

Abstract
Background
Abnormal stromal-epithelial cell communication is a pathogenic mechanism in endometriosis, and metformin can modulate it. Insulin-like growth factor binding protein-1 (IGFBP1) plays a role in endometriosis, but the exact mechanism is unknown. IGFBP1 is reportedly a downstream target of metformin in some diseases. We aimed to investigate the role of IGFBP1 in endometriosis development, whether it is associated with abnormal communication, and whether metformin affects IGFBP1 expression.
Methods
Patients who underwent surgical treatment for endometriosis or other diseases were enrolled. Ten patients with ovarian-type endometriosis and eight patients each who underwent surgical treatment for other lesions with or without endometriosis were selected, and their tissues taken for cell proliferation, western blotting, polymerase chain reaction, and knockdown experiments.
Results
Ectopic and eutopic stromal cells (EcSCs and EuSCs) lost their ability to inhibit epithelial cell proliferation, and IGFBP1 expression was downregulated in both groups of stromal cells compared to that in normal stromal cells (NSCs; 1.09 vs. 0.25, p = .0002 1.09 vs. 0.57, p = .0029). In an EcSC IGFBP1 overexpression model, the ability of EcSCs to inhibit epithelial cell proliferation was enhanced (EdU positivity decreased from 38% to 25%, p = .0001). Furthermore, adenosine 5’-monophosphate-activated protein kinase (AMPK) phosphorylation was downregulated in EcSCs and EuSCs compared to that in NSCs (0.99 vs. 0.42, p = .0006/0.99 vs. 0.57, p = 0.0032). Treatment of EcSCs with metformin increased AMPK phosphorylation (0.47 vs. 1.04, p = .0107) while upregulating IGFBP1 expression (0.69 vs. 1.01, p = .0164), whereas pre-treatment with an AMPK phosphorylation inhibitor abrogated metformin-induced IGFBP1 upregulation.
Conclusions
IGFBP1 mediates aberrant stromal-epithelial communication in endometriosis. Metformin can upregulate IGFBP1 expression in EcSCs by activating AMPK, and upregulated IGFBP1 enhances the inhibition of epithelial cell proliferation. IGFBP1 is expected to be a therapeutic target for endometriosis.
PLAIN LANGUAGE SUMMARY
Insulin-like growth factor binding protein 1 (IGFBP1) is a protein that regulates cell growth and proliferation and is expressed at abnormal levels in patients with endometriosis. In some cases, metformin has been shown to modulate the expression of this protein. Here, we investigated the role of IGFBP1 in endometriosis development, whether it is associated with abnormal communication, and whether metformin affects IGFBP1 expression in endometrial cells. We found that downregulation of IGFBP1 in endometriosis diminished the ability of stromal cells to inhibit the proliferation of epithelial cells through inhibition of the protein kinase B and extracellular regulated protein kinase pathways. In addition, metformin upregulated IGFBP1 expression by activating adenosine 5’-monophosphate-activated protein kinase, suggesting that IGFBP1 may be one of the potential targets for drug therapy for endometriosis.
Introduction
Approximately, 190 million women globally are affected by endometriosis during their lifetime (at least until menopause) according to the World Bank’s 2017 population estimates (Horne and Missmer Citation2022). Endometriosis affects metabolism in various tissues, including the liver and adipose tissues, leading to systemic inflammation (Taylor et al. Citation2021). A progressive understanding of the disease’s nature has expanded treatment options, and the potential of some drugs for treating endometriosis is being explored.
The growth and differentiation of healthy endometrial epithelial cells are regulated by stromal cells (Arnold et al. Citation2001), while a loss of this ability has been implicated in endometriosis. Some factors secreted by ectopic stromal cells, including Wnt family member 2 (Wnt2), are involved in this aberrant communication (Zhang et al. Citation2015).
Insulin-like growth factor-binding protein 1 (IGFBP1) is associated with endometriosis development, although the exact mechanism remains unclear. This study aimed to elucidate this mechanism. IGFBP1 is primarily secreted by stromal cells of the secretory endometrium and decreases during endometriosis (Meola et al. Citation2010, Shih et al. Citation2022). It competes with the insulin-like growth factor (IGF) receptor for IGFs, affecting cell proliferation, growth, differentiation, apoptosis, migration, invasion, and adhesion in normal and tumour cells (Lin et al. Citation2021). During gestation, decidualised endometrial stromal cells highly express IGFBP1, which regulates the growth of extravillous trophoblast cells, a highly migratory cell subset important in embryogenesis. Trophoblasts are more invasive in vitro, wherein IGFBP1 expression is inhibited (Irwin et al. Citation1999). Progesterone induces IGFBP1 expression by endometrial stromal cells in animals. High IGFBP1 levels impede endometrial epithelial cell proliferation by suppressing IGF action (Murphy and Ghahary Citation1990, Seppälä et al. Citation1994). Excess IGFBP1 impairs the rapid division of breast cancer cells by inhibiting the interaction between IGF1 and IGF receptor 1 (Figueroa et al. Citation1993). We hypothesised that abnormal IGFBP1 expression in stromal cells in individuals with endometriosis causes abnormal stromal–epithelial cell communication. We examined whether abnormal IGFBP1 levels contribute to the diminished ability of stromal cells to regulate epithelial cells.
Metformin can target Wnt2 and thereby alleviate abnormal stromal–epithelial communication (Zhang et al. Citation2010). Metformin increases serum IGFBP1 levels in patients with polycystic ovary syndrome (Pawelczyk et al. Citation2004). In lung cancer, metformin potentiates the effect of solamargine in upregulating IGFBP1 expression (Tang et al. Citation2017b). Thus, we aimed to examine the potential effect of metformin on IGFBP1 expression in endometrial stromal cells and elucidate its underlying mechanism.
Methods
Materials
Collagenase IA and metformin were purchased from Sigma–Aldrich Ltd. (St Louis, MO, USA). Trypsin, Dulbecco’s modified Eagle’s medium/nutrient mixture F-12 (DMEM/F12; 1:1) medium, and charcoal-stripped foetal bovine serum (FBS) were purchased from Gibco (Billings, MT, USA). Rabbit anti-human IGFBP1, AMPK, phospho-AMPK, protein kinase B (AKT), phospho-AKT, extracellular regulated protein kinase (ERK), and phospho-ERK were purchased from Abcam (Cambridge, MA, USA). Rabbit anti-human β-actin and GAPDH primary antibodies and goat anti-rabbit horseradish peroxidase-conjugated secondary antibodies were purchased from Proteintech (Wuhan, China). Compound C (an AMPK inhibitor) was purchased from MedChemExpress (Shanghai, China).
Patients and tissue samples
This was an experimental study in which ectopic and eutopic endometrium of ovarian-type endometriosis and normal endometrium of non-endometriosis were collected from patients who underwent surgical treatment from June 2022 to June 2023 at the Department of Gynaecology of Shandong Provincial Hospital. Endometriosis was defined as the presence of endometrial-like tissue outside the uterine cavity, and ovarian-type endometriosis was defined as ectopic endometrium colonising the ovaries.
The inclusion and exclusion criteria were as follows:
Inclusion criteria: (1) patients with ovarian-type endometriosis clearly diagnosed by pathology, patients diagnosed with ovarian-type endometriosis without comorbidity of oestrogen-dependent diseases, such as adenomyosis, uterine fibroids, endometrial polyps, and endometrial cancer, and non-endometriosis patients without the combination of the above oestrogen-dependent diseases (control group).
(2) All participants were premenopausal and had regular menstrual cycles. All samples were collected during the secretory phase of the menstrual period based on menstrual history and histological assessment.
Exclusion criteria: (1) Combination of oestrogen-progestin dysregulation caused by other reproductive endocrine-related diseases, such as polycystic ovary syndrome.
(2) Comorbid oestrogen-related malignant tumours, such as endometrial cancer and breast cancer or previous relevant medical history.
(3) Received hormone therapy within 6 months prior to surgery.
(4) Taking hormonal contraceptives within 6 months prior to surgery.
(5) Hormonal birth control device placed in the uterine cavity.
A total of 26 patients were included in this experiment; the experimental group contained 18 patients with ovarian-type endometriosis. The ectopic endometrium located in the ovaries of 10 patients with ovarian-type endometriosis and the intrauterine eutopic endometrium of 8 patients with ovarian-type endometriosis were collected; the normal endometrium of eight patients with non-endometriosis (control group) was also collected. The mean age of the experimental group was 43.23 ± 5.07 years (mean ± standard deviation), and that of the control group was 44.83 ± 5.62 years (mean ± standard deviation).
Primary cell culture
Endometrial cells were isolated as previously described (Zhang et al. Citation2010). Endometriotic cysts in the ovaries (ectopic stromal cells [EcSCs]), eutopic stromal cells (EuSCs; endometriomas), and normal stromal cells (NSCs; patients without endometriosis) were cultured in complete culture media (DMEM/F12 [1:1]) supplemented with 10% charcoal-stripped FBS and 1% penicillin/streptomycin (HyClone, Logan, UT, USA) at 37 °C with 5% CO2. The medium was replaced every 2–3 d. The cultured cells were identified using immunohistochemistry with mouse anti-human vimentin antibodies (Zhang et al. Citation2010). The purity of stromal cells was >98%.
A well-differentiated human endometrioid adenocarcinoma epithelial cell line (Ishikawa; Central Laboratory of Shandong Provincial Hospital) was grown in a complete culture medium.
Cell proliferation assay
The 5-ethynyl-2’-deoxyuridine (EdU) assay was performed to assess the effect of IGFBP1 overexpression and metformin treatment on Ishikawa cell proliferation, using the Cell Light EdU Apollo 567 in Vitro Imaging Kit (RiboBio Co., Ltd., Guangzhou, China) following the manufacturer’s instructions. By removing the upper layer of the non-contact co-culture system, proliferation in the lower layer was assayed. Images were captured using a ZEISS inverted fluorescence microscope (ZEISS, Germany), and the results were analysed using the ImageJ software (version 2.3.0, https://imagej.nih.gov/).
HTS Transwell-96 plates with 0.4-µm pore polyester membrane inserts (Corning, NY, USA) were used in Cell Counting. ISK cells (3 × 103/well) were seeded in 96-well plates. The stromal cells were added to the upper chamber to co-culture with ISK cells for specific times (0, 24, 48, and 72 h), and then, Cell Counting Kit-8 (MedChemExpress, Shanghai, China) was used for incubation at 2 h in the dark. The absorbance value was measured at 450 nm.
Enzyme-linked immunosorbent assay (ELISA)
An ELISA Kit (Elabscience Biotechnology, Wuhan, China) was used to determine IGFBP1 levels secreted by NSCs, EuSCs, and EcSCs following the manufacturer’s instructions with supernatants from cells cultured for 72 h. The mean of duplicate readings for the standard and sample was calculated, and the average zero standard optical density was deducted from the readings. A four-parameter logistic curve was plotted on a log–log axis, with standard concentrations and optical density values on the x- and y-axes, respectively, to determine sample concentrations.
Transfection
Lentiviral vectors for IGFBP1 overexpression and a negative control lentivirus, both expressing eGFP, were purchased from GeneChem (Shanghai, China). One group of cells was transfected with the lentivirus overexpressing IGFBP1 (IGFBP1-LV group), whereas the other was transfected with an empty lentivirus (NC–LV group). The multiplicity of infection was 50% in both groups. Lentivirus was introduced to the medium the following day when the cells had reached 40% confluence. The lentivirus-containing medium was replaced with fresh media after 24 h. After 72 h, the transfection efficiency was estimated using an inverted phase/fluorescence microscope (ZEISS, Germany).
Non-contact co-culture in transwell plates
We used stromal and Ishikawa cells from passages 1–3. Regular 24-well and Transwell plates with 0.4-µm pore polyester membrane inserts were purchased from Corning (NY, USA). Stromal cells were seeded into regular twenty-four-well plates and cultured in serum-free DMEM/F-12 (1:1). After adhesion, the cells were pre-treated for 3 d with DMEM/F12 (1:1) containing 2% FBS and 20 μM metformin; based on clinical pharmacokinetics, 20 μM is the accepted clinically equivalent in-vitro dose (Isoda et al. Citation2006, Graham et al. Citation2011). Subsequently, metformin-treated stromal cells were trypsinised and seeded in the upper chamber of a Transwell 24-well culture plate, and Ishikawa cells were seeded in the lower chamber, with 0.1 and 0.6 mL of 2% charcoal-stripped FBS in the upper and lower chambers, respectively. After 3 d, the upper chamber was detached; cell proliferation in the lower chamber was examined using the EdU assay, or proteins were extracted for western blotting.
Total protein extraction and western blotting
Total protein was extracted using radio-immunoprecipitation assay lysis buffer (Beyotime, Shanghai, China) containing 1% PMSF reagent (Beyotime, Shanghai, China) and a 1% phosphatase inhibitor cocktail (MedChemExpress, Shanghai, China). Denatured total proteins were electrophoresed on a 12% polyacrylamide gel and transferred onto polyvinylidene fluoride membranes. Blocking was performed for 1 h in 5% bovine serum albumin (Solarbio, Beijing, China), and membranes were incubated overnight at 4 °C with the indicated rabbit anti-human antibodies (1:3000). Subsequently, the membranes were incubated with HRP-conjugated goat anti-rabbit secondary antibodies (1:5000) for 1 h. The blots were visualised using the BeyoECL Moon chemiluminescence kit (Beyotime, Shanghai, China) on the Amersham Imager 680 imager. Grey values were analysed using the ImageJ software. Band densities were normalised to that of β-actin or GAPDH.
AMPK phosphorylation assay
Cells were inoculated in culture plates and left to adhere before serum starvation in serum-free DMEM/F-12 (1:1) for 16 h. The cells were treated for 1 h with or without 40 μM Compound C (Xue et al. Citation2013); 20 μM metformin was added, and the cells were maintained for 3 d. We determined AMPK phosphorylation and IGFBP1 expression by western blotting and polymerase chain reaction (PCR), respectively.
RNA extraction and reverse transcription quantitative PCR (RT-qPCR)
Total RNA was extracted using the TRIzol reagent. One microgram of RNA was reverse-transcribed into complementary DNA (cDNA) using transcription reagents from Vazyme. Primers for RT-qPCR were designed by Takara Bio (Shiga, Japan), and the homo gene primer sequences are shown in . RT-qPCR was conducted using a 20-μL reaction mixture containing 2 μL of cDNA. IGFBP1 expression was assessed using 2 × ChamQ SYBR qPCR Master Mix (Vazyme, Shanghai, China) on the Roche Applied Science LightCycler 480 II (Roche Ltd, Germany). The internal reference was β-actin, and the relative IGFBP1 expression was evaluated using the 2-ΔΔCT method.
Table 1. Homo gene primers for reverse transcription PCR.
Immunohistochemistry
Paraffin sections of ectopic and eutopic endometrial tissue from patients with ovarian-type endometriosis and normal endometrial tissue from patients without endometriosis were collected. The sections were incubated at 65 °C for 2 h, deparaffinised with xylene, and hydrated with ethanol. Antigen repair was performed using high-pressure boiling, followed by blocking of endogenous peroxidase blocker (ZSGB, Beijing, China) and incubation with anti-IGFBP1 primary antibody (Proteintech, Wuhan, China) (1:200) at 4 °C overnight. Next, sections were incubated with HRP-labelled secondary antibody (ZSGB, Beijing, China) for 30 min. Colour was developed using the diaminobenzidine (DAB) substrate kit (ZSGB, Beijing, China) for 1 to 2 min, and nuclei were re-stained with haematoxylin.
Statistical analysis
Statistical analyses were conducted using the GraphPad Prism software (version 9.0, http://www.graphpad.com). All experiments were independently repeated thrice, and the data are reported as mean ± standard deviation. The normality of the data was assessed using the Shapiro–Wilk test. The test and control groups were compared using Student’s t-test or a one-way analysis of variance followed by Dunnett’s multiple comparison test. Statistical significance was set at p < .05.
Results
NSCs inhibited Ishikawa cell proliferation, whereas EuSCs and EcSCs lost this ability
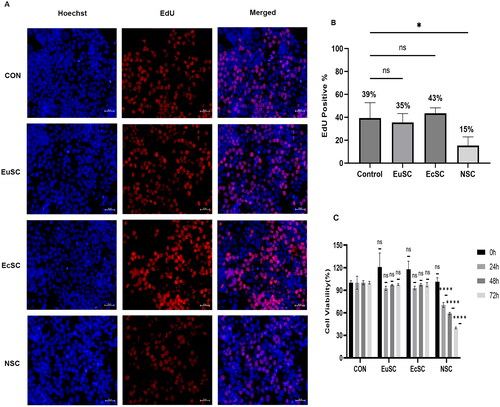
The EdU assay and CCK-8 (expressed as percentages) revealed that EuSCs and EcSCs lost their ability to suppress Ishikawa cell proliferation to varying degrees compared to NSCs ().
Figure 1. Normal stromal cells (NSCs) inhibit the growth of Ishikawa cells but not that of eutopic stromal cells (EuSCs) and ectopic stromal cells (EcSCs) (EdU A and B, CCK-8 C). NSCs exert a suppressive effect on Ishikawa cell proliferation compared to the controls; however, the effect of EcSCs and EuSCs was not significantly different from that of the control (p > .05). Scale bar: 50 μm. Values are mean ± SD (n = 3; ns, not significantly different; *, p < .05; ****, p < .0001).
IGFBP1 expression in EuSCs and EcSCs of patients with endometriosis was downregulated
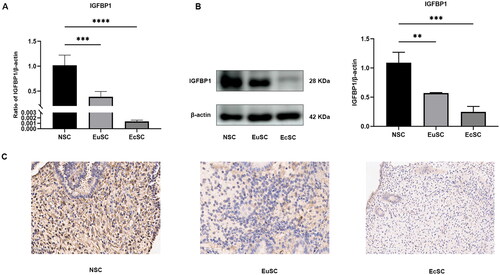
IGFBP1 mRNA and protein levels in EuSCs and EcSCs were lower than those in NSCs (). Further, low IGFBP1 expression levels were detected in both EuSCs and EcSCs ()
Figure 2. IGFBP1 expression in EuSCs and EcSCs of patients with endometriosis was downregulated. (A) IGFBP1 levels in EuSCs and EcSCs were lower than those in NSCs, as determined using RT-qPCR. (B) IGFBP1 levels in EuSCs and EcSCs were lower than those in NSCs, as determined using western blotting. Values are mean ± SD (n = 3; **, p < .01; ***, p < .001; ****, p < .0001). (C) Immunohistochemistry of IGFBP1 expression in both EuSCs and EcSCs.
Quantification of IGFBP1 secretion in stromal cells
ELISA showed that IGFBP1 secretion by NSCs (1.8 ng/mL) was approximately 3.8 and 10 times higher than that in EuSCs (0.47 ng/mL) and EcSCs (0.18 ng/mL), respectively (Fig. S1).
Construction of IGFBP1 overexpression vector
We hypothesised that the effects of differences in stromal cells on Ishikawa cell proliferation are related to IGFBP1 levels. EcSCs with low IGFBP1 expression were selected for the subsequent construction of an IGFBP1 overexpression model. Western blotting and RT-qPCR demonstrated IGFBP1 overexpression in EcSCs (Fig. S2).
High IGFBP1 expression in EcSCs increased their ability to inhibit Ishikawa cell proliferation
We co-cultured IGFBP1-overexpressing EcSCs with Ishikawa cells and performed the EdU assay to assess the effect on cell proliferation. EcSCs with high IGFBP1 expression showed enhanced inhibition of Ishikawa cell proliferation. Metformin-|stimulated and IGFBP1-overexpressing EcSCs inhibited Ishikawa cell proliferation to a similar extent (Fig. S3).
Decreased AMPK phosphorylation in EuSCs and EcSCs
AMPK phosphorylation was reduced in EuSCs and EcSCs, whereas total AMPK protein expression in the two cell types was similar to that in NSCs (Fig. S4).
Metformin treatment increases AMPK phosphorylation in EcSCs and upregulates IGFBP1 expression
Metformin treatment increased AMPK phosphorylation in EcSCs and upregulated IGFBP1 expression (Fig. S5A and B). When AMPK phosphorylation was inhibited using Compound C, metformin lost its ability to increase IGFBP1 expression in these cells.
Metformin inhibits ERK and AKT phosphorylation in Ishikawa cells through IGFBP1 upregulation in EcSCs
To explore whether metformin inhibited epithelial cell proliferation through IGFBP1 expressional upregulation, we performed further experiments using Ishikawa cells. Western blotting showed consistent inhibition of AKT and ERK phosphorylation (Fig. S6).
Overall, stromal cells in patients with endometriosis had a reduced ability to inhibit the growth of epithelial cells than those from patients with no endometriosis. These cells showed differences in the levels of IGFBP1 and AMPK phosphorylation; specifically, expression was low in EuSCs and EcSCs. Metformin stimulated AMPK phosphorylation and upregulated IGFBP1 expression in EuSCs and EcSCs. EcSCs stimulated with metformin or IGFBP1 overexpression inhibited the AKT and ERK pathways in epithelial cells and enhanced the inhibitory effect on their growth.
Discussion
We found lower IGFBP1 expression in eutopic and ectopic endometriosis endometrium compared to normal endometrial tissues, especially in ectopic endometrium. Downregulating IGFBP1 in ectopic endometrial stromal cells reduced their ability to inhibit epithelial cell proliferation. Because of the limited growth potential of primary epithelial cells, we used the Ishikawa cell line (Guzel et al. Citation2011). When lentivirus was used to overexpress IGFBP1 in ectopic stromal cells, their ability to inhibit epithelial cell growth was enhanced. Further mechanistic studies revealed that IGFBP1 acts by inhibiting AKT and ERK-related proliferative signalling pathways.
Metformin is an insulin sensitiser that plays a role in modulating IGFBP1 levels (Tang et al. Citation2017b). Therefore, we treated EcSCs with equivalent in-vitro concentrations of metformin and found that it upregulated IGFBP1. Additionally, metformin is an agonist of AMPK and acts mainly by activating AMPK (Wang and Wei Citation2024). AMPK activation plays an important role in various benign and malignant diseases in humans. It regulates cellular energy metabolism and is also a central node in the regulation of malignant tumour progression (Xu et al. Citation2024). For example, in colon cancer, it is involved in autophagy induction, leading to inhibition of tumour cell growth (Zhou et al. Citation2023). Conversely, its activation may be associated with oncogenesis and drug resistance development (Lin et al. Citation2023). We observed reduced AMPK phosphorylation in both EuSCs and EcSCs, while metformin treatment upregulated IGFBP1 expression and enhanced AMPK phosphorylation. After pre-treatment with an AMPK inhibitor, IGFBP1 upregulation disappeared, while metformin lost its role in activating AMPK phosphorylation. Therefore, we conjectured that metformin might exert its up-regulatory effect on IGFBP1 by activating AMPK.
Ursolic acid and rhodopsin inhibit the growth of cancer cells, such as hepatocellular carcinoma and lung cancer cells, by increasing IGFBP1 expression (Yang et al. Citation2016, Tang et al. Citation2017a). High IGFBP1 expression correlates with good recurrence-free survival in patients with breast cancer (Wang et al. Citation2019). IGFBP1 levels decrease in the follicular fluid of patients with endometriosis, and its extent is linked to the severity of the condition (Cunha-Filho et al. Citation2003). Our findings are consistent with previous findings that aberrant IGFBP1 expression in patients with endometriosis is involved in aberrant stromal–epithelial cell communication (stromal cells have a diminished ability to inhibit epithelial cells). Increasing IGFBP1 levels inhibited epithelial cell proliferation.
The PI3K/AKT and MEK/ERK pathways mediate cell metabolism, proliferation, survival, and angiogenesis. Activation of the PI3K/AKT, mTOR, and Ras/Raf/MEK/ERK pathways promotes the metabolic activity of cancer cells (Asati et al. Citation2016). Inhibition of AKT-related pathways reduces endometrial epithelial cell proliferation (Yoo et al. Citation2018), and the MEK/ERK1/2 signalling pathway is involved in regulating endometrial epithelial cell growth (Chen et al. Citation2018). We assessed whether inhibition of ectopic epithelial cells by IGFBP1 was related to these pathways. Metformin treatment or IGFBP1 overexpression in stromal cells inhibited AKT and ERK phosphorylation in Ishikawa cells, suggesting that IGFBP1-mediated inhibition of epithelial cell growth occurs via both ATK- and ERK-associated pathways.
This study has some limitations. Our experiments were performed only in primary cells and cell lines derived from patients with endometriosis or endometrial carcinoma and non-endometrial diseases and were not validated in animal models. We plan to address this in the future. Moreover, we did not explore the specific targets of IGFBP1 in epithelial cells. Furthermore, our study is limited to theory and has not been confirmed in clinical practice; hence, clinical correlation analysis between metformin treatment and pathological parameters of endometriosis is lacking.
In summary, IGFBP1 secretion from endometrial cells could influence epithelial cell proliferation. Metformin upregulated IGFBP1 expression in stromal cells by activating AMPK, which in turn exerted antiproliferative effects on the endometriotic epithelium (Fig. S7).
This study revealed another mechanism through which metformin affects stromal–epithelial crosstalk in endometriosis, laying a foundation for clinical research on the therapeutic potential of metformin in managing endometriosis. In addition, IGFBP1 is expected to be another important target for the treatment of endometriosis.
Ethics statement
All women recruited provided written informed consent, and the utilisation of human tissues was approved by the Ethics Review Board of Shandong Provincial Hospital, following the Declaration of Helsinki (Approval number: SWYX: NO. 2023-356).
Authorship contribution statement
Changzhong Li conceived the study, Xuping Shao designed and conducted the experiments, analysed the data, and completed the first draft of the article, and Junhui Liang helped in collecting human tissues and participated in the revision of the manuscript. All authors have reviewed and approved the final manuscript.
Supplemental Material
Download Zip (18.9 MB)Acknowledgements
We thank all individuals who supported this study and participated in multiple revisions of the manuscript. We would like to thank Editage for English language editing.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Data availability statement
Data will be made available on request.
Additional information
Funding
References
- Arnold, J.T., et al., 2001. Endometrial stromal cells regulate epithelial cell growth in vitro: a new co-culture model. Human Reproduction (Oxford, England), 16 (5), 1–7.
- Asati, V., et al., 2016. PI3K/Akt/mTOR and Ras/Raf/MEK/ERK signaling pathways inhibitors as anticancer agents: Structural and pharmacological perspectives. European Journal of Medicinal Chemistry, 109, 314–341.
- Chen, Q., et al., 2018. USP10 promotes proliferation and migration and inhibits apoptosis of endometrial stromal cells in endometriosis through activating the Raf-1/MEK/ERK pathway. American Journal of Physiology. Cell Physiology, 315 (6), C863–c872.
- Cunha-Filho, J.S., et al., 2003. Insulin-like growth factor (IGF)-1 and IGF binding protein-1 and -3 in the follicular fluid of infertile patients with endometriosis. Human Reproduction (Oxford, England), 18 (2), 423–428.
- Figueroa, J.A., et al., 1993. Recombinant insulin-like growth factor binding protein-1 inhibits IGF-I, serum, and estrogen-dependent growth of MCF-7 human breast cancer cells. Journal of Cellular Physiology, 157 (2), 229–236.
- Graham, G.G., et al., 2011. Clinical pharmacokinetics of metformin. Clinical Pharmacokinetics, 50 (2), 81–98.
- Guzel, E., et al., 2011. Bidirectional interaction between unfolded-protein-response key protein HSPA5 and estrogen signaling in human endometrium. Biology of Reproduction, 85 (1), 121–127.
- Horne, A.W. and Missmer, S.A., 2022. Pathophysiology, diagnosis, and management of endometriosis. BMJ (Clinical Research ed.), 379, e070750.
- Irwin, J.C., et al., 1999. Role of the IGF system in trophoblast invasion and pre-eclampsia. Human Reproduction (Oxford, England), 14 Suppl 2, 90–96.
- Isoda, K., et al., 2006. Metformin inhibits proinflammatory responses and nuclear factor-kappaB in human vascular wall cells. Arteriosclerosis, Thrombosis, and Vascular Biology, 26 (3), 611–617.
- Lin, X., et al., 2023. KIAA1429 promotes tumorigenesis and gefitinib resistance in lung adenocarcinoma by activating the JNK/MAPK pathway in an m(6)A-dependent manner. Drug Resistance Updates: Reviews and Commentaries in Antimicrobial and Anticancer Chemotherapy, 66, 100908.
- Lin, Y.W., et al., 2021. IGFBP-1 in cancer: expression, molecular mechanisms, and potential clinical implications. American Journal of Translational Research, 13 (3), 813–832.
- Meola, J., et al., 2010. Differentially expressed genes in eutopic and ectopic endometrium of women with endometriosis. Fertility and Sterility, 93 (6), 1750–1773.
- Murphy, L.J. and Ghahary, A., 1990. Uterine insulin-like growth factor-1: regulation of expression and its role in estrogen-induced uterine proliferation. Endocrine Reviews, 11 (3), 443–453.
- Pawelczyk, L., et al., 2004. Metformin therapy increases insulin-like growth factor binding protein-1 in hyperinsulinemic women with polycystic ovary syndrome. European Journal of Obstetrics, Gynecology, and Reproductive Biology, 113 (2), 209–213.
- Seppälä, M., et al., 1994. Uterine endocrinology and paracrinology: insulin-like growth factor binding protein-1 and placental protein 14 revisited. Human Reproduction (Oxford, England), 9 (5), 917–925.
- Shih, A.J., et al., 2022. Single-cell analysis of menstrual endometrial tissues defines phenotypes associated with endometriosis. BMC Medicine, 20 (1), 315.
- Tang, Q., et al., 2017a. Emodin increases expression of insulin-like growth factor binding protein 1 through activation of MEK/ERK/AMPKalpha and interaction of PPARgamma and Sp1 in lung cancer. Cellular Physiology and Biochemistry: International Journal of Experimental Cellular Physiology, Biochemistry, and Pharmacology, 41 (1), 339–357.
- Tang, Q., et al., 2017b. Combination of solamargine and metformin strengthens IGFBP1 gene expression through inactivation of Stat3 and reciprocal interaction between FOXO3a and SP1. Cellular Physiology and Biochemistry: International Journal of Experimental Cellular Physiology, Biochemistry, and Pharmacology, 43 (6), 2310–2326.
- Taylor, H.S., et al., 2021. Endometriosis is a chronic systemic disease: clinical challenges and novel innovations. Lancet (London, England), 397 (10276), 839–852.
- Wang, J., et al., 2019. The prognostic values of insulin-like growth factor binding protein in breast cancer. Medicine, 98 (19), e15561.
- Wang, Q. and Wei, X., 2024. Research progress on the use of metformin in leukemia treatment. Current Treatment Options in Oncology, 25 (2), 220–236.
- Xu, X., et al., 2024. Regulation of AMPK activation by extracellular matrix stiffness in pancreatic cancer. Genes & Diseases, 11 (3), 101035.
- Xue, J., et al., 2013. Metformin inhibits growth of eutopic stromal cells from adenomyotic endometrium via AMPK activation and subsequent inhibition of AKT phosphorylation: a possible role in the treatment of adenomyosis. Reproduction, 146 (4), 397–406.
- Yang, L.J., et al., 2016. Inter-regulation of IGFBP1 and FOXO3a unveils novel mechanism in ursolic acid-inhibited growth of hepatocellular carcinoma cells. Journal of Experimental & Clinical Cancer Research, 35 (1), 59.
- Yoo, J.Y., et al., 2018. MIG-6 suppresses endometrial epithelial cell proliferation by inhibiting phospho-AKT. BMC Cancer, 18 (1), 605.
- Zhang, H., et al., 2010. Endometriotic epithelial cells induce MMPs expression in endometrial stromal cells via an NFkappaB-dependent pathway. Gynecological Endocrinology, 26 (6), 456–467.
- Zhang, H., et al., 2015. Metformin regulates stromal-epithelial cells communication via Wnt2/beta-catenin signaling in endometriosis. Molecular and Cellular Endocrinology, 413, 61–65.
- Zhou, X., et al., 2023. Rigosertib promotes anti-tumor immunity via autophagic degradation of PD-L1 in colorectal cancer cells. Cancer Letters, 577, 216422.