
Abstract
A cGMP-dependent protein kinase (PKG) has previously been shown to regulate synaptic transmission at the Drosophila neuromuscular junction (NMJ) during acute oxidative stress, potentially through modulation of downstream K+ channel kinetics; however, the specific K+ channels through which PKG functions remains unclear. In this study, we hypothesized that PKG may be acting on calcium-activated large-conductance Slo K+ channels, or BK channels. We found that genetic elimination and pharmacological inhibition of BK channel conductance increases synaptic transmission tolerance to acute H2O2-induced oxidative stress. Furthermore, we discovered that activation of PKG in BK channel loss-of-function (Slo4) mutants significantly decreases time to stimulus-induced synaptic failure, providing the first evidence of PKG and BK channels functioning independently to control synaptic transmission tolerance to acute oxidative stress.
Introduction
Most organisms require oxygen for survival; however, the metabolism of oxygen produces cytotoxic by-products known as reactive oxygen species (ROS) (Liu, Fiskum, & Schubert, Citation2002). An overabundance of ROS can induce oxidative stress, which has been implicated in the pathophysiology of stroke (Cuzzocrea, Riley, Caputi, & Salvemini, Citation2001), traumatic brain injury (Juurlink & Paterson, Citation1998), and various neurodegenerative disorders (Federico et al., Citation2012). Because of its ability to withstand extreme fluctuations in oxygen levels (Haddad, Citation2006) and temperature (Feder, Citation1997; Misener, Chen, & Walker, Citation2001), the common fruit fly, Drosophila melanogaster, serves as an ideal model system for investigating mechanisms that control tolerance to acute stress. Previous studies have established that modulation of the cGMP-dependent protein kinase (PKG) pathway in Drosophila regulates tolerance to acute stress, including anoxia/hypoxia (Dawson-Scully et al., Citation2010), hyperthermia (Dawson-Scully, Armstrong, Kent, Robertson, & Sokolowski, Citation2007), and oxidative stress (Caplan, Milton, & Dawson-Scully, Citation2013).
In numerous studies, acute oxidative stress has been induced via application of hydrogen peroxide (H2O2), a ROS that increases intracellular calcium (Ca2+) to cytotoxic levels, a phenomenon called Ca2+ overload (Castro et al., Citation2004; Herson, Lee, Pinnock, Hughes, & Ashford, Citation1999; Josephson, Silverman, Lakatta, Stern, & Zweier, Citation1991; Zhang & Ma, Citation2008). It has been proposed that stress-induced Ca2+ overload increases potassium (K+) efflux, which disrupts ionic homeostasis via hyperpolarization of the neuron, triggering cellular damage, synaptic failure, and activation of apoptotic mechanisms (Caplan et al., Citation2013; Gerbino et al., Citation2009; Hughes, Bortner, Purdy, & Cidlowski, Citation1997; Money, Rodgers, McGregor, & Robertson, Citation2009). A previous publication from our lab found that inhibition of both PKG activity and K+ channel conductance increases synaptic transmission tolerance to H2O2-induced oxidative stress at the Drosophila neuromuscular junction (NMJ); further, the study demonstrated that 2.25 mM H2O2 exposure depolarizes resting membrane potential (RMP), potentially contributing to synaptic failure (Caplan et al., Citation2013). Findings from other studies have proposed a connection between PKG activity and K+ channel conductance (Dawson-Scully et al., Citation2010; Renger, Yao, Sokolowski, & Wu, Citation1999; White et al., Citation1993), suggesting that K+ channels may be downstream targets of the PKG signaling cascade; however, the specific K+ channels that facilitate the effects of the PKG pathway remain elusive.
Research in Drosophila has facilitated the discovery of various types of K+ channels, including Shab, Shaker, Shaw, and Slo channels (Frolov, Bagati, Casino, & Singh, Citation2012). In an attempt to isolate K+ channels involved downstream of PKG, we focused on Slo channels. Specifically, we targeted BK channels, or calcium-activated large-conductance Slo K+ channels, which have been shown to have a ten-fold increase in K+ conductance compared to classic voltage-gated K+ channels (Pallotta, Magleby, & Barrett, Citation1981; Stefani et al., Citation1997). We hypothesized that blockade of BK channels would increase synaptic transmission tolerance to H2O2-induced stress by reducing K+ efflux. Pharmacological activation of PKG using 8-bromo-cGMP has been shown to increase BK channel conductance (Alioua et al., Citation1998; White et al., Citation1993); consequently, we proposed that PKG modulates downstream BK channels in the regulation of synaptic transmission tolerance to acute oxidative stress.
In the present study, we compared synaptic transmission tolerance to 2.25 mM H2O2-induced acute oxidative stress at the Drosophila larval glutamatergic NMJ in w1118 control larvae and BK channel loss-of-function (Slo4) mutants. PKG activation in w1118 control and Slo4 larvae was induced using sildenafil citrate and 3′,5′-cyclic monophosphate 8-bromo sodium salt (8-bromo-cGMP). Activation of voltage-gated K+ channels in both strains was induced using dichloroacetate (DCA). Pharmacological reduction of BK channel activity in w1118 control larvae was induced with iberiotoxin. K+ channel inhibition was achieved in w1118 larvae using tetraethylammonium (TEA).
Materials and methods
Animals
Drosophila melanogaster third instar wandering larvae (≈110-h old) were utilized throughout this study. All larvae were reared on standard Bloomington stock fly food in an incubator at 25 °C under a 12:12-h light–dark cycle. Slo4 mutants on a w1118 genetic background were used as BK channel loss-of-function mutants (Atkinson, Robertson, & Ganetzky, Citation1991). Slo4 is a gamma ray-induced chromosomal inversion on the third chromosome between 96A17 and 96F5-8 (Atkinson et al., Citation1991; Kadas, Ryglewski, & Duch, Citation2015). The Slo4 mutants were gifted by Dr. Gregory Macleod, but were originally obtained from Dr. Nigel Atkinson. The w1118 control larvae were also gifted by Dr. Gregory Macleod, but were originally obtained from Dr. Nancy Bonnini.
Electrophysiology
All electrophysiological experiments were performed as previously described (Caplan et al., Citation2013). Briefly, individual w1118 or Slo4 mutant larvae were pinned to a glass dissecting dish containing Schneider’s insect medium (Sigma, St. Louis, MO), and a longitudinal incision was made along the dorsal side of the larva in order to remove the central nervous system and internal organs. An extracellular glass suction electrode was fashioned to stimulate segmental nerves synapsing with individual muscle segments, and a 60–90 MΩ intracellular recording electrode was utilized in place of a 40 mΩ electrode. Further, the insect medium was replaced with HL3 recording saline containing 1.5 mM Ca2+ and 20 mM Mg2+ (Caplan et al., Citation2013; Stewart, Schuster, Goodman, & Atwood, Citation1996).
To quantify synaptic transmission tolerance to various treatment conditions, excitatory junction potentials (EJPs) were evoked via repetitive stimulation (0.3-ms pulses delivered suprathreshold with a frequency of 1 Hz) and time to synaptic failure (EJP amplitude <1 mV) was recorded (Caplan et al., Citation2013); preparations that took a longer duration of time (minutes) to reach synaptic failure in the presence of 2.25 mM H2O2 possessed greater tolerance for acute oxidative stress compared to those that took a shorter duration of time. Measurements from larval preparations were initially recorded in HL3 saline; after 5 min, the HL3 saline bath was replaced with HL3 saline containing 2.25 mM H2O2 and a drug, depending on the treatment condition. After 10 min of H2O2 exposure, larval preparations were tested for acute effects on characteristic EJP parameters, including differences in waveform shape and amplitude. Intracellular recordings of resting membrane potential (RMP) were measured from larval muscle 6.
Pharmacology
Acute oxidative stress was elicited in larval preparations via the addition of 2.25 mM H2O2 to HL3 saline. Addition of pharmacological agents to the larval NMJ preparation was performed as previously described (Caplan et al., Citation2013); however, all drugs were water-soluble at the concentrations used and thus were dissolved in ddH2O as opposed to dimethyl sulfoxide (DMSO). A published dosage of 40 µM 8-bromo-cGMP (Caplan et al., Citation2013; Dawson-Scully et al., Citation2007) was administered to w1118 control and Slo4 mutant larvae to activate PKG. Sildenafil citrate, another PKG activator (Broderick, Kean, Dow, Pyne, & Davies, Citation2004; Day, Dow, Houslay, & Davies, Citation2005; Francis, Turko, & Corbin, Citation2001), was also administered to w1118 and Slo4 mutant larvae at the 40 µM dosage. Iberiotoxin, a BK channel inhibitor (McKay et al., Citation1994), was administered at the 500 pM dosage in w1118 and Slo4 mutant larvae. Additionally, 0.25 mM DCA, a voltage-gated K+ channel activator (Bonnet et al., Citation2007; Michelakis et al., Citation2002), was administered to w1118 and Slo4 mutant larvae. A published dosage of 0.25 mM TEA, a K+ channel inhibitor (Caplan et al., Citation2013), was administered to w1118 larvae. Dose–response curves were utilized to determine the optimal dosage of all drugs presented in this manuscript. All drugs and chemicals used in this study were obtained from Sigma-Aldrich Inc. (Sigma-Aldrich Inc., St. Louis, MO).
Statistics
All data were analyzed using SigmaPlot (Systat Software Inc., San Jose, CA). One-way analysis of variance (ANOVA) and Holm–Sidak post hoc analysis were used to compare significant differences across groups. Statistical significance (p < .05) in the vertical bar charts was designated using letters, where different letters show statistically significant differences and the same letter indicates non-significance. The letter ‘A’ designates the highest mean, ‘B’ indicates the next highest mean, and so forth. All vertical bar charts are presented as mean ± SEM.
Results
Genetic elimination of BK channels regulates synaptic transmission tolerance to acute oxidative stress
To investigate whether inhibition of BK channel conductance would increase synaptic transmission tolerance to 2.25 mM H2O2-induced oxidative stress, average time to synaptic failure of BK channel loss-of-function (Slo4) mutants and w1118 control larvae was compared; w1118 control larvae underwent synaptic failure significantly earlier than Slo4 mutants under conditions of acute oxidative stress [one-way ANOVA, F(3, 16) = 17.76, p < .05; ]. w1118 and Slo4 mutant larvae displayed varying rates of EJP amplitude decline, as shown in . Further, application of 2.25 mM H2O2 in both the w1118 control and Slo4 mutant larvae did not acutely affect the characteristic shape of the EJP waveforms (). Interestingly, time to synaptic failure did not significantly differ in Slo4 mutants in either the presence or absence of H2O2 [p = .917; ]. Elimination of BK channels in Slo4 mutants seems to have conferred resistance to H2O2 due to the large reduction of K+ channel conductance.
Figure 1. Genetic elimination of BK channels increases synaptic transmission tolerance to acute oxidative stress. (A) Average time to synaptic failure is prolonged in Slo4 mutants compared to w1118 control larvae during H2O2 exposure [one-way ANOVA, F(3, 16) = 17.76, p < .05]. Vertical bar chart is shown as mean ± SEM. Different letters on bar charts indicate statistical significance, whereas the same letter indicates a nonsignificant difference. (B) Average amplitude decline of evoked EJPs in w1118 controls and Slo4 mutants in the presence and absence of 2.25 mM H2O2 (n = 4–6 preparations per group). Black arrows indicate time to synaptic failure of individual w1118 control larval preparations in HL3 saline. Each data point is displayed as mean ± SEM. (C) Representative waveforms of evoked EJPs from w1118 control larvae 1 min before H2O2 exposure (black line) and 10 min after H2O2 exposure (gray line). (D) Representative waveforms of evoked EJPs from Slo4 mutant larvae 1 min before H2O2 exposure (black line) and 10 min after H2O2 exposure (gray line). H2O2 does not acutely affect characteristic EJP shape in both w1118 control and Slo4 mutant larvae.
![Figure 1. Genetic elimination of BK channels increases synaptic transmission tolerance to acute oxidative stress. (A) Average time to synaptic failure is prolonged in Slo4 mutants compared to w1118 control larvae during H2O2 exposure [one-way ANOVA, F(3, 16) = 17.76, p < .05]. Vertical bar chart is shown as mean ± SEM. Different letters on bar charts indicate statistical significance, whereas the same letter indicates a nonsignificant difference. (B) Average amplitude decline of evoked EJPs in w1118 controls and Slo4 mutants in the presence and absence of 2.25 mM H2O2 (n = 4–6 preparations per group). Black arrows indicate time to synaptic failure of individual w1118 control larval preparations in HL3 saline. Each data point is displayed as mean ± SEM. (C) Representative waveforms of evoked EJPs from w1118 control larvae 1 min before H2O2 exposure (black line) and 10 min after H2O2 exposure (gray line). (D) Representative waveforms of evoked EJPs from Slo4 mutant larvae 1 min before H2O2 exposure (black line) and 10 min after H2O2 exposure (gray line). H2O2 does not acutely affect characteristic EJP shape in both w1118 control and Slo4 mutant larvae.](/cms/asset/146c5645-c6ca-4b6f-aec4-d4848776db9a/ineg_a_1500571_f0001_b.jpg)
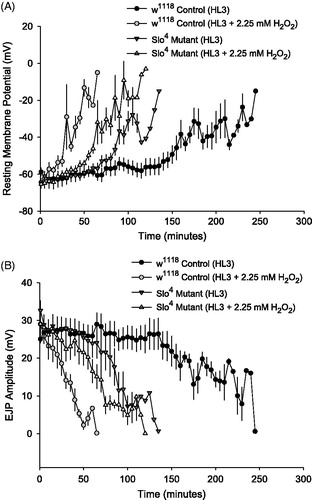
Further, application of 2.25 mM H2O2 to both w1118 and Slo4 mutant larvae affected resting membrane potential (RMP) and, consequently, EJP amplitude (. RMP depolarization occurred more rapidly in w1118 and Slo4 larvae exposed to 2.25 mM H2O2 compared to HL3 controls (); a corresponding decrease in EJP amplitude was observed in the same larval preparations (). Hyperpolarized RMPs corresponded to higher EJP amplitudes, whereas depolarized RMPs corresponded to lower EJP amplitudes.
Figure 2. Drosophila muscle resting membrane potential (RMP) and EJP amplitude decline during acute oxidative stress. (A) RMP from Drosophila larval muscle 6 was recorded in w1118 control larvae and Slo4 mutants in the presence and absence of 2.25 mM H2O2 (n = 6 per group). RMP depolarization occurs more rapidly in w1118 and Slo4 mutant larvae exposed to 2.25 mM H2O2 compared to HL3 control larvae. (B) In the same larval preparations, EJP amplitude was also recorded (n = 6 per group). Loss of RMP tracks EJP amplitude decline across all treatments.
PKG modulation of downstream K+ channels mediates synaptic transmission tolerance to acute stress
Genetic elimination of BK channel conductance and pharmacological activation of the PKG pathway were combined to determine the potential mechanistic action of PKG on downstream BK channels. Slo4 mutants were exposed to oxidative stress (2.25 mM H2O2) and treated with 40 µM of PKG activators 8-bromo-cGMP and sildenafil citrate. Application of 40 µM 8-bromo-cGMP significantly reduced time to synaptic failure in Slo4 mutants compared to mutants only treated with H2O2 [one-way ANOVA, F(9, 53) = 13.24, p < .001; ], as did application of 40 µM sildenafil citrate [p < .001; ]. Contrary to our original hypothesis, these results suggest that PKG modulates K+ channels other than BK channels to regulate neuromuscular communication during H2O2-induced stress. Activation of PKG using 8-bromo-cGMP in w1118 control larvae exposed to acute oxidative stress did not significantly alter synaptic transmission tolerance compared to w1118 larvae only treated with H2O2 [p = .651; ]. Additionally, activation of PKG using sildenafil citrate did not significantly alter time to synaptic failure during acute oxidative stress in w1118 larvae [p = 0.313; ]. These results suggest that activation of PKG does not exacerbate the deleterious effects of H2O2 in w1118 control larvae.
Figure 3. PKG functions independently of BK channels to protect synaptic function from H2O2 exposure. PKG activation using 40 µM 8-bromo-cGMP [one-way ANOVA, F(9, 53) = 13.24, p < .001] and 40 µM sildenafil citrate [p < .001] in Slo4 mutant larvae significantly decreases time to synaptic failure during acute oxidative stress. Activation of PKG with 40 µM 8-bromo-cGMP [p = .651] and 40 µM sildenafil citrate [p = .313] does not alter time to synaptic failure in w1118 control larvae. Activation of voltage-gated K+ channels using 0.25 mM DCA in w1118 control larvae does not alter time to synaptic failure during acute oxidative stress [p = .561]. Application of 0.25 mM DCA to Slo4 mutant larval preparations decreases time to synaptic failure during oxidative stress [p < .05]. Vertical bar chart is shown as mean ± SEM. n = 5–9 preparations per group.
![Figure 3. PKG functions independently of BK channels to protect synaptic function from H2O2 exposure. PKG activation using 40 µM 8-bromo-cGMP [one-way ANOVA, F(9, 53) = 13.24, p < .001] and 40 µM sildenafil citrate [p < .001] in Slo4 mutant larvae significantly decreases time to synaptic failure during acute oxidative stress. Activation of PKG with 40 µM 8-bromo-cGMP [p = .651] and 40 µM sildenafil citrate [p = .313] does not alter time to synaptic failure in w1118 control larvae. Activation of voltage-gated K+ channels using 0.25 mM DCA in w1118 control larvae does not alter time to synaptic failure during acute oxidative stress [p = .561]. Application of 0.25 mM DCA to Slo4 mutant larval preparations decreases time to synaptic failure during oxidative stress [p < .05]. Vertical bar chart is shown as mean ± SEM. n = 5–9 preparations per group.](/cms/asset/21c6ad0b-9611-4e2c-8e7f-6496a5541f5e/ineg_a_1500571_f0003_b.jpg)
To further confirm that PKG acts on K+ channels other than BK channels, we manipulated K+ channel kinetics using DCA, a voltage-gated K+ channel activator. Administration of 0.25 mM DCA and 2.25 mM H2O2 to Slo4 mutant larvae led to a significant reduction in time to synaptic failure compared to Slo4 H2O2 control larvae [p < .05; ], suggesting that non-BK K+ channels also modulate synaptic transmission during H2O2 stress. In w1118 larvae, 0.25 mM DCA did not significantly alter synaptic transmission tolerance to acute stress [p = .561; ]. Taken together, these results suggest that the PKG pathway functions independently of BK channels in the regulation of synaptic transmission tolerance to acute oxidative stress, potentially through the modulation of other downstream voltage-gated K+ channels.
Pharmacological inhibition of K+ channel conductance regulates neurotransmission during acute oxidative stress
To further examine the role of BK channels in protecting synaptic transmission during acute oxidative stress, low doses of iberiotoxin were administered to w1118 control larval preparations. Application of 500 pM iberiotoxin increased synaptic transmission tolerance in w1118 larvae exposed to 2.25 mM H2O2 compared to w1118 H2O2 control larvae [one-way ANOVA, F(3, 19) = 9.449, p < .05; ]. Thus, acute pharmacological inhibition of BK channel conductance in w1118 larvae has similar protective effects compared to genetic elimination of BK channel conductance.
Figure 4. Pharmacological blockade of BK channel conductance protects synaptic transmission from acute stress. Pharmacological inhibition of BK channel conductance using 500 pM iberiotoxin significantly increases time to synaptic failure in w1118 control larvae exposed to 2.25 mM H2O2 compared to w1118 H2O2 controls [one-way ANOVA, F(3, 19) = 9.449, p < .05]. Nonselective K+ channel inhibition using 0.25 mM TEA also increases synaptic transmission tolerance to acute oxidative stress in w1118 control larvae [p < .05]. Vertical bar chart is shown as mean ± SEM. n = 5–6 preparations per group.
![Figure 4. Pharmacological blockade of BK channel conductance protects synaptic transmission from acute stress. Pharmacological inhibition of BK channel conductance using 500 pM iberiotoxin significantly increases time to synaptic failure in w1118 control larvae exposed to 2.25 mM H2O2 compared to w1118 H2O2 controls [one-way ANOVA, F(3, 19) = 9.449, p < .05]. Nonselective K+ channel inhibition using 0.25 mM TEA also increases synaptic transmission tolerance to acute oxidative stress in w1118 control larvae [p < .05]. Vertical bar chart is shown as mean ± SEM. n = 5–6 preparations per group.](/cms/asset/61c8b311-5307-4cb6-9700-45022edf3083/ineg_a_1500571_f0004_b.jpg)
To explore the effects of nonselective K+ channel inhibition on synaptic transmission, we applied TEA to w1118 larvae exposed to 2.25 mM H2O2. We found that administration of 0.25 mM TEA increased synaptic transmission tolerance during acute oxidative stress, similar to the effects of 500 pM iberiotoxin in w1118 larval preparations [p < .05; ]. These results suggest that pharmacological inhibition of both BK channels and classic voltage-gated K+ channels protects synaptic transmission from the harmful effects of acute oxidative stress.
Iberiotoxin does not possess major off-target effects in Slo4 mutants
Although iberiotoxin has been shown to inhibit Drosophila BK channels in GH3 cells (McKay et al., Citation1994), its effects have not been observed at the Drosophila NMJ; to address this concern, we tested for potential off-target effects by administering 500 pM iberiotoxin to Slo4 mutants. Because Slo4 mutants lack BK channels, a low dosage of iberiotoxin should not significantly increase synaptic transmission tolerance to acute oxidative stress; consistent with this hypothesis, 500 pM iberiotoxin did not significantly alter time to synaptic failure in Slo4 mutants during H2O2 exposure [one-way ANOVA, F(2, 16), p = .736; ], indicating that iberiotoxin specifically modulates BK channels at this dosage and does not possess any major off-target effects using our synaptic transmission assay.
Figure 5. Iberiotoxin does not possess major off-target effects using this experimental model. Administration of BK channel inhibitor iberiotoxin does not significantly alter time to synaptic failure in Slo4 mutant larvae [one-way ANOVA, F(2, 16), p = .736], suggesting that iberiotoxin does not possess any major off-target effects at the Drosophila larval NMJ. Vertical bar chart is shown as mean ± SEM. n = 4–6 preparations per group.
![Figure 5. Iberiotoxin does not possess major off-target effects using this experimental model. Administration of BK channel inhibitor iberiotoxin does not significantly alter time to synaptic failure in Slo4 mutant larvae [one-way ANOVA, F(2, 16), p = .736], suggesting that iberiotoxin does not possess any major off-target effects at the Drosophila larval NMJ. Vertical bar chart is shown as mean ± SEM. n = 4–6 preparations per group.](/cms/asset/bc090c7b-2187-42be-ab85-0672c815a93d/ineg_a_1500571_f0005_b.jpg)
Discussion
Previous work demonstrated that PKG regulates synaptic transmission tolerance to acute oxidative stress at the Drosophila NMJ, possibly through modulation of K+ channel conductance (Caplan et al., Citation2013). The present study served to further investigate which K+ channels might be functioning downstream of the PKG pathway. Findings from the present study provide evidence of PKG and BK channel regulation of synaptic function during acute oxidative stress. Genetic and pharmacological inhibition of BK channels led to increases in synaptic transmission tolerance to 2.25 mM H2O2-induced oxidative stress; in addition, nonselective inhibition of K+ channels in w1118 larvae also increases time to synaptic failure during acute oxidative stress. The results indicate that PKG and BK channels function independently to regulate synaptic transmission during acute stress; in other words, PKG acts on downstream K+ channels other than BK channels to control neuromuscular communication during H2O2 exposure.
Using H2O2-induced stress as a model to probe effects of PKG pathway manipulation on neurotransmission in Drosophila
H2O2 has been used to model oxidative stress in various neurological disorders, including Alzheimer’s disease (Behl, Davis, Lesley, & Schubert, Citation1994; Miyata & Smith, Citation1996), Parkinson’s disease (Jenner, Citation2003), and amyotrophic lateral sclerosis (Nani, Cifra, & Nistri, Citation2010). H2O2 has also been shown to modulate neurotransmission in multiple electrophysiological paradigms, including the frog NMJ (Giniatullin & Giniatullin, Citation2003), rat striatal neurons (Smith, Herson, Lee, Pinnock, & Ashford, Citation2003), and the Drosophila NMJ (Caplan et al., Citation2013). Thus, studying H2O2 exposure at the Drosophila NMJ serves as an ideal model to study novel targets of the PKG signaling cascade, enabling future work in the development of protective pathways that can counteract the deleterious effects of oxidative stress.
Role of BK channels in ionic homeostasis and synaptic transmission
A stimulus-induced rise in [Ca2+]i has been shown to activate BK channels in Drosophila and mammalian neurons (Bian, Favre, & Moczydlowski, Citation2001; DiChiara & Reinhart, Citation1995; Nimigean & Magleby, Citation1999; Piskorowski & Aldrich, Citation2002), resulting in BK channel-dependent efflux of [K+]i, repolarization of the membrane, and reduction of Ca2+ influx (Faber & Sah, Citation2003; Lancaster & Nicoll, Citation1987; Vetri, Choudhury, Pelligrino, & Sundivakkam, Citation2014). BK channels also modulate various neurophysiological processes, including neurotransmitter release, interspike interval, and spike frequency (Faber & Sah, Citation2003). Consequently, BK channels have been shown to regulate many essential physiological processes in both fruit flies and mammals. For instance, BK channels have been implicated in several human ailments, such as urinary incontinence (Lawson, Citation2000), neuromuscular abnormalities (Lehmann & Jurkat, Citation1999), hypertension (Yang et al., Citation2013), and epilepsy (Mantegazza, Curia, Biagini, Ragsdale, & Avoli, Citation2010). In the present study, BK channels were found to mediate synaptic transmission tolerance to acute oxidative stress at the Drosophila larval NMJ.
Potential downstream targets of the PKG pathway
Our results demonstrate that activation of PKG in Slo4 mutants significantly reduces synaptic transmission tolerance to acute oxidative stress. TEA was used as a nonselective K+ channel inhibitor in the present study. Interestingly, it has been shown that low doses of TEA inhibit Drosophila slo channels expressed in Xenopus oocytes (Shen et al., Citation1994) as well as voltage-gated K+ channels, including K3 channels, in Drosophila muscle (Gorczyca & Wu, Citation1991). Our results demonstrate that application of 0.25 mM TEA to w1118 controls protects synaptic transmission from oxidative stress, likely due to inhibition of both BK channels and classic voltage-gated K+ channels. Moreover, we propose that PKG functions independently of BK channel conductance in the modulation of synaptic transmission during acute stress.
Notably, application of DCA to Slo4 mutants significantly decreased time to synaptic failure during acute oxidative stress. A previous publication similarly utilized DCA to activate voltage-gated K+ channels in Drosophila (Dawson-Scully et al., Citation2010); however, this study did not directly demonstrate DCA activation of voltage-gated K+ channels in neurons or muscles. Recently, it has been shown that mature flies fed DCA display acute increases in ROS production and oxidative stress (Pandey et al., Citation2014). Therefore, the DCA-induced decrease in time to synaptic failure in Slo4 mutants could potentially be attributed to DCA activating K+ channels, exacerbating oxidative stress, or modulating other metabolic processes.
Action potential dynamics as potential mechanism for protective effect of K+ channel conductance inhibition
In the current study, genetic inhibition of BK channel conductance using Slo4 mutants and pharmacological inhibition using iberiotoxin led to increased synaptic transmission tolerance to acute oxidative stress. Inhibition of K+ channel conductance using TEA resulted in similar extensions in time to synaptic failure. Inhibiting K+ channel conductance results in reduced K+ efflux, which has been shown to prevent the initiation of cellular apoptotic cascades (Bortner, Hughes, & Cidlowski, Citation1997). Additionally, blocking K+ channel conductance depolarizes neurons, broadening the action potential to release neurotransmitters; this increased transmitter release may delay rundown of synaptic transmission, or increase time to synaptic failure during acute oxidative stress. Previously published work provides support for this potential mechanism by demonstrating that pharmacological inhibition of Shaker K+ channels in Slo4 mutants increases neurotransmitter release and excitatory postsynaptic potential amplitude compared to wild-type controls at the Drosophila NMJ (Lee, Ueda, & Wu, Citation2008; Lee, Ueda, & Wu, Citation2014). Consistent with this finding, pharmacological inhibition of BK channel conductance leads to an increase in transmitter release within the mammalian hippocampus (Griguoli, Sgritta, & Cherubini, Citation2016; Raffaelli, Saviane, Mohajerani, Pedarzani, & Cherubini, Citation2004).
Additionally, the relationship between synaptic failure and resting membrane potential has been previously studied. A group found that RMP in Drosophila larval body wall muscles is directly correlated with EJP amplitude in electrophysiological recordings utilizing HL3 saline; consistent with our findings, they demonstrated that hyperpolarized RMPs were correlated with higher EJP amplitudes, whereas depolarized RMPs were correlated with lower EJP amplitudes (Krans, Parfitt, Gawera, Rivlin, & Hoy, Citation2010). Thus, synaptic transmission failure could be partly attributed to changes in RMP.
Using BK channel inhibitors to confer neuroprotection
Recent studies have indicated that activation of BK channel conductance protects neurons from stroke-induced cell death (Chi et al., Citation2010; Gribkoff et al., Citation2001; Li et al., Citation2014). However, the drugs utilized in these studies (chlorpromazine, BMS-204352, and Baifuzi) possess major off-target effects; for instance, CPZ inhibits dopamine D2 receptors (Carpenter & Koenig, Citation2008), reduces serotonergic activity (Croll, Baker, Khabarova, Voronezhskaya, & Sakharov, Citation1997), and modulates Ca2+ channels (McNaughton, Green, & Randall, Citation2001), Cl− channels (Quamme, Citation1997), and nicotinic receptor channels (Benoit & Changeux, Citation1993). Thus, protection of neurons from stroke may not be specifically attributable to BK channel activation.
A previous study assessed the protective effect of BK channel blockade in a human neuronal cell line SH-SY5Y during H2O2-induced oxidative stress and CoCl2-induced hypoxia mimetic insults; the results revealed that specific BK channel activators, isopimaric acid and NS1619, exacerbated cytotoxicity, whereas inhibition of BK channels using TEA and tetrandrine provided a neuroprotective effect (Zhang, Citation2016). Because increases in [Ca2+]i activate BK channels, K+ channel blockers may be utilized in protecting neurons from oxidative stress. The findings from Zhang (Citation2016) are consistent with our results, demonstrating that inhibition of BK channel and voltage-gated K+ channel conductance protects synaptic transmission during acute oxidative stress. Further confirming the role of BK channel inhibition in neuroprotection, a recent study provided evidence of BK channel inhibitors, including iberiotoxin, charybdotoxin, paxilline, and TEA, protecting neurons from stroke-induced insults (Chen et al., Citation2013). Consequently, the results from Chen et al. (Citation2013) indicate that optimizing the dosage range of specific BK channel blockers (iberiotoxin) and K+ channel inhibitors (TEA) may assist in the development of effective treatments for stroke-induced cell death.
Iberiotoxin has been used to inhibit BK channel conductance in mammalian systems (McKay et al., Citation1994). BK channels expressed in Xenopus oocytes have been shown to be insensitive to iberiotoxin up to 100 nM (Meera, Wallner, Song, & Toro, Citation1997). Thus, there is no direct evidence of low doses of iberiotoxin inhibiting Drosophila BK channels; however, 200 nM charybdotoxin has been shown to inhibit the BK channel encoded K+ current (ICF current) at the Drosophila NMJ (Kadas et al., Citation2015). Despite its highly similar amino acid sequence and mechanism of action, iberiotoxin has been shown to be approximately ten times more effective at inhibiting rat BK channels compared to charybdotoxin (Candia, Garcia, & Latorre, Citation1992). If low doses of iberiotoxin also inhibited non-BK K+ channels, we might expect to observe an increase in time to synaptic failure in Slo4 mutants using our assay; contrarily, our results demonstrate that application of 0.5 nM iberiotoxin to the Drosophila NMJ does not significantly modify time to synaptic failure in Slo4 mutants. Thus, 0.5 nM iberiotoxin may specifically act on BK channels as opposed to other K+ channels at the Drosophila NMJ. Iberiotoxin was used in this study because of its structural similarity to charybdotoxin; unlike charybdotoxin, however, iberiotoxin has not demonstrated any off-target effects at the larval NMJ (Candia et al., Citation1992; Kadas et al., Citation2015).
Localization of BK channel-specific neuroprotection at the Drosophila NMJ
Although this study provided evidence of increased synaptic transmission tolerance to H2O2 exposure through BK channel inhibition, we did not localize the effect to presynaptic motor neurons or postsynaptic muscles. Expression of BK channels in presynaptic boutons at the Drosophila larval NMJ has been shown (Zhou et al., Citation1999); however, expression patterns of BK channels in postsynaptic muscles are not well-characterized via antibody expression. Despite this, various morphological and electrophysiological studies strongly suggest that BK channels are expressed in both larval (Komatsu, Singh, Rathe, & Wu, Citation1990; Lee et al., Citation2008; Singh & Wu, Citation1990) and adult muscles (Elkins & Ganetzky, Citation1988). Future work will elucidate the mechanisms through which muscle-specific BK channels function to protect synaptic transmission from acute oxidative stress.
Quantifying synaptic transmission in the presence of physiological stress at the larval NMJ is an effective way to identify protective genes and pharmacological agents
Many groups have utilized the Drosophila larval NMJ to identify drugs or genes that mediate cell death, or synaptic failure. For instance, results from a previous study indicate that a prior heat-shock protects synaptic transmission during subsequent heat-induced stress and that this effect is largely mediated by genes encoding Hsp70 (Karunanithi, Barclay, Robertson, Brown, & Atwood, Citation1999). Additionally, pharmacological inhibition of the PKG pathway was found to increase synaptic transmission tolerance to acute heat stress at the larval NMJ (Dawson-Scully et al., Citation2007; Krill & Dawson-Scully, Citation2016). Recently, it was discovered that pseudopterosin A (PsA), a natural product, increases synaptic transmission tolerance to acute oxidative stress at the Drosophila NMJ (Caplan, Zheng, Dawson-Scully, White, & West, Citation2016). Thus, the Drosophila larval NMJ is a well-established model for studying neurotransmission during physiological stress and serves as a starting point for identifying neuromodulatory drugs that protect synaptic function from acute stress.
Conclusions
In this study, we provide evidence of increased synaptic transmission tolerance to acute oxidative stress through genetic elimination and acute pharmacological inhibition of BK channel conductance. Additionally, we demonstrated that BK channels function independently of PKG-modulated K+ channels to protect synaptic transmission during H2O2 exposure. Consistent with these results, we found that acute activation of voltage-gated K+ channel conductance reduced time to synaptic failure during oxidative stress in Slo4 mutants. These results suggest that reduction of the overall conductance of BK channels, non-BK voltage-gated K+ channels, and PKG-modulated K+ channels prolongs synaptic function during acute stress at the Drosophila NMJ. Thus, development of specific K+ channel modulators may emerge as novel therapies for the treatment of oxidative stress-induced physiological insults.
Acknowledgements
We would like to thank Dr. Gregory Macleod for providing fly stocks and critical feedback. We also thank Dr. William Ja, Dr. Colin Hughes, Stephanie Kelly, Karlis Justs, and anonymous reviewers for providing valuable insights.
This article is dedicated to Professor Harold Atwood, who spent a career building a body of knowledge on the integration of morphological and physiological processes that modulate neuromuscular junction synaptic transmission. He also created a vast field of study based on the numerous investigators he trained through the decades of his works. On a personal note, I strive to create a scientific and social environment like the one I experienced as a Ph.D. student. As most young people, I mistakenly assumed that having such an environment was an easy thing to implement; but I see now that skill and expertise is also required to create an environment that built, what most of us remember as, the Atwood Lab. I will continue to strive for this with my own laboratory.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Alioua, A., Tanaka, Y., Wallner, M., Hofmann, F., Ruth, P., Meera, P., & Toro, L. (1998). The large conductance, voltage-dependent, and calcium-sensitive K + channel, Hslo, is a target of cGMP-dependent protein kinase phosphorylation in vivo. Journal of Biological Chemistry, 273, 32950–32956. doi:10.1074/jbc.273.49.32950
- Atkinson, N., Robertson, G., & Ganetzky, B. (1991). A component of calcium-activated potassium channels encoded by the Drosophila slo locus. Science, 253, 551–555. doi:10.1126/science.1857984
- Behl, C., Davis, J.B., Lesley, R., & Schubert, D. (1994). Hydrogen peroxide mediates amyloid β protein toxicity. Cell, 77, 817–827. doi:10.1016/0092-8674(94)90131-7
- Benoit, P., & Changeux, J. P. (1993). Voltage dependencies of the effects of chlorpromazine on the nicotinic receptor channel from mouse muscle cell line So18. Neuroscience Letters, 160, 81–84. doi:10.1016/0304-3940(93)90918-B
- Bian, S., Favre, I., & Moczydlowski, E. (2001). Ca2+-binding activity of a COOH-terminal fragment of the Drosophila BK channel involved in Ca2+-dependent activation. Proceedings of the National Academy of Sciences, 98, 4776–4781. doi:10.1073/pnas.081072398
- Bonnet, S., Archer, S. L., Allalunis-Turner, J., Haromy, A., Beaulieu, C., Thompson, R., … Michelakis, E. D. (2007). A mitochondria-K + channel axis is suppressed in cancer and its normalization promotes apoptosis and inhibits cancer growth. Cancer Cell, 11, 37–51. doi:10.1016/j.ccr.2006.10.020
- Bortner, C. D., Hughes, F. M., & Cidlowski, J. A. (1997). A primary role for K + and Na + efflux in the activation of apoptosis. Journal of Biological Chemistry, 272, 32436–32442. doi:10.1074/jbc.272.51.32436
- Broderick, K. E., Kean, L., Dow, J. A. T., Pyne, N. J., & Davies, S. A. (2004). Ectopic expression of bovine type 5 phosphodiesterase confers a renal phenotype in Drosophila. Journal of Biological Chemistry, 279, 8159–8168. doi:10.1074/jbc.M304679200
- Candia, S., Garcia, M. L., & Latorre, R. (1992). Mode of action of iberiotoxin, a potent blocker of the large conductance Ca(2+)-activated K + channel. Biophysical Journal, 63, 583–590. doi:10.1016/S0006-3495(92)81630-2
- Caplan, S. L., Milton, S. L., & Dawson-Scully, K. (2013). A cGMP-dependent protein kinase (PKG) controls synaptic transmission tolerance to acute oxidative stress at the Drosophila larval neuromuscular junction. Journal of Neurophysiology, 109, 649–658. doi:10.1152/jn.00784.2011
- Caplan, S. L., Zheng, B., Dawson-Scully, K., White, C. A., & West, L. M. (2016). Pseudopterosin a: Protection of synaptic function and potential as a neuromodulatory agent. Marine Drugs, 14, 55. doi:10.3390/md14030055
- Carpenter, W. T., & Koenig, J. I. (2008). The evolution of drug development in schizophrenia: Past issues and future opportunities. Neuropsychopharmacology: Official Publication of the American College of Neuropsychopharmacology, 33, 2061–2079. doi:10.1038/sj.npp.1301639
- Castro, J., Bittner, C. X., Humeres, A., Montecinos, V. P., Vera, J. C., & Barros, L. F. (2004). A cytosolic source of calcium unveiled by hydrogen peroxide with relevance for epithelial cell death. Cell Death and Differentiation, 11, 468–478. doi:10.1038/sj.cdd.4401372
- Chen, M., Sun, H. Y., Hu, P., Wang, C. F., Li, B. X., Li, S. J., … Gao, T. M. (2013). Activation of BKCa channels mediates hippocampal neuronal death after reoxygenation and reperfusion. Molecular Neurobiology, 48, 794–807. https://doi.org/10.1007/s12035-013-8467-x
- Chi, S., Cai, W., Liu, P., Zhang, Z., Chen, X., Gao, L., … Qi, Z. (2010). Baifuzi reduces transient ischemic brain damage through an interaction with the STREX domain of BKCa channels. Cell Death Dis, 1, e13. doi:10.1038/cddis.2009.10
- Croll, R. P., Baker, M. W., Khabarova, M., Voronezhskaya, E. E., & Sakharov, D. A. (1997). Serotonin depletion after prolonged chlorpromazine treatment in a simpler model system. General Pharmacology, 29, 91–96. doi:10.1016/S0306-3623(96)00530-7
- Cuzzocrea, S., Riley, D. P., Caputi, A. P., & Salvemini, D. (2001). Antioxidant therapy: A new pharmacological approach in shock, inflammation, and ischemia/reperfusion injury. Pharmacological Reviews, 426, 1–159. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/11171943
- Dawson-Scully, K., Armstrong, G. A. B., Kent, C., Robertson, R. M., & Sokolowski, M. B. (2007). Natural variation in the thermotolerance of neural function and behavior due to a cGMP-dependent protein kinase. PLoS Ond, 2, e773. doi:10.1371/journal.pone.0000773
- Dawson-Scully, K., Bukvic, D., Chakaborty-Chatterjee, M., Ferreira, R., Milton, S. L., & Sokolowski, M. B. (2010). Controlling anoxic tolerance in adult Drosophila via the cGMP-PKG pathway. Journal of Experimental Biology, 213, 2410–2416. doi:10.1242/jeb.041319
- Day, J. P., Dow, J. A. T., Houslay, M. D., & Davies, S. (2005). Cyclic nucleotide phosphodiesterases in Drosophila melanogaster. The Biochemical Journal, 388, 333–342. doi:10.1042/BJ20050057
- DiChiara, T. J., & Reinhart, P. H. (1995). Distinct effects of Ca2+ and voltage on the activation and deactivation of cloned Ca(2+)-activated K + channels. The Journal of Physiology, 489, 403–418. doi:10.1113/jphysiol.1995.sp021061
- Elkins, T., & Ganetzky, B. (1988). The roles of potassium currents in Drosophila flight muscles. Journal of Neuroscience, 8, 428–434. doi:10.1523/JNEUROSCI.08-02-00428.1988
- Feder, M. E. (1997). Necrotic fruit: A novel model system for thermal ecologists. Journal of Thermal Biology, 22, 1–9. doi:10.1016/S0306-4565(96)00028-9
- Faber, E. S. L., & Sah, P. (2003). Calcium-activated potassium channels: Multiple contributions to neuronal function. The Neuroscientist: A Review Journal Bringing Neurobiology, Neurology and Psychiatry, 9, 181–194. https://doi.org/10.1177/1073858403252673
- Federico, A., Cardaioli, E., Da Pozzo, P., Formichi, P., Gallus, G. N., & Radi, E. (2012). Mitochondria, oxidative stress and neurodegeneration. Journal of the Neurological Sciences, 322, 254–262. doi:10.1016/j.jns.2012.05.030
- Francis, S. H., Turko, I. V., & Corbin, J. D. (2001). Cyclic nucleotide phosphodiesterases: Relating structure and function. Progress in Nucleic Acid Research and Molecular Biology, 65, 1–52. https://doi.org/10.1016/S0079-6603(00)65001-8
- Frolov, R. V., Bagati, A., Casino, B., & Singh, S. (2012). Potassium channels in drosophila: Historical breakthroughs, significance, and perspectives. Journal of Neurogenetics, 26, 275–290. https://doi.org/10.3109/01677063.2012.744990
- Gerbino, A., Ranieri, M., Lupo, S., Caroppo, R., Debellis, L., Maiellaro, I., … Colella, M. (2009). Ca2+-dependent K + efflux regulates deoxycholate-induced apoptosis of BHK-21 and Caco-2 cells. Gastroenterology, 137, 955. doi:10.1053/j.gastro.2009.03.038
- Giniatullin, A. R., & Giniatullin, R. A. (2003). Dual action of hydrogen peroxide on synaptic transmission at the frog neuromuscular junction. The Journal of Physiology, 552, 283–293. doi:10.1113/jphysiol.2003.050690
- Gorczyca, M. G., & Wu, C. F. (1991). Single-channel K + currents in Drosophila muscle and their pharmacological block. The Journal of Membrane Biology, 121, 237–248. https://doi.org/10.1007/BF01951557
- Gribkoff, V. K., Starrett, J. E., Dworetzky, S. I., Hewawasam, P., Boissard, C. G., Cook, D. A., … Yeola, S. W. (2001). Targeting acute ischemic stroke with a calcium-sensitive opener of maxi-K potassium channels. Nature Medicine, 7, 471–477. doi:10.1038/86546
- Griguoli, M., Sgritta, M., & Cherubini, E. (2016). Presynaptic BK channels control transmitter release: Physiological relevance and potential therapeutic implications. The Journal of Physiology, 594, 3489–3500. doi:10.1113/JP271841
- Haddad, G. G. (2006). Tolerance to low O2: Lessons from invertebrate genetic models. Experimental Physiology, 91, 277–282. doi:10.1113/expphysiol.2005.030767
- Herson, P. S., Lee, K., Pinnock, R. D., Hughes, J., & Ashford, M. L. J. (1999). Hydrogen peroxide induces intracellular calcium overload by activation of a non-selective cation channel in an insulin-secreting cell line. Journal of Biological Chemistry, 274, 833–841. doi:10.1074/jbc.274.2.833
- Hughes, F. M., Bortner, C. D., Purdy, G. D., & Cidlowski, J. A. (1997). Intracellular K + suppresses the activation of apoptosis in lymphocytes. Journal of Biological Chemistry, 272, 30567–30576. doi:10.1074/jbc.272.48.30567
- Jenner, P. (2003). Oxidative stress in Parkinson’s disease. Annals of Neurology, 53, S26–S36. discussion S36–8. doi:10.1002/ana.10483
- Josephson, R. A., Silverman, H. S., Lakatta, E. G., Stern, M. D., & Zweier, J. L. (1991). Study of the mechanisms of hydrogen peroxide and hydroxyl free radical-induced cellular injury and calcium overload in cardiac myocytes. Journal of Biological Chemistry, 266, 2354–2361. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/1846625
- Juurlink, B. H., & Paterson, P. G. (1998). Review of oxidative stress in brain and spinal cord injury: Suggestions for pharmacological and nutritional management strategies. Journal of Spinal Cord Medicine, 21, 309–334. doi:10.1080/10790268.1998.11719540
- Kadas, D., Ryglewski, S., & Duch, C. (2015). Transient BK outward current enhances motoneurone firing rates during Drosophila larval locomotion. The Journal of Physiology, 593, 4871–4888. doi:10.1113/JP271323
- Karunanithi, S., Barclay, J. W., Robertson, R. M., Brown, I. R., & Atwood, H. L. (1999). Neuroprotection at Drosophila synapses conferred by prior heat shock. The Journal of Neuroscience, 19, 4360–4369. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/10341239 doi:10.1523/JNEUROSCI.19-11-04360.1999
- Komatsu, A., Singh, S., Rathe, P., & Wu, C. F. (1990). Mutational and gene dosage analysis of calcium-activated potassium channels in Drosophila: Correlation of micro- and macroscopic currents. Neuron, 4, 313–321. doi:10.1016/0896-6273(90)90105-O
- Krans, J. L., Parfitt, K. D., Gawera, K. D., Rivlin, P. K., & Hoy, R. R. (2010). The resting membrane potential of Drosophila melanogaster larval muscle depends strongly on external calcium concentration. Journal of Insect Physiology, 56, 304–313. doi:10.1016/j.jinsphys.2009.11.002
- Krill, J. L., & Dawson-Scully, K. (2016). CGMP-dependent protein kinase inhibition extends the upper temperature limit of stimulus-evoked calcium responses in motoneuronal boutons of Drosophila melanogaster larvae. PLoS One, 11, e0164114. doi:10.1371/journal.pone.0164114
- Lancaster, B., & Nicoll, R. A. (1987). Properties of two calcium-activated hyperpolarizations in rat hippocampal neurones. The Journal of Physiology, 389, 187–203. doi:10.1113/jphysiol.1987.sp016653
- Lawson, K. (2000). Potassium channel openers as potential therapeutic weapons in ion channel disease. Kidney International, 57, 838–845. doi:10.1046/j.1523-1755.2000.00923.x
- Lee, J., Ueda, A., & Wu, C. F. (2008). Pre- and post-synaptic mechanisms of synaptic strength homeostasis revealed by slowpoke and shaker K + channel mutations in Drosophila. Neuroscience, 154, 1283–1296. doi:10.1016/j.neuroscience.2008.04.043
- Lee, J., Ueda, A., & Wu, C. F. (2014). Distinct roles of Drosophila cacophony and Dmca1D Ca2+ channels in synaptic homeostasis: Genetic interactions with slowpoke Ca2+-activated BK channels in presynaptic excitability and postsynaptic response. Developmental Neurobiology, 74, 1–15. doi:10.1002/dneu.22120
- Lehmann, H. F., & Jurkat, R. K. (1999). Voltage-gated ion channels and hereditary disease. Physiological Reviews, 79, 1317–1372. doi:10.1152/physrev.1999.79.4.1317
- Li, H. J., Zhang, Y. J., Zhou, L., Han, F., Wang, M. Y., Xue, M. Q., & Qi, Z. (2014). Chlorpromazine confers neuroprotection against brain ischemia by activating BKCa channel. European Journal of Pharmacology, 735, 38–43. doi:10.1016/j.ejphar.2014.04.017
- Liu, Y., Fiskum, G., & Schubert, D. (2002). Generation of reactive oxygen species by the mitochondrial electron transport chain. Journal of Neurochemistry, 80, 780–787. doi:10.1046/j.0022-3042.2002.00744.x
- Mantegazza, M., Curia, G., Biagini, G., Ragsdale, D. S., & Avoli, M. (2010). Voltage-gated sodium channels as therapeutic targets in epilepsy and other neurological disorders. The Lancet Neurology, 9, 413. doi:10.1016/S1474-4422(10)70059-4
- McKay, M. C., Dworetzky, S. I., Meanwell, N. A., Olesen, S. P., Reinhart, P. H., Levitan, I. B., … Gribkoff, V. K. (1994). Opening of large-conductance calcium-activated potassium channels by the substituted benzimidazolone NS004. Journal of Neurophysiology, 71, 1873–1882. doi:10.1152/jn.1994.71.5.1873
- McNaughton, N. C. L., Green, P. J., & Randall, A. D. (2001). Inhibition of human α1E subunit-mediated Ca2+ channels by the antipsychotic agent chlorpromazine. Acta Physiologica Scandinavica, 173, 401–408. doi:10.1046/j.1365-201X.2001.00914.x
- Meera, P., Wallner, M., Song, M., & Toro, L. (1997). Large conductance voltage- and calcium-dependent K + channel, a distinct member of voltage-dependent ion channels with seven N-terminal transmembrane segments (S0-S6), an extracellular N terminus, and an intracellular (S9-S10) C terminus. Proceedings of the National Academy of Sciences, 94, 14066–14071. doi:10.1073/pnas.94.25.14066
- Michelakis, E. D., McMurtry, M. S., Wu, X. C., Dyck, J. R., Moudgil, R., Hopkins, T. A., … Archer, S. L. (2002). Dichloroacetate, a metabolic modulator, prevents and reverses chronic hypoxic pulmonary hypertension in rats: Role of increased expression and activity of voltage-gated potassium channels. Circulation, 105, 244–250. doi:10.1161/hc0202.101974
- Misener, S. R., Chen, C.-P., & Walker, V. K. (2001). Cold tolerance and proline metabolic gene expression in Drosophila melanogaster. Journal of Insect Physiology, 47, 393–400. doi:10.1016/S0022-1910(00)00141-4
- Miyata, M., & Smith, J. D. (1996). Apolipoprotein E allele–specific antioxidant activity and effects on cytotoxicity by oxidative insults and β-amyloid peptides. Nature Genetics, 14, 55–61. doi:10.1038/ng0996-55
- Money, T. G. A., Rodgers, C. I., McGregor, S. M. K., & Robertson, R. M. (2009). Loss of potassium homeostasis underlies hyperthermic conduction failure in control and preconditioned locusts. Journal of Neurophysiology, 102, 285–293. doi:10.1152/jn.91174.2008
- Nani, F., Cifra, A., & Nistri, A. (2010). Transient oxidative stress evokes early changes in the functional properties of neonatal rat hypoglossal motoneurons in vitro. European Journal of Neuroscience, 31, 951–966. doi:10.1111/j.1460-9568.2010.07108.x
- Nimigean, C. M., & Magleby, K. L. (1999). The beta subunit increases the Ca2+ sensitivity of large conductance Ca2+-activated potassium channels by retaining the gating in the bursting states. The Journal of General Physiology, 113, 425–440. doi:10.1085/jgp.113.3.425
- Pallotta, B. S., Magleby, K. L., & Barrett, J. N. (1981). Single channel recordings of Ca2+-activated K + currents in rat muscle cell culture. Nature, 293, 471–474. doi:10.1038/293471a0
- Pandey, A., Vimal, D., Chandra, S., Saini, S., Narayan, G. K., & Chowdhuri, D. (2014). Long-term dietary exposure to low concentration of dichloroacetic acid promoted longevity and attenuated cellular and functional declines in aged Drosophila melanogaster. Age, 36, 1139–1154. https://doi.org/10.1007/s11357-014-9628-1
- Piskorowski, R., & Aldrich, R. W. (2002). Calcium activation of BKCa potassium channels lacking the calcium bowl and RCK domains. Nature, 420, 499–502. doi:10.1038/nature01199
- Quamme, G. A. (1997). Chlorpromazine activates chloride currents in Xenopus oocytes. Biochimica et Biophysica Acta – Biomembranes, 1324, 18–26. doi:10.1016/S0005-2736(96)00205-2
- Raffaelli, G., Saviane, C., Mohajerani, M. H., Pedarzani, P., & Cherubini, E. (2004). BK potassium channels control transmitter release at CA3–CA3 synapses in the rat hippocampus. The Journal of Physiology, 557, 147–157. doi:10.1113/jphysiol.2004.062661
- Renger, J. J., Yao, W. D., Sokolowski, M. B., & Wu, C. F. (1999). Neuronal polymorphism among natural alleles of a cGMP-dependent kinase gene, foraging, in Drosophila. The Journal of Neuroscience, 19, RC28. doi:10.1523/JNEUROSCI.19-19-j0002.1999
- Shen, K. Z., Lagrutta, A., Davies, N. W., Standen, N. B., Adelman, J. P., & North, R. A. (1994). Tetraethylammonium block of Slowpoke calcium-activated potassium channels expressed in Xenopus oocytes: Evidence for tetrameric channel formation. Pflügers Archive European Journal of Physiology, 426, 440–445 https://doi.org/10.1007/BF00388308
- Singh, S., & Wu, C. F. (1990). Properties of potassium currents and their role in membrane excitability in Drosophila larval muscle fibers. The Journal of Experimental Biology, 152, 59–76. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/2121887
- Smith, M. A., Herson, P. S., Lee, K., Pinnock, R. D., & Ashford, M. L. J. (2003). Hydrogen-peroxide-induced toxicity of rat striatal neurones involves activation of a non-selective cation channel. The Journal of Physiology, 547, 417–425. doi:10.1113/jphysiol.2002.034561
- Stefani, E., Ottolia, M., Noceti, F., Olcese, R., Wallner, M., Latorre, R., & Toro, L. (1997). Voltage-controlled gating in a large conductance Ca2+-sensitive K + channel (hslo). Proceedings of the National Academy of Sciences of the United States of America, 94, 5427–5431. doi:10.1073/pnas.94.10.5427
- Stewart, B. A., Schuster, C. M., Goodman, C. S., & Atwood, H. L. (1996). Homeostasis of synaptic transmission in Drosophila with genetically altered nerve terminal morphology. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 16, 3877–3886. Retrieved from https://doi.org/10.1523/JNEUROSCI.16-12-03877.1996
- Vetri, F., Choudhury, M. S. R., Pelligrino, D. A., & Sundivakkam, P. (2014). BKca channels as physiological regulators: A focused review. Journal of Receptor, Ligand and Channel Research, 7, 3–13. https://doi.org/10.2147/JRLCR.S36065
- White, R. E., Lee, A. B., Shcherbatko, A. D., Lincoln, T. M., Schonbrunn, A., & Armstrong, D. L. (1993). Potassium channel stimulation by natriuretic peptides through cGMP-dependent dephosphorylation. Nature, 361, 263–266. https://doi.org/10.1038/361263a0
- Yang, Y., Li, P.-Y., Cheng, J., Mao, L., Wen, J., Tan, X.-Q., … Zeng, X.-R. (2013). Function of BKCa channels is reduced in human vascular smooth muscle cells from Han Chinese patients with hypertension. Hypertension, 61, 519–525. https://doi.org/10.1161/HYPERTENSIONAHA.111.00211
- Zhang, B., & Ma, J. (2008). SERPINA3K prevents oxidative stress induced necrotic cell death by inhibiting calcium overload. PloS One, 3, e4077. https://doi.org/10.1371/journal.pone.0004077
- Zhang, J. (2016). Blockade of large conductance Ca(2+) activated K(+) channel may protect neuronal cells from hypoxia mimetic insult and oxidative stress. Acta Poloniae Pharmaceutica, 73, 895–902. Retrieved from https://europepmc.org/abstract/med/29648715
- Zhou, Y., Schopperle, W. M., Murrey, H., Jaramillo, A., Dagan, D., Griffith, L. C., & Levitan, I. B. (1999). A dynamically regulated 14-3-3, Slob, and Slowpoke potassium channel complex in Drosophila presynaptic nerve terminals. Neuron, 22, 809–818. https://doi.org/10.1016/S0896-6273(00)80739-4