell cycle progression. Journal of Diabetes Investigation, 11 (5), 1137–1149.
- Sobecki-Rausch, J. and Barroilhet, L., 2019. Anti-programmed death-1 immunotherapy for endometrial cancer with microsatellite instability–high tumors. Current Treatment Options in Oncology, 20 (11), 83.
- Su, Y., et al., 2020. Ferroptosis, a novel pharmacological mechanism of anti-cancer drugs. Cancer Letters, 483, 127–136.
- Suzuki, K., et al., 2019. Identification of 28 new susceptibility loci for type 2 diabetes in the Japanese population. Nature Genetics, 51 (3), 379–386. (Suppl 1).
- Timper, K., et al., 2020. GLP-1 receptor signaling in astrocytes regulates fatty acid oxidation, mitochondrial integrity, and function. Cell Metabolism, 31 (6), 1189–1205.e13.
- Vetter, M. H., et al., 2019. Preoperative predictors of endometrial cancer at time of hysterectomy for endometrial intraepithelial neoplasia or complex atypical hyperplasia. Autonomic Neuroscience: basic & Clinical, 222, 60.e1–60.e7.
- Wang, H., et al., 2021. Silencing of PTPN18 induced ferroptosis in endometrial cancer cells through p-P38-mediated GPX4/xCT down-regulation. Cancer Management and Research, 13, 1757–1765.
- Xiao, Y., et al., 2021. Inhibition of PFKFB3 induces cell death and synergistically enhances chemosensitivity in endometrial cancer. Oncogene, 40 (8), 1409–1424.
- Xing, N. A., et al., 2022. Ferroptosis, necroptosis, and pyroptosis in the tumor microenvironment: Perspectives for immunotherapy of SCLC.
- Zhao, H. J., et al., 2020. Activation of GLP-1 receptor enhances the chemosensitivity of pancreatic cancer cells. Journal of Molecular Endocrinology, 64 (2), 103–113.
- Zhao, H., et al., 2014. Activation of glucagon-like peptide-1 receptor inhibits growth and promotes apoptosis of human pancreatic cancer cells in a cAMP-dependent manner. American Journal of Physiology. Endocrinology and Metabolism, 306 (12), E1431–E1441.
- Zhou, W. J., et al., 2019. Estrogen inhibits autophagy and promotes growth of endometrial cancer by promoting glutamine metabolism. Cell Communication and Signaling, 17 (1), 99.
- Zhu, X., et al., 2019. EMT-mediated acquired EGFR-TKI resistance in NSCLC: mechanisms and strategies. Frontiers in Oncology, 9, 1044.