

 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.Abstract
Background
Existing treatments for primary dysmenorrhoea (PD), such as NSAIDs, impart side effects. Ge-Gen decoction (GGD), a traditional Chinese medicine, has shown promise in treating PD, but its exact mechanisms remain unclear. Here, we aimed to investigate the efficiency of GGD in alleviating PD using a rat model to understand its precise mechanism of action.
Methods
We established a rat model of dysmenorrhoea induced by oestradiol and oxytocin. The PD rats were administered GGD or Ibuprofen (positive control) intragastrically once daily for seven consecutive days. Serum levels of prostaglandin E2 (PGE2), prostaglandin F2 alpha (PGF2α), β-endorphin (β-EP), thromboxane B2 (TXB2), 6-keto-prostaglandin F1α (6-keto-PGF1α) were determined using an enzyme-linked immunosorbent assay (ELISA). The expression levels of oestrogen receptor alpha (ERα) and cyclooxygenase-2 (COX-2) in uterine tissue were measured using immunohistochemical assays, and those of phosphorylated and total extracellular signal-regulated protein kinases 1 and 2 (ERK1/2) were assessed using western blot analysis.
Results
Treatment with GGD significantly reduced writhing behaviour, histopathological scores, and levels of COX-2, PGE2, and PGF2α in the serum of PD rats. Additionally, GGD increased β-EP content and inhibited ERK1/2 activation and ERα expression in uterine tissues.
Conclusions
The results of this study suggest that GGD alleviates PD in rats by suppressing the COX-2-mediated release of PGE2 and PGF2α, modulating the ERα/ERK1/2/COX-2 pathway, and increasing β-EP content. These results provide insights into the potential mechanisms of GGD in treating PD and support its further investigation as an alternative therapy for this condition.
PLAIN LANGUAGE SUMMARY
Ge-Gen decoction is commonly used to alleviate primary dysmenorrhoea. However, its anti-dysmenorrhoea mechanism remains elusive. In this study, using a rat model of primary dysmenorrhoea, we demonstrate that Ge-Gen decoction reduced the levels of cyclooxygenase-2, prostaglandin E2, and prostaglandin F2 alpha in serum and phosphorylated extracellular signal-regulated protein kinases 1 and 2 in the uterus. These results suggest that Ge-Gen decoction alleviates primary dysmenorrhoea via inactivation of the oestrogen receptor alpha/extracellular signal-regulated protein kinases 1 and 2/cyclooxygenase-2 pathway. This study enhances our understanding of the pathogenesis of primary dysmenorrhoea and may potentially inform the development of novel treatment approaches.
Graphical Abstract
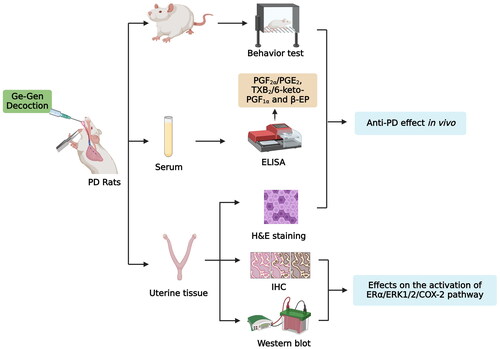
Introduction
Primary dysmenorrhoea (PD) is a prevalent and debilitating gynaecological problem characterised by menstrual pain without evident pelvic abnormalities (Tu and Hellman Citation2021). Traditionally, PD is treated with NSAIDs, which often prove ineffective and lead to adverse effects such as drowsiness, indigestion, and headache (Tu and Hellman,Citation2021 Ahmadi et al. Citation2022). To overcome these limitations, traditional Chinese medicines (TCM), which exert minimal adverse effects, have been recognised as an effective alternative approach to alleviate the symptoms of PD (Leem et al. Citation2019, Chai et al. Citation2020, Li et al. Citation2021, 2022 Ma et al.Citation2021, Yuan et al. Citation2021).
Ge-Gen decoction (GGD) is a polyherbal TCM comprising various concentrations of Gegen (Pueraria lobata), Ephedrae (Ephedra sinica), Cinnamon Twig (Cinnamomum cassia), Bai Shao (Paeonia lactiflora), ginger (Gingiber officinale), Glycyrrhizae Radix Et Rhizome (Glycyrrhiza uralensis), and Red Dates (Ziziphus jujuba). A previous clinical study has demonstrated its beneficial effects against PD without inducing any significant adverse effects (Chai et al. Citation2020). In rat models of PD, GGD has been shown to markedly reduce uterine tetanic contraction and increase uterine artery blood velocity, possibly involving prostaglandin F2α (PGF2α) and calcium signalling (Yang et al. Citation2016). However, the precise mechanisms underlying the therapeutic effects of GGD against PD remain unclear.
Increased intrauterine secretion of PGs is responsible for pelvic pain associated with PD (Tu and Hellman Citation2021). Cyclooxygenase-2 (COX-2) is a key enzyme that catalyses the rate-limiting step in converting arachidonic acid to PGs (Ahmadi et al. Citation2022). COX-2 expression is regulated by extracellular signal-regulated protein kinases 1 and 2 (ERK1/2) signalling (Guo et al. Citation2020, Nakano et al. Citation2020, Nakayama et al. Citation2022), which is activated and expressed in PD (Tong et al. Citation2021, Liu et al. Citation2022, Zhao et al. Citation2022). Increasing evidence suggests that COX-2-mediated secretion of PGs is crucial for the development of PD (Tu and Hellman,Citation2021 Ahmadi et al. Citation2022). However, it remains unclear whether ERK1/2 is responsible for COX-2-mediated PG secretion in patients with PD. Oestrogen receptor alpha (ERα) is a nuclear transcription factor responsible for maintaining oestrogen homeostasis in the uterus to ensure physiological responsiveness (Wright and Lannigan Citation2023). Moreover, the ERK1/2 pathway is involved in the regulation of ERα-mediated responses (Lannigan,Citation2022 Wright and Lannigan Citation2023). Therefore, we hypothesised that the ERα/ERK1/2/COX-2 pathway may play a role in the pathogenesis of PD, and therapeutic interventions targeting this pathway may exert protective effects against PD. However, the mechanistic link between the ERα/ERK1/2/COX-2 pathway and the anti-PD activity of GGD remains unexplored.
To test this hypothesis, in this study, we aimed to elucidate the mechanistic link between the ERα/ERK1/2/COX-2 pathway and the anti-PD activity of GGD and explore the potential therapeutic effects of targeting this pathway. The findings of this study will provide mechanistic insights into the understanding of the pathogenesis and inflammatory processes in the pelvic region and potentially aid the development of novel treatment approaches.
Methods
GGD preparation
The GGD preparation comprised Paeonia lactiflora (15 g), Ephedrae (5 g), Pueraria lobata (20 g), Cinnamon Twig (15 g), ginger (10 g), Glycyrrhizae Radix Et (10 g), and Red Dates (20 g), and Rhizome (10 g). All materials were obtained from the Taicang TCM Hospital, and GGD was prepared as described in a previous study (Xie et al. Citation2020). Briefly, the materials were soaked in 1000 mL water for 30 min and then decocted with boiling water twice for 2 h each time. Finally, the extracts were pooled, filtered, incubated in a water bath at 60 °C, and evaporated to 100 mL. The final extract was transferred to a bottle and stored at 4 °C for further use.
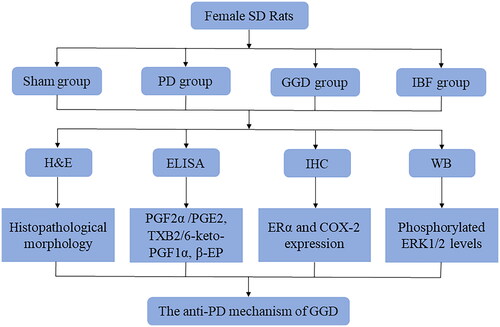
Animals and treatment
A total of 24 female Sprague Dawley (SD) 8-week-old rats weighing 200–250 g were obtained from Huachuang Sino Co., Ltd. (Taizhou, China). The rats were housed in a controlled environment with a 12-h light/12-h dark cycle at a temperature of 22 ± 2 °C and humidity of 45 ± 10%. All animal experiments were approved by the Ethics Committee of Taicang Hospital of Traditional Chinese Medicine (approval number: 2021-031).
The PD model of rats was established as described in a previous study (Xie et al. Citation2020). Each rat was subcutaneously injected with oestradiol benzoate once a day for 10 consecutive days; on days 1 and 10, the dose was 0.5 mg/rat, while on days 2–9, the dose was reduced to 0.2 mg/rat. On the 11th day of modelling, each rat was given an intraperitoneal injection of oxytocin (2 U/rat). After a one-week adaptation, 24 rats were randomly divided into four groups (n = 6 rats/group): sham, PD, GGD, and ibuprofen. The rats in the GGD and ibuprofen groups were administered GGD (1.8 mg/g) and Ibuprofen (0.06 mg/g) solubilised in distilled water once daily via the intragastric route. Rats in the PD and sham groups received an equivalent amount of distilled water once daily. The flowchart of the study is shown in .
Figure 1. A flow chart of this study. SD: Sprague Dawley; PD: Primary dysmenorrhoea; GGD: Ge-Gen decoction; IBF: Ibuprofen; H&E: Haemotoxylin and Eosin; ELISA: Enzyme-linked immunosorbent assay; IHC: Immunohistochemistry; WB: Western blot; PGE2: Prostaglandin E2; PGF2α: Prostaglandin F2 alpha; ERα: Oestrogen receptor alpha; ERK1/2: Extracellular signal-regulated protein kinases 1 and 2; COX-2: Cyclooxygenase-2.
Determination of writhing reaction
Writhing frequency and latency were measured 30 min after oxytocin administration through careful observation of the twists and contortions of the physique of each rat.
Histopathological examination
Uterine tissues were washed in phosphate buffer, fixed in 5% formalin at room temperature, dehydrated in a graded concentration of ethanol, and then embedded in paraffin. Tissue sections of 4 μm were stained with haematoxylin and eosin (H&E) to evaluate the histopathological changes. Digital images of uterine morphology were obtained using a light microscope (Olympus CKX53, Tokyo, Japan). The morphological changes in the uterus of the rats were graded as 0–3: 0, no pathological observations [the uterine structure is clear, with a three-layer structure, including the endometrium, myometrium, and adventitia from inside to outside. The endometrial epithelial cells are complete, the distribution of glands in the lamina propria is normal, and the thickness of the endometrium and myometrium is normal] and 3, severe pathological damage [the uterine structure is disordered, the uterine cavity is not smooth, gland distribution in lamina propria is abnormal, gland hypertrophy, endometrial hyperplasia is obvious, myometrial hyperplasia is obvious, and inflammatory cell infiltration is obvious].
Immunohistochemistry (IHC)
The expression of COX-2 and ERα in uterine tissues was determined using IHC. The tissue sections were incubated at 4 °C with anti-COX2 and anti-ERα primary immunoglobulins for 12 h, followed by incubation with the appropriate secondary immunoglobulins for 1 h. After staining the sections with DAB and counterstaining with haematoxylin, images were acquired using a scanner (Pannoramic MIDI, 3D histech, Hungary). Image-Pro Plus software (version 6.0) was used to quantify the integrated optical density (IOD) and average optical density (AOD) based on IOD as follows:
Western blot assay
Western blotting was performed as previously described (Xie et al. Citation2020). The antibodies used in this study included anti-phosphorylated ERK1/2 (Cat. No. ab201015, 1:1000, Abcam, USA), anti-ERK1/2 (Cat. No. 11257-1-AP, 1:1000, Proteintech, USA), and anti-ERα (Cat. No. 21244-1-AP, 1:1000, Proteintech, USA).
Enzyme-linked immunosorbent assay (ELISA)
Serum levels of PGF2α, PGE2, β-endorphin (β-EP), thromboxane B2 (TXB2), 6-keto-PGF1α in each rat group were measured using commercial ELISA kits: PGF2α (Catalog No. 516011) and PGE2 (Cat. No. 514010) obtained from Cayman Chemicals (MI, USA), β-EP (Cat. No. H221-1-1), TXB2 (Cat. No. H173-1-1), and 6-keto-PGF1α (Cat. No. H204-1) kits obtained from Nanjing Jincheng Bioengineering Institute (Jiangsu, China). All procedures were performed in accordance with the manufacturer’s instructions.
Statistical analysis
All statistical analyses were conducted using the GraphPad Prism software (version 9.0, USA). Data are presented as the mean ± standard deviation (SD). Two-group comparisons were performed using the Student’s t-test, whereas multiple groups were compared using one-way analysis of variance and the Student–Newman–Keuls test. Statistical significance was set at p < 0.05.
Results
GGD suppressed PD
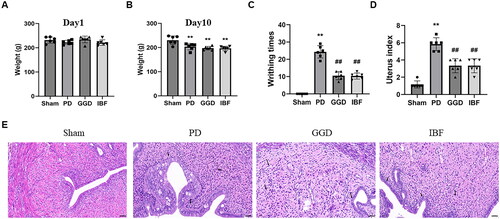
The basal body weight (before treatment administration) did not differ among the groups (), while all groups except for the control group lost weight after 10 days of modelling (, p < 0.01). As shown in , compared with the sham control group, the oxytocin-induced writhing times increased significantly in the PD group (p < 0.01). In contrast, the writhing frequencies were significantly reduced in the GGD and ibuprofen groups compared to that in the PD group (p < 0.01). However, those of the GGD and ibuprofen groups were not comparable (p > 0.05). Furthermore, H&E staining revealed significantly higher histopathological scores in PD rats than in sham rats (p < 0.01), whereas GGD significantly reduced the histopathological score (p < 0.01). These data indicate the suppressive influence of GGD on PD.
Figure 2. The anti-PD effects of GGD on PD model rat. (A) The weight of the rat on day 1 of the experiment. (B) The weight of the rat on day 10. (C–E) The writhing numbers (C), uterine index (D), and pathological changes in the uterine tissue (E) in each group, as determined using H&E staining, scale bar = 50 µM. The black arrow represents the lesion. Data are presented as mean ± SD (n = 6). **, p < 0.01 vs. Sham group, ##, p < 0.01 vs. GGD group (Tukey–Kramer’s test). PD: Primary dysmenorrhoea; GGD: Ge-Gen decoction; IBF: Ibuprofen.
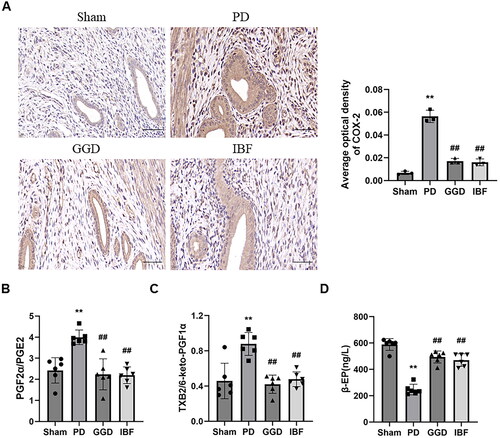
Next, we assessed the effect of GGD on COX-2 levels in PD rat uteri using IHC. As shown in , the relative protein level of COX-2 in the PD group was significantly higher than that in the sham group (p < 0.01). However, the relative protein level of COX-2 in the GGD group was significantly lower than that in the PD group (p < 0.01). Furthermore, the ratio of PGF2α/PGE2 () and TXB2/6-keto-PGF1α () increased significantly in the PD group (p < 0.01), while the ratios decreased significantly in the GGD group (p < 0.01). The β-EP content of the PD group was significantly lower than that of the sham group (; p < 0.01). In contrast, the β-EP content of the GGD group increased significantly (p < 0.01).
Figure 3. Effect of GGD on COX-2 expression and serum biochemical indexes. (A) The protein levels of COX-2 in the uterine tissue of each group, as determined using IHC, scale bar = 20 µM. (B) The ratio of PGF2α/PGE2 and TXB2/6-keto-PGF1α and β-EP content in the serum of rats determined using corresponding ELISA kits. Data are presented as mean ± SD (n = 6). **, p < 0.01 vs. Sham group, ##, p < 0.01 vs. GGD group (Tukey–Kramer’s test). PD: Primary dysmenorrhoea; GGD: Ge-Gen decoction; IBF: Ibuprofen; β-EP: β-endorphin; TXB2, thromboxane B2; 6-keto-PGF1α: 6-keto-prostaglandin F1α; ELISA: Enzyme-linked immunoabsorbent assay.
GGD inhibited ERK1/2 activation in the uterine tissue of PD rats
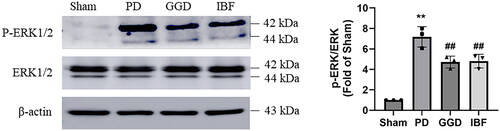
Next, we examined the influence of GGD on ERK1/2 activation in the uterine tissues of PD rats using western blotting. Relative to sham rats, phosphorylated ERK1/2 expression was markedly increased in PD rats, whereas the total ERK1/2 levels remained unchanged (). In contrast, phosphorylated ERK1/2 levels were significantly reduced following GGD administration (, p < 0.01), indicating that GGD inhibited ERK1/2 activation in the uterus of PD rats.
Figure 4. Effect of GGD on the activation of ERK1/2 signalling. The protein levels of phosphorylated ERK1/2 and total ERK1/2 in the uterine tissue of each group were measured with western blotting. Data are presented as mean ± SD (n = 3). **, p < 0.01 vs. Sham group, ##, p < 0.01 vs. GGD group (Tukey–Kramer’s test). PD: Primary dysmenorrhoea; GGD: Ge-Gen decoction; IBF: Ibuprofen.
GGD reduced uterine tissue ERα expression
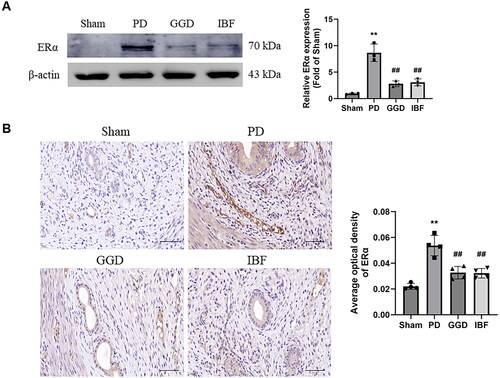
Evaluation of the effects of GGD on ERα levels in the uterine tissues of PD rats revealed significantly higher expression of Erα in the PD group than that in the sham group (; p < 0.01), which was significantly reduced in the GGD group (p < 0.01). Consistently, IHC analysis revealed increased ERα levels in the PD group (, p < 0.01), which were decreased in the GGD group (p < 0.01). These data indicated that uterine ERα expression was reduced by GGD.
Figure 5. Effect of GGD on ERα expression. The protein level of ERα in the uterine tissue of each group, as measured with western blotting (A) and IHC (B). scale bar = 20 µM. Data are presented as mean ± SD (n = 3–4). **, p < 0.01 vs. Sham group, ##, p < 0.01 vs. GGD group (Tukey–Kramer’s test). PD: Primary dysmenorrhoea; GGD: Ge-Gen decoction; IBF: Ibuprofen.
Discussion
This study investigates the effectiveness of GGD in treating PD using a rat model and underscores its mechanism of action. The rat PD model is widely used in research on possible anti-PD mechanisms of TCM (Cheng et al. Citation2018, Zhang et al. Citation2021, Su et al. Citation2022). The ratio of PGF2α/PGE2 is recognised as a diagnostic indicator for PD, while the ratio of TXB2, a metabolite of thromboxane A2 (TXA2) that plays a role in promoting blood clot formation (thrombosis) and 6-keto-PGF1α, a metabolite of prostacyclin (PGI2) that has a vasodilatory effect is associated with blood flow velocity (Zhang et al. Citation2021, Zhao et al. Citation2022). β-EP has a powerful endogenous analgesic effect (Zhang et al. Citation2021, Zhao et al. Citation2022), is influenced by sex hormones, and participates in the regulation of reproductive endocrine (Zhang et al. Citation2021, Zhao et al. Citation2022). In this study, a PD model of rats was successfully established as evidenced by the decrease in β-EP and increase in PGF2α/PGE2, TXB2/6-keto-PGF1α, and the significant increase in writhing time, combined with histopathological changes of uterine tissues. Our results revealed that GGD treatment significantly reduced the frequency of writhes, the histopathological score, the ratio of PGF2α/PGE2 and TXB2/6-keto-PGF1α, and increased β-EP content, indicating that GGD suppressed PD.
PGE2 and PGF2α, the primary members of the PG family, have been implicated in uterine contractility through interaction with their corresponding receptors located on the spiral arterioles (Iacovides et al. Citation2015, Barcikowska et al. Citation2020). PGE2 and PGF2α levels are upregulated in the peripheral blood and uterine tissues of patients with PD (Iacovides et al. Citation2015, Barcikowska et al. Citation2020). Moreover, increasing evidence has demonstrated that high levels of PGE2 and PGF2α lead to pathological pain associated with uterine contractions (Iacovides et al. Citation2015, Barcikowska et al. Citation2020). COX-2, an isoform of COX, is crucial for synthesising PGs and is involved in pain and inflammation processes (Marjoribanks et al. Citation2015, Ahmadi et al. Citation2022). In conditions like PD, COX2 has been shown to regulate the levels of PGE2 and PGF2α, contributing to its pathogenesis (Marjoribanks et al. Citation2015, Ahmadi et al. Citation2022). The findings of this study showed that GGD treatment reduced the levels of PGE2 and PGF2α in the serum of PD rats, suggesting that GGD may alleviate PD by suppressing the COX-2-mediated release of PGE2 and PGF2α.
The ERK1/2 pathway is crucial for the production of inflammatory cytokines and the mediation of oxytocin signalling (Morita et al. Citation2019, Lucas et al. Citation2021, Devost et al. Citation2022, El-Ganainy et al. Citation2022), both of which are implicated in the pathogenesis of PD (Barcikowska et al. Citation2020, Tu and Hellman Citation2021). Studies have demonstrated the upregulation of the ERK1/2 pathway in PD, suggesting a potential therapeutic effect of TCM through its inactivation (Tong et al. Citation2021, Liu et al. Citation2022, Zhao et al. Citation2022). Previous studies have shown that Sclareol, a natural product isolated from Salvia sclarea, alleviated dysmenorrhoea by downregulating the expression of phosphorylated ERK (Wong et al. Citation2020). Furthermore, the ERK1/2 pathway regulates COX-2 expression in various cells, including human endometriotic cells (Guo et al. Citation2020, Nakano et al. Citation2020, Nakayama et al. Citation2022). In this study, GGD reduced the uterine tissue levels of p-ERK1/2, indicating that COX-2 downregulation may be attributed to the inactivation of ERK1/2. However, the exact mechanism by which ERK1/2 mediates this reduction in COX-2 expression in PD rats is not yet understood. A previous study suggested that ERK1/2 may act through the RNA-binding protein p42 AUF1 to regulate COX-2 expression (Zhai et al. Citation2010). Recently, a study in PD mice has shown that uterine COX-2 expression is influenced by NF-κB, a downstream pathway of ERK1/2 signalling (Tang et al. Citation2020). Nevertheless, further investigation is essential to clarify the involvement of p42 AUF1 and NF-κB in the GGD-induced reduction of COX-2. Additionally, COX-2 has been shown to be responsible for the activation of the ERK1/2 pathway (Guo et al. Citation2013), indicating a regulatory loop between ERK1/2 and COX-2. Exploring this regulatory loop in patients with PD after GGD treatment will be of interest.
The nuclear transcription factor ERα has been implicated in the regulation of many complex physiological processes in humans (Fuentes and Silveyra Citation2019). Dysregulated ERα signalling is associated with various diseases, prompting research into potential therapeutic agents that target ERα for the management of different pathological conditions, including inflammation (Lin et al. Citation2020, Cephus et al. Citation2021, Frump et al. Citation2021). In addition, ERα plays a critical role in uterine oestrogen homeostasis, thereby maintaining physiological responsiveness (Wright and Lannigan Citation2023). Studies have indicated that higher ERα expression is linked to moderate to severe dysmenorrhoea and deep dyspareunia, suggesting that symptom severity is influenced not just by circulating oestrogens but also by tissue expression of ERα (Pluchino et al. Citation2020). In the current study, GGD reduced ERα levels in the uterine tissue of PD rats. A previous study has demonstrated that ERα mediates the oestrogen-induced activation of ERK1/2 (Sun et al. Citation2017), and conversely, ERK1/2 regulates ERα-mediated responses (Lannigan,Citation2022 Wright and Lannigan Citation2023). These findings suggest that GGD-mediated inactivation of the ERK1/2 pathway in the PD rat uterus could be due to decreased ERα expression.
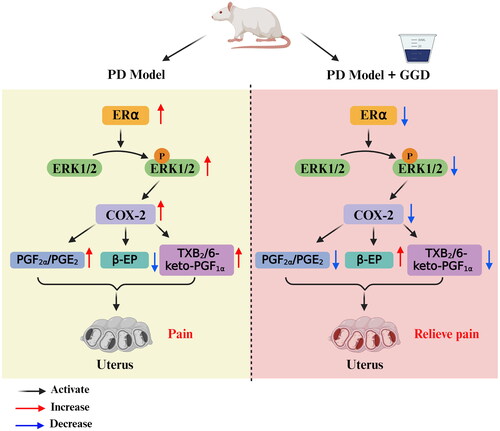
In conclusion, this study has shown that the treatment effects of GGD on PD are associated with the inactivation of the ERα/ERK1/2/COX-2 pathway in the uterus (). These findings provide new insights into the treatment of PD and support the potential use of GGD as an alternative therapy for PD.
Figure 6. Schematic summary illustration. GGD alleviates PD in rats by suppressing the COX-2-mediated release of PGE2 and PGF2α, modulating the ERα/ERK1/2/COX-2 pathway, and increasing β-EP content. PD: Primary dysmenorrhoea; GGD: Ge-Gen decoction; PGE2: Prostaglandin E2; PGF2α: Prostaglandin F2 alpha; ERα: Oestrogen receptor alpha; ERK1/2: Extracellular signal-regulated protein kinases 1 and 2; COX-2: Cyclooxygenase-2, β-EP: β-endorphin; TXB2, thromboxane B2; 6-keto-PGF1α: 6-keto-prostaglandin F1α.
Ethical approval
Animal experiments were approved by the Ethics Committee of Taicang Hospital of Traditional Chinese Medicine (approval number: 2021-031).
Authors’ contributions
Yazhen Xie: Project development, funding acquisition, data analysis, and manuscript editing. Haifeng Xu: Data collection, data analysis, and manuscript writing. Zhijuan Gu: Data collection and analysis. All authors have read and approved the final manuscript.
Data availability statement
The data supporting the findings of this study are available from the corresponding author upon reasonable request.
Additional information
Funding
References
- Ahmadi, M., et al., 2022. Non-steroidal anti-inflammatory drugs: Recent advances in the use of synthetic COX-2 inhibitors. RSC Medicinal Chemistry, 13 (5), 1–9.
- Barcikowska, Z., et al., 2020. Inflammatory markers in dysmenorrhea and therapeutic options. International Journal of Environmental Research and Public Health, 17 (4), 1191.
- Cephus, J.Y., et al., 2021. Estrogen receptor-alpha signaling increases allergen-induced IL-33 release and airway inflammation. Allergy, 76 (1), 255–268.
- Chai, C., et al., 2020. Effect of traditional Chinese medicine formula GeGen decoction on primary dysmenorrhea: A randomized controlled trial study. Journal of Ethnopharmacology, 261, 113053.
- Cheng, Y., et al., 2018. Pharmacokinetic-pharmacodynamic modeling to study the anti-dysmenorrhea effect of Guizhi Fuling capsule on primary dysmenorrhea rats. Phytomedicine: International Journal of Phytotherapy and Phytopharmacology, 48, 141–151.
- Devost, D., Zingg, H.H. and Hébert, T.E., 2022. The MAP kinase ERK5/MAPK7 is a downstream effector of oxytocin signaling in myometrial cells. Cellular Signalling, 90, 110211.
- El-Ganainy, S.O., et al., 2022. Intranasal oxytocin attenuates cognitive impairment, β-amyloid burden and tau deposition in female rats with Alzheimer’s disease: Interplay of ERK1/2/GSK3β/caspase-3. Neurochemical Research, 47 (8), 2345–2356.
- Frump, A.L., et al., 2021. 17beta-estradiol and estrogen receptor alpha protect right ventricular function in pulmonary hypertension via BMPR2 and apelin. Journal of Clinical Investigation, 131 (6), e129433.
- Fuentes, N. and Silveyra, P., 2019. Estrogen receptor signaling mechanisms. Advances in Protein Chemistry and Structural Biology, 116, 135–170.
- Guo, W., et al., 2020. Pyruvate kinase M2 promotes prostate cancer metastasis through regulating ERK1/2-COX-2 signaling. Frontiers in Oncology, 10, 544288.
- Guo, Y.J., et al., 2013. Analgesic effects of the COX-2 inhibitor parecoxib on surgical pain through suppression of spinal ERK signaling. Experimental and Therapeutic Medicine, 6 (1), 275–279.
- Iacovides, S., Avidon, I. and Baker, F.C., 2015. What we know about primary dysmenorrhea today: A critical review. Human Reproduction Update, 21 (6), 762–778.
- Lannigan, D.A., 2022. ERK1/2-RSK2 signaling in regulation of ERalpha-mediated responses. Endocrinology, 163 (9), bqac106
- Leem, J., et al., 2019. Herbal medicine (Hyeolbuchukeo-tang or Xuefu Zhuyu decoction) for treating primary dysmenorrhea: A systematic review and meta-analysis of randomized controlled trials. Medicine, 98 (5), e14170.
- Li, G., et al., 2021. Chinese herbal formula Xuefu Zhuyu for primary dysmenorrhea patients (CheruPDYS): A study protocol for a randomized placebo-controlled trial. Trials, 22 (1), 95.
- Li, N., Li, J. and Gai, P., 2022. Effects of modified Wenjing decoction combined with online publicity and education on the treatment of primary dysmenorrhea of cold coagulation and blood stasis. Journal of Healthcare Engineering, 2022, 1899356.
- Lin, X., Xiang, H. and Luo, G., 2020. Targeting estrogen receptor alpha for degradation with PROTACs: A promising approach to overcome endocrine resistance. European Journal of Medicinal Chemistry, 206, 112689.
- Liu, X., et al., 2022. The mechanism of ginger and its processed products in the treatment of estradiol valerate coupled with oxytocin-induced dysmenorrhea in mice via regulating the TRP ion channel-mediated ERK1/2/NF-kappaB signaling pathway. Food and Function, 13 (21), 11236–11248.
- Lucas, R.M., et al., 2021. SCIMP is a spatiotemporal transmembrane scaffold for ERK1/2 to direct pro-inflammatory signaling in TLR-activated macrophages. Cell Reports, 36 (10), 109662.
- Ma, C., et al., 2021. Danggui sini decoction (herbal medicine) for the treatment of primary dysmenorrhoea: A systematic review and meta-analysis. Journal of Obstetrics and Gynaecology: The Journal of the Institute of Obstetrics and Gynaecology, 41 (7), 1001–1009.
- Marjoribanks, J., et al., 2015. Nonsteroidal anti-inflammatory drugs for dysmenorrhoea. The Cochrane Database of Systematic Reviews, 2015 (7), CD001751.
- Morita, W., et al., 2019. ERK1/2 drives IL-1beta-induced expression of TGF-beta1 and BMP-2 in torn tendons. Scientific Reports, 9 (1), 19005.
- Nakano, R., et al., 2020. Non-transcriptional and translational function of canonical NF-κB signaling in activating ERK1/2 in IL-1beta-induced COX-2 expression in synovial fibroblasts. Frontiers in Immunology, 11, 579266.
- Nakayama, M., et al., 2022. Porphyromonas gingivalis gingipains induce cyclooxygenase-2 expression and prostaglandin E2 production via ERK1/2-activated AP-1 (c-Jun/c-Fos) and IKK/NF-κp65 cascades. Journal of Immunology (Baltimore, MD. : 1950)), 208 (5), 1146–1154.
- Pluchino, N., et al., 2020. Estrogen receptor-alpha immunoreactivity predicts symptom severity and pain recurrence in deep endometriosis. Fertility and Sterility, 113 (6), 1224–1231.e1.
- Su, L., et al., 2022. Revealing the mechanism of raw and vinegar-processed Curcuma aromatica Salisb. [Zingiberaceae] regulates primary dysmenorrhea in rats via integrated metabolomics. Frontiers in Pharmacology, 13, 926291.
- Sun, Q., et al., 2017. ER-α36 mediates estrogen-stimulated MAPK/ERK activation and regulates migration, invasion, proliferation in cervical cancer cells. Biochemical and Biophysical Research Communications, 487 (3), 625–632.
- Tang, B., et al., 2020. NLRP3 inflammasome inhibitor MCC950 attenuates primary dysmenorrhea in mice via the NF-κB/COX-2/PG pathway. Journal of Inflammation (London, England), 17 (1), 22.
- Tong, H., et al., 2021. Bioactive constituents and the molecular mechanism of Curcumae rhizoma in the treatment of primary dysmenorrhea based on network pharmacology and molecular docking. Phytomedicine: International Journal of Phytotherapy and Phytopharmacology, 86, 153558.
- Tu, F. and Hellman, K., 2021. Primary dysmenorrhea: Diagnosis and therapy. Obstetrics and Gynecology, 137 (4), 752–752.
- Wong, J., et al., 2020. Salvia sclarea L. essential oil extract and its antioxidative phytochemical sclareol inhibit oxytocin-induced uterine hypercontraction dysmenorrhea model by inhibiting the Ca2+-MLCK-MLC20 signaling cascade: An ex vivo and in vivo study. Antioxidants, 9 (10), 991.
- Wright, E.B. and Lannigan, D.A., 2023. ERK1/2-RSK regulation of oestrogen homeostasis. The FEBS Journal, 290 (8), 1943–1953.
- Xie, Y., Qian, J. and Lu, Q., 2020. The therapeutic effect of Ge-Gen decoction on a rat model of primary dysmenorrhea: Label-free quantitative proteomics and bioinformatic analyses. BioMed Research International, 2020, 5840967–12.
- Yang, L., et al., 2016. Ge-Gen Decoction attenuates oxytocin-induced uterine contraction and writhing response: Potential application in primary dysmenorrhea therapy. Chinese Journal of Natural Medicines, 14 (2), 124–132.
- Yuan, L., et al., 2021. Shaofu Zhuyu decoction for primary dysmenorrhea: A protocol for systematic review and meta-analysis. Medicine, 100 (23), e26226.
- Zhai, B., et al., 2010. Leukotriene B(4) BLT receptor signaling regulates the level and stability of cyclooxygenase-2 (COX-2) mRNA through restricted activation of Ras/Raf/ERK/p42 AUF1 pathway. The Journal of Biological Chemistry, 285 (31), 23568–23580.
- Zhang, Y., et al., 2021. Metabolomics study of Guizhi Fuling capsules in rats with cold coagulation dysmenorrhea. Frontiers in Pharmacology, 12, 764904.
- Zhao, Q., et al., 2022. Pharmacokinetics-derived absorbed components responsible for Guizhi-Fuling capsule target PI3K/Akt-Erk to exert an anti-dysmenorrhea effect. Journal of Ethnopharmacology, 297, 115525.