
Abstract
Membrane proteins play a lead role in the formation and function of synapses, but, despite revolutions in immunology and molecular genetics, limitations persist in our ability to investigate membrane proteins in the context of an intact synapse. Here, we introduce a simple but novel approach to resolving the distribution of endogenous membrane proteins in either live or fixed tissues. The technique involves transgenic expression of a protein with an extracellular tag, a generic transmembrane domain, and an intracellular terminus that mimics the intracellular anchoring motifs of the endogenous protein of interest. We provide three examples where these kisser probes can be used to answer questions regarding the synaptic distribution of endogenous proteins and their microenvironment that would be difficult to resolve by other contemporary means: (i) the live distribution of untagged proteins at the neuromuscular junction (Cacophony and Shaker), (ii) the relative distribution of an untagged protein (PMCA) in pre- versus post-synaptic membranes separated by only 20 nm across the cleft of a fixed synapse, and (iii) the live targeting of functional probes (chemical and protein fluorescent pH reporters) to membrane protein-defined subcellular domains.
Introduction
Plasmamembrane (PM) proteins contribute to a variety of cellular processes that regulate neurodevelopment and synaptic transmission in neurons (von Heijne, Citation2007). PM proteins are commonly tagged to study their function and trafficking, either in live or fixed tissues, but there are shortcomings to this approach and some PM proteins (such as ion channels) cannot be easily tagged without perturbing their function. A host of tagging approaches was developed over the years, comprehensively reviewed by Crivat and Taraska (Citation2012) and Bolbat and Schultz (Citation2017). These include chemical tags (Jing & Cornish, Citation2011), peptide tags (Fritze & Anderson, Citation2000), fluorescent proteins (FPs), self-labeling protein tags (SNAP-tag, CLIP-tag, HALO-tag) and novel epitope tags such as Snorkel (Brown et al., Citation2013). As might be anticipated, there are caveats in the use of each. Generally, any translational fusion might affect folding and localization of the protein to which a peptide is appended by interfering with protein–protein interactions. Fusion proteins also disrupt cellular functions as observed in epitope-tagged ion channels (Maue, Citation2007). Overabundance caused by overexpression may also induce physiological changes (Shastry, Citation1995; Zhang et al., Citation2009). Although gene editing to incorporate a tag using the CRISPR/Cas9 technique can sidestep issues caused by overexpression, it remains a challenge to identify sites that will accommodate tags without disrupting protein function. Proteins, such as ion channels, have only a limited number of ‘permissive’ regions that can support protein tags without grossly affecting cell physiology (Watschinger et al., Citation2008).
Here, we introduce a novel approach for ‘associating’ tags with endogenous PM proteins to study their distribution in vivo while minimizing the impact on cellular function. The tags, called ‘kissers’, are based on endogenous proteins stripped of their functional domains, leaving ostensibly inert intracellular fragments. As the reporter tag is not fused with the targeted protein, the tag is less likely to disrupt the folding, trafficking, interaction, and function of the endogenous protein. Similarly, transgenic overexpression of kisser probes should be more benign than overexpression of the translational fusions that include the full endogenous protein, especially where there are multiple tightly regulated isoforms of that protein.
In the following sections, we demonstrate that kisser probes can be used to: (1) resolve the distribution of untagged proteins in vivo, (2) identify discrete pools of the same protein located in pre- versus post-synaptic membranes separated by a ∼20 nm synaptic cleft, and (3) target probes to protein-defined subcellular domains of the PM and perform live functional imaging within these domains. Each of these applications is illustrated with one or more probes based on proteins expressed endogenously at the Drosophila larval NMJ. However, as the kisser probes will compete with endogenous protein for the same protein interaction sites on intracellular scaffolds, their expression may have unintended physiological consequences. As such each kisser probe must be validated before use and may not be useful for every application, caveats shared with other translational fusions.
Methods
Fly lines
The fly lines used in this study are listed below, and unless indicated otherwise, each line was made in-house, with details provided in the section below: UAS-Cac-kisser-pHerry; UAS-Cac-kisser-FAP; UAS-Sh-kisser-FAP; UAS-PMCA-kisser-pHerry; UAS-PMCA-kisser-FAP; UAS-TagBFP (BDSC); CacsfGFP-N (developed by Gratz et al. Citation2018); UAS-mTFP1-CEPIA3; UAS-iRFP670-RCaMP1a; 24B-Gal4 (BDSC); nSyb-Gal4 (BDSC).
Maintenance of fly lines
All flies were kept on standard media. In every case, a single copy of a transgene was driven by a single copy of a driver. Crosses involving FAP-kisser probes were maintained at 28 °C to increase expression.
Cloning
The plasmid encoding the αFAPtag was purchased from Spectra Genetics, Inc. (Pittsburgh, PA). The αFAPtag gene was subcloned into backbones biosynthesized by Genescript and placed in the pJFRC14 plasmid (Cac-kisser-FAP, Sh-kisser-FAP, and PMCA-kisser-FAP). All backbones encoded (from 5′ end) a signal peptide from wingless (directs the recombinant protein to the secretory pathway) at the 5′ end, an MCS to accept FAPtags, protein tags placed directly upstream and downstream of the MCS (HA-tag and myc-tag, respectively), and a transmembrane (TM) domain of the human platelet-derived growth factor receptor (hPDGFR). Downstream of TM domain, the Cac-kisser backbone contained the C-terminus of Cac, with scrambled PDZ binding sites and CaM binding domain, and was terminated with first the PDZ domain of RIM and then the SH3 binding motif of RIM. In the Sh-kisser-FAP construct, an unmodified C-terminus of Shaker (Sh) was cloned downstream of the TM domain. Finally, the C-terminal portion of PMCA-kisser backbone encoded a modified C-terminus of Drosophila melanogaster PMCA. The CaM binding site in the PMCA C-terminus was scrambled, to prevent unwanted oligomerization (Benetti et al., Citation2011) and cleavage of PMCA-kisser by calpain (Di Leva, Domi, Fedrizzi, Lim, & Carafoli, Citation2008). PMCA-kisser-pHerry and Cac-kisser-pHerry probes were developed by subcloning pHerry into the MCS. Plasmids encoding mTFP1, CEPIA3, iRFP670 and RCaMP1a proteins were obtained from Addgene (Cambridge, MA). The myr-mTFP1-CEPIA3 and myr-iRFP670-RCaMP1a constructs were generated by subcloning the genes encoding the respective proteins into a backbone biosynthesized by Genescript and placed in the pJFRC14 plasmid. The backbone encoded an N-terminal myristoylation signal peptide and two MCSs separated by a flexible linker. The upstream MCS was used to subclone mTFP1 or iRFP670, while the downstream one (C-terminal to the first) was used to subclone CEPIA3 (with mitochondrial signal sequence removed) or RCaMP1a, respectively. Completed constructs were purified with QIAGEN Plasmid Maxi Kits and injected into Drosophila embryos harboring the transposable element insertion site attP2, by Rainbow Transgenic Flies. Transgenes on the 3rd chromosome were balanced over TM6B, Tb1.
Preparation of specimens for live imaging and immunocytochemistry
With the exception of Cac-kisser-FAP animals, all experiments were performed on female 3rd instar larvae. In Cac-kisser-FAP animals, the expression of the Cac-kisser-FAP construct was consistently higher in the males (despite all transgenes being on autosomes) and therefore the experiments were performed on 3rd instar male larvae. Briefly, the larvae were fillet-dissected in cold Schneider’s insect medium (Sigma, St. Louis, MO). In live staining experiments, the brain hemispheres and ventral ganglia were removed to limit contractions. Segmental nerves were trimmed and the preparation was incubated for 30 min at room temperature (RT) with the fluorogen staining solution (Schneider’s medium, 100 nM Malachite Green-based fluorogen). The preparation was then washed thrice with cold HL3 and imaged with a Nikon A1 confocal microscope fitted with a 100× water immersion (1.1 NA) objective. For immunocytochemistry, the samples were stained for 30 min with a fluorogen and then briefly fixed (10 min) with 4% PFA at RT. The samples were then washed four times with 1× PBS at RT and immune-stained with antibodies against a subset of Brp, Dlg, PMCA, HA-tag and HRP. Nc82 (anti-Brp) and 4F3 (anti-Dlg) were procured from the Developmental Studies Hybridoma Bank, anti-HA antibody (Roche, Basel, Switzerland), and anti-HRP antibodies were purchased from Abcam (Cambridge, UK), and the anti-PMCA antibody was a gift from Dr. Greg Lnenicka. Fluorogens were purchased from Spectra Genetics, Inc. (Pittsburgh, PA). All fixed immunohistochemically processed preparations were examined under a glass coverslip using a 100× oil (1.3 NA) objective on a Nikon A1 confocal microscope fitted with GaAsP detectors. Preparations were scanned sequentially starting with the longest wavelengths and progressing to the shortest (640 nm, 561 nm, 488 nm, 405 nm). All bouton images represent a collapsed Z-series that encompasses the entire depth of boutons (except for , which represents a collapsed Z-series of four sections 0.4 µm apart, through the center of the bouton). Colocalization of Cac-kisser-pHerry and Brp was analyzed with the Colocalization Threshold plugin in ImageJ. Statistical significance of the Pearson colocalization coefficients calculated for each value was tested with permutation tests. To analyze whether PMCA-kisser-pHerry is relatively enriched in the SSR in comparison to the rest of the muscle, we compared the distribution ratio of PMCA-kisser-pHerry with that of myristoylated iRFP670-RCaMP1a and mTFP1-CEPIA3. Z-stacks encompassing the entire depth of boutons were collected, then collapsed into one plane by summing pixel fluorescence intensities across all slices. An ROI was selected just peripheral to the bouton in a region that would be coincident with the SSR, along with an ROI identical in area representing muscle PM, but not within the SSR. The average fluorescence intensities of either SE-pHluorin and iRFP670, or mCherry and mTFP1, were measured in these ROIs, and the SSR to non-SSR ratios were then calculated for each protein.
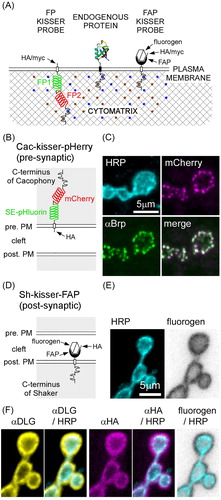
Figure 1. Kisser probes mimic the subcellular localization of endogenous proteins. (A) Schematic representation of kisser probes described in the manuscript. The fluorescent protein-based kisser probe (FP kisser) encodes two intracellular fluorescent proteins (FP1 and FP2) SE-pHluorin fused to mCherry (pHerry), while the fluorogen-activating peptide (FAP)-based probe (FAP-kisser) encodes an extracellular FAPtag. Kisser probes bind intracellular scaffolds through C-terminal tails homologous to endogenous proteins. Peptide tags (e.g. myc-tag and HA-tag) enable immunodetection. (B) A diagram of Cac-kisser-pHerry, a probe designed to localize with endogenous Cac. (C) A fixed type-Ib bouton where Cac-kisser-pHerry expression is driven pre-synaptically (nSyb-GAL4): Cac-kisser-pHerry (mCherry fluorescence) overlaps with the distribution of endogenous Brp (immunohistochemistry; Cy5). HRP staining (immunohistochemistry; DyLight 405) was used to visualize motor neuron terminals. (D) A diagram of post-synaptic Sh-kisser-FAP, a probe designed to localize with endogenous Sh. The crossed hexagon indicates the fluorogen bound to the FAPtag. (E) A fixed type-Ib bouton where Sh-kisser-FAP expression is driven post-synaptically (24B-GAL4), and the associated fluorogen (Malachite Green; ex. 635 nm/em. 660 nm) labelling. HRP staining (immunohistochemistry; DyLight 405) was used to visualize motor neuron terminals. (F) Same preparation as in E: HA-tag (immunohistochemistry; Cy3) staining was used to detect post-synaptic Sh-kisser-FAP. The probe overlaps in its expression with endogenous Dlg (immunohistochemistry; AF488) and surrounds motor neuron terminals visualized with HRP staining as does the fluorogen (from E).
Preparation of specimens for functional imaging
For pH imaging experiments, the ventral ganglion was transected close to the brain hemispheres and the brain hemispheres were removed, while the segmental nerves were left attached to the remaining part of the ventral ganglion. A loop of one segmental nerve projecting to abdominal segment 4 was drawn into a suction pipette. Schneider’s medium was replaced with HL6 supplemented with 2 mM Ca2+ and 100 µM ryanodine (Enzo Life Sciences, Farmingdale, NY). The preparation was then allowed to equilibrate for a few minutes and then moved to the imaging rig. Imaging was performed on a wide-field Nikon microscope fitted with a 100× water immersion (1.1 NA) objective. Samples were illuminated with a Lumencor fluorescent light source and the fluorescence emission was captured with two Andor iXon3 CCD cameras (DU-897) fitted to a TuCam beam-splitter. PMCA-kisser-FAP preparations were excited with 568 and 640 nm light. PMCA-kisser-pHerry preparations were excited with 488 and 568 nm light (pHluorin/mCherry). Separate emission wavelengths were collected by separate cameras, with sequential excitation of fluorophores used to minimize channel cross-talk. During analysis both channels were treated as if recorded at the same time. 568/640 (FAP) and 488/568 (pHerry) emission ratios were calculated. Imaging and electrical stimulation components were controlled through a PC running NIS Elements software (Nikon, Tokyo, Japan). Timing of electrical stimulation was controlled by a Master-8 controller (A.M.P.I., Israel) triggered by PC software, with pulses generated by a constant voltage stimulator (model DS2A-Mk. II, Digitimer Ltd., Cambridge, UK). Changes in pH in the synaptic cleft and the cytosol were elicited by a train of electrical stimuli impulses (2 V, 0.3 ms each) delivered to the nerve at 30 Hz for 1 s. Average fluorescence at 1–2 s prior to stimulus delivery was treated as baseline and used to calculate ΔF values for individual channels. ΔR was calculated from ratio values normalized to the baseline value.
Electrophysiology
All larvae were dissected in chilled HL6 (no Ca2+) and switched to HL3 (2 mM Ca2+ and 15 mM Mg2+) before electrophysiology. Recordings were conducted 5–10 min after transfer of HL3 solution. For post-synaptic expression of PMCA-kisser-FAP, preparations with and without dye were incubated for 30 min at RT in HL6 before recordings, and dye staining was confirmed via subsequent confocal microscopy. For pre-synaptic expression of PMCA-kisser-pHerry, HL3 was added immediately to preparations after dissections. Current clamp recordings were made from muscle 6 in abdominal segment 4. A loop of the segmental nerve projecting into segment 4 was drawn into suction pipette for stimulation. Signals were detected, digitized and recorded using an Axoclamp 900 A amplifier (Molecular Devices, Sunnyvale, CA) connected to a 4/35 PowerLab (ADInstruments, Colorado Springs, CO) and a PC running LabChart v8.0. Micropipettes were pulled from borosilicate capillary tubing (Sutter Instruments, Novato, CA) on a Flaming/Brown P-97 micropipette puller, and filled with a 1:1 mixture of 3 M KCl and 3 M KAc. A 0.1 gain head-stage was used for recording. Recording electrode resistance was 20–40 MΩ. Peak-to-peak noise was <0.15 mV after 2 kHz low-pass filtering. Only records from muscle fibers that maintained a resting membrane potential (RMP) of greater than –65 mV were analyzed. Quantal content (QC) was calculated as follows: QC = average EJP amplitude/average mEJP amplitude, where average EJP amplitude is the average amplitude of 15 consecutive EJPs collected at 0.2 Hz and average mEJP amplitude is the average amplitude of >30 consecutive mEJPs. Average EJP amplitude was corrected for non-linear summation (McLachlan & Martin, Citation1981). One-way ANOVA was used on PMCA-kisser-FAP experiments, while PMCA-kisser-pHerry experiments were analyzed with Student’s t-test.
Results
Kisser probes are designed to mimic the localization of specific PM proteins. Their design has three basic elements: an intracellular tail based on the protein of interest, a generic TM region, and a reporter tag (). Subcellular localization of PM proteins is determined through the binding of protein scaffolds close to the inner leaflet of the PM. Scaffold-binding domains are most often present on N- and C-terminals of PM proteins, although in some cases interaction sites may be located within intracellular loops between PM-spanning regions. The former are the most amenable to be exploited using the kisser probe design, although it is possible to construct hybridized kissers sharing binding domains of different proteins or using localization determinants located within intracellular loops. Regions that do not confer localization specificity (such as calmodulin (CaM) binding, oligomerization, and protease cleavage sites) are removed from kisser probes. The intracellular domain is fused to a TM region of the hPDGFR (Gronwald et al., Citation1988) and a reporter tag. The reporter tags can reside in either the extracellular milieu or the cytosol (Figure S1).
Our kisser probes use either FPs or fluorogen activating peptides (FAPs) as reporter tags. FP-based kisser probes (, left) were designed to enable functional pH imaging using an intracellular pH sensitive ratiometric protein called pHerry. pHerry is a translational fusion of pH-sensitive Super-ecliptic pHluorin (SE-pHluorin) and pH-insensitive (near neutral) mCherry (Rossano, Kato, Minard, Romero, & Macleod, Citation2017). FAP-based kisser probes (, right) were designed with a FAPtag facing the extracellular milieu (Szent-Gyorgyi et al., Citation2008; Szent-Gyorgyi, Schmidt, Fitzpatrick, & Bruchez, Citation2010). The FAP ligand is a fluorogen which is not fluorescent unless bound to the FAPtag. A range of fluorogens are available (Spectra Genetics, Inc., Pittsburgh, PA); green or red emitting, cell permeant or impermeant, pH sensitive or pH insensitive. FAP-based kisser probes are therefore hybrid sensors with the brightness and kinetics of chemical dyes and the specificity of genetically encoded reporters. FAP-based kisser probes can also be modified to support an intracellular GFP, which enables the localization of probes prior to fluorogen binding.
Kisser probes are particularly useful in the study of the subcellular organization of synapses. As with the tagging of other proteins, synaptic protein tagging is not straight-forward as the tags can be placed in only a limited number of places without adversely affecting synaptic physiology. We focused on three proteins expressed at Drosophila larval NMJ to demonstrate the utility of kisser probes: a pre-synaptic voltage-gated calcium channel Cacophony (Cac), a post-synaptic potassium channel Sh, and the plasma membrane Ca2+-ATPase (PMCA) which is expressed on both sides of the synaptic cleft.
Using kisser probes to examine the distribution of Cacophony and Shaker at the larval NMJ
We compared the subcellular distribution of Cac-kisser probes (Figures S2–S7) and Sh-kisser probes ( Figures S8–S10) with the distribution of their respective endogenous proteins, along with proteins known to exist in the same molecular complex at the Drosophila larval NMJ.
The Cac-kisser probe was originally designed to include only the PDZ domain and SH3 binding motif of RIM, a scaffolding protein involved in the localization of Cac to active zones (Graf et al., Citation2012). The PDZ domain binds directly to Cac (Kaeser et al., Citation2011), while the SH3 binding motif interacts with Rim binding protein (RBP) (Hibino et al., Citation2002), another scaffolding protein involved in the organization of active zones at the Drosophila larval NMJ. The inclusion of these two domains resulted in only a modest localization of the probe at active zones (compared with a PM-anchored FP), with a significant fraction of the probe dispersed uniformly within the bouton (data not shown). The probe was subsequently modified to include the C-terminus of Cac which binds RIM and RBP, and while this improved localization of Cac-kisser to active zones, it likely did so at the expense of Cac-kisser’s specificity for Cac itself. The CaM binding domain and PDZ binding sites present in the C-terminus of endogenous Cac were scrambled to prevent spurious interactions (Figures S2 and S3).
We compared the expression pattern of Cac-kisser-pHerry () with that of Bruchpilot (Brp), a protein that promotes active zone assembly and Ca2+ channel clustering (Kittel et al., Citation2006). In larval NMJ type-Ib boutons of fixed preparations, the intracellular mCherry signal of Cac-kisser-pHerry is in close register with Brp defined active zones () (r = 0.809 ± 0.031, mean ± SD; N = 6; Figure S4). This result is consistent with Cac-kisser-pHerry recognizing the microenvironment of Cac as Gratz et al. (Citation2018) reported a high degree of correlation between levels of Cac and Brp. There have been reports of mCherry being mislocalized to endosomal compartments (Costantini et al., Citation2015), in which case, a translational fusion such as Cac-kisser-pHerry containing mCherry might become mislocalized. Highly fluorescent structures were observed in the somata of motor neurons expressing Cac-kisser-pHerry, but such structures were mostly excluded from motor neuron axons (Figure S5). In separate analyses, we compared the distribution of SE-pHluorin of Cac-kisser-pHerry with Brp at the NMJ, and here too we found Cac-kisser-pHerry to localize to active zones (Figure S6).
We also designed a FAP-based Cac-kisser probe, with the same C-terminus that provided improved localization of Cac-kisser-pHerry to the PM, but without intracellular FPs, and with a FAPtag facing the extracellular milieu (Figure S7(A)). At the NMJ of fixed and permeabilized preparations, we compared the expression pattern of Cac-kisser-FAP with that of endogenous Cac-bearing super-folder GFP at its N-terminus (CacsfGFP-N) (Gratz et al., Citation2018). To generate the CacsfGFP-N fly line, the O’Connor-Giles laboratory targeted the translational start site of the cac locus for CRISPR-mediated incorporation of sfGFP, and this resulting in tagging the full complement of endogenous Cac channel isoforms. They also established that CacsfGFP-N localizes to active zones as predicted by previous studies with ectopically expressed UAS-CacGFP (Kawasaki, Zou, Xu, & Ordway, Citation2004; Rasse et al., Citation2005). While Cac-kisser-FAP formed puncta that were usually coincident with CacsfGFP-N (arrowheads), more than half of the CacsfGFP-N puncta were unaccompanied by a Cac-kisser-FAP punctum (open arrowhead), and in some cases Cac-kisser-FAP formed punctae in the absence of CacsfGFP-N punctae (solid arrowheads) (Figure S7(B)). It is quite possible that these features are the result of mislocalization or oligomerization of the FAP-based probe. Intriquingly, however, while edited endogenous Cac has been shown to reside at every Brp-defined active zone (Gratz et al., Citation2018), and the GFP-tagged Cac transgene can also be detected at every active zone (Akbergenova, Zhang, Weiss-Sharabi, Cunningham, & Littleton, Citation2017; Rasse et al., Citation2005), the Drosophila Cac antibody reveals an endogenous Cac distribution pattern remarkably similar to the pattern revealed by Cac-kisser-FAP (Chang, Hazelett, Stewart, & Morton, Citation2014). Application of the membrane impermeant fluorogen Malachite-Green to the live preparation revealed a similar pattern to that seen in the fixed and permeabilized tissue (Figure S7(C,D)).
The FAP-based Sh-kisser probe contains the minimal region necessary for Sh to maintain its subcellular localization (Figures S8 and S9) in the sub-synaptic reticulum (SSR) of the NMJ (Guerrero et al., Citation2005; Peled & Isacoff, Citation2011; Zito, Fetter, Goodman, & Isacoff, Citation1997). Live imaging using the Malachite Green fluorogen () revealed that Sh-kisser-FAP is preferentially transported to the SSR, similarly to the endogenous Sh (Tejedor et al., Citation1997) and a truncated Sh-tagged with CD8 (Zito et al., Citation1997). We compared the distribution of Sh-kisser-FAP relative to that of Dlg, a protein involved in the organization of SSR beneath the type-I boutons at the larval NMJ () and necessary for the proper intracellular localization of endogenous Sh (Tejedor et al., Citation1997; Zito et al., Citation1997). Fluorogen labelling, followed by immunocytochemistry against Dlg, shows that the subcellular distribution of Sh-kisser-FAP probe overlaps with that of Dlg, and, by extension, Sh (). Immunostaining against the HA-tag in Sh-kisser-FAP also demonstrates the specificity of Sh-kisser-FAP for Dlg, and Sh (). Evidence for the specificity of Sh-kisser-FAP is also found when examining different terminal types. Sh is expressed strongly in the SSR of type-Ib boutons and to a lesser degree in the SSR of type-Is boutons and is excluded from any SSR of type-II boutons on muscle 13, similar to Dlg (Lahey, Gorczyca, Jia, & Budnik, Citation1994; Menon, Carrillo, & Zinn, Citation2013; Wang et al., Citation2011), and fluorogen labelling of Sh-kisser-FAP also reflected this distribution (Figure S10).
Using kisser probes to examine the relative distribution of proteins in closely spaced cellular compartments
The distribution of the PMCA in the pre-synaptic PM relative to the PMCA in the post-synaptic PM is unknown, as the application of antibodies and subsequent confocal microscopic examination cannot resolve the two populations of proteins separated only by a 20 nm synaptic cleft. Super-resolution microscopy would similarly struggle to resolve the relative distribution of the closely apposed, but discrete populations. Kisser probes can be deployed to resolve these populations.
The PMCA is essential for maintaining neurotransmission, as it prevents the build-up of intracellular Ca2+. At Drosophila larval NMJ it is the major route for the removal of pre-synaptic Ca2+ subsequent to motor activity (Lnenicka, Grizzaffi, Lee, & Rumpal, Citation2006), and also plays a substantial role in removing Ca2+ from the post-synaptic compartment (Desai & Lnenicka, Citation2011). The PMCA is a TM protein with both N- and C-terminals extending into the cytosol. The N-terminus has a mostly regulatory function (Di Leva et al., Citation2008), while the C-terminus governs the localization of the protein (Lopreiato, Giacomello, & Carafoli, Citation2014; Rimessi et al., Citation2005). A CaM binding site in the C-terminus is involved in the oligomerization of PMCA (Benetti et al., Citation2011; Falchetto, Vorherr, & Carafoli, Citation1992; Lopreiato et al., Citation2014). An alternate pathway for PMCA regulation involves calpain cleavage within the CaM binding domain (Di Leva et al., Citation2008; Wang, Roufogalis, & Kuo, Citation1991). PMCA-kisser probes were fitted with an engineered C-terminus of endogenous PMCA, and the CaM binding site was scrambled (Figure S11).
Unlike antibodies, kisser probes can be specifically targeted to either side of the NMJ. We pre-synaptically targeted PMCA-kisser-FAP () and PMCA-kisser-pHerry () through expression in motor neurons (nSyb-Gal4). Expression of the FAP-based probe resulted in an overall lower staining intensity (, inverted grayscale) in comparison to PMCA-kisser-pHerry () despite being expressed under the control of the same driver. While the fluorogen labeling of PMCA-kisser-FAP is somewhat granular (, inverted grayscale), this pattern is not observed in the stronger αHA stain of PMCA-kisser-pHerry (). The finding that pre-synaptically expressed PMCA-kisser-FAP mirrors the distribution of antibody revealed endogenous PMCA (), was somewhat surprising, as the antibody would be expected to bind both pre- and post-synaptic PMCA. A possible explanation for this, is that PMCA-kisser-FAP is indeed found in the post-synaptic membrane, and that this is due to pre-synaptic membrane shedding during synaptic growth, a phenomenon previously described for pre-synaptic TM proteins at the larval NMJ (Fuentes-Medel et al., Citation2009).
Figure 2. Kisser probes can be used to study the subcellular localization of a protein found in closely apposed membranes of two different cells. (A) A diagram of PMCA-kisser-FAP in the pre-synaptic membrane, a probe designed to localize with endogenous PMCA. The crossed hexagon indicates a fluorogen bound to a FAPtag. (B) A fixed type-Ib bouton where PMCA-kisser-FAP expression is driven in motor neurons (nSyb-GAL4): HRP staining (immunohistochemistry AF488) was used to visualize motor neuron terminals, PMCA-kisser-FAP was stained with Malachite Green. (C) Same preparation as in B: endogenous Brp (immunohistochemistry; AF405), endogenous PMCA (immunohistochemistry; Cy3). Note that endogenous PMCA staining identifies pre- and post-synaptic populations of PMCA, while PMCA-kisser-FAP stained with Malachite Green is pre-synaptic only. (D) A diagram of PMCA-kisser-pHerry in the pre-synaptic membrane. (E) A fixed type-Ib bouton where PMCA-kisser-pHerry expression is driven in motor neurons (nSyb-GAL4): HRP (immunohistochemistry; DyLight 405) staining indicates motor neuron terminals, HA-tag (immunohistochemistry; AF488) was used to detect pre-synaptic PMCA-kisser-pHerry, endogenous PMCA (immunohistochemistry; Cy5). Note that endogenous PMCA staining identifies pre- and post-synaptic populations of PMCA, while HA-tag staining indicates only the pre-synaptic PMCA-kisser-pHerry. Pre-synaptic PMCA-kisser-pHerry shows synaptic and peri-synaptic localization. (F) A diagram PMCA-kisser-pHerry in the post-synaptic membrane. (G) A fixed type-Ib bouton where PMCA-kisser-pHerry expression is driven in the muscle (24B-GAL4): HRP (immunohistochemistry; DyLight 405) staining indicates motor neuron terminals, SE-pH (immunohistochemistry; anti-GFP; AF488) fluorescence indicates the subcellular localization of post-synaptic PMCAkisser-pHerry, endogenous GluRIII (immunohistochemistry; Cy5) indicates the syb-synaptic reticulum. Post-synaptic PMCA-kisser-pHerry has a peri-synaptic localization, partially overlapping with endogenous GluRIII.
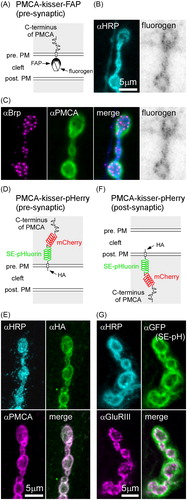
When PMCA-kisser-pHerry is expressed post-synaptically (), the distribution of SE-pHluorin is peripheral to GluRIII, which defines the inner extent of the post-synaptic SSR (). In a test of the possibility that the post-synaptically expressed PMCA-kisser probes are evenly distributed over the PM of the muscle, rather than being concentrated in the PMCA microenvironment of the SSR, we compared PMCA-kisser probe distribution with the distribution of two proteins (translational fusions) that attach to the inner leaflet of the PM through myristoylation (mTFP1-CEPIA3 and iRFP670-RCaMP1a). In both cases, the PMCA-kisser probes were significantly enriched in the SSR relative to the two proteins more generally associated with the PM (Figure S12). Somewhat unexpectedly then, while the post-synaptically expressed PMCA-kisser probe likely defines the post-synaptic extent of the PMCA in the SSR, the pre-synaptically expressed PMCA-kisser probe localizes to both the pre-synaptic membrane and the post-synaptic SSR.
Live targeting of functional probes to protein-defined subdomains of PM
As the PMCA provides the main route for Ca2+ clearance on either side of the NMJ, and antiports two protons (2H+) for each calcium ion (Ca2+) pumped (Thomas, Citation2009), pH microdomains might form on either side of the synaptic membranes. Kisser probes can be used to study such phenomena by targeting functional reporters to protein-defined subdomains of synaptic membranes. We targeted a pH-sensitive PMCA-kisser to the synaptic cleft (PMCA-kisser-FAP; ), which was tagged with a pH-sensitive ratiometric fluorogen (se-Red-pH; Spectra Genetics, Inc., Pittsburgh, PA). The pH-sensitive fluorogen is alternately excited at its absorbance maxima (551 nm and 633 nm) () and the fluorescence emitted when excited at either excitation wavelength is inversely proportional to pH. A train of electrical impulses delivered to the nerve at 30 Hz for 1 s, evoked pH change in the synaptic cleft of type-Ib boutons. The synaptic cleft alkalinized, with the fluorescence of the fluorogen decreasing in both imaging channels (Δ551 nm =11.36 ± 1.59%, Δ633 nm =1.31 ± 1.23%, mean ± SEM; ). The stimulus resulted in a ∼7% decrease in the fluorescence intensity ratio of PMCA-kisser-FAP (ΔR = 6.86 ± 1.46%, mean ± SEM; N = 6, p<.001, Student’s t test; ), representing a transient activity-evoked alkalization of the synaptic cleft. The cleft pH returns to baseline value after several seconds.
Figure 3. Kisser probes enable functional imaging in protein-defined subdomains of the plasma membrane (PM). (A) A diagram of post-synaptic PMCA-kisser-FAP. The hexagon represents a ratiometric pH-sensitive fluorogen bound to the probe's FAPtag. (B) PMCA-kisser-FAP expressed in the muscle (24B-GAL4), stained with the fluorogen, and visible in the sub-synaptic reticulum of type-Ib boutons. Panels show fluorescence of the fluorogen excited at 551 nm and 633 nm. (C) A 1 s 30 Hz stimulus train causes a decrease (note reversed axes) in fluorogen fluorescence at both wavelengths. A 10% decrease in the 551/633 ratio indicates a transient alkalization of the synaptic cleft. The re-acidification of the synaptic cleft extends beyond 1s post-train cessation. Traincommenced at time = 0. ΔF − change in fluorescence intensity, Δratio − change in ratio of emission wavelength intensities. (D) Untagged post-synaptic PMCA-kisser-FAP and tagged post-synaptic PMCA-kisser-FAP have no detectable impact on neurotransmission: EJP amplitudes are identical to driver control and quantal content is similar as well. (E) A diagram of pre-synaptic PMCA-kisser-pHerry. (F) PMCA-kisser-pHerry expressed in motor neurons and visible in type-Ib boutons (nSyb-GAL4): SE-pHluorin is pH-sensitive; mCherry is a ratiometric control. (G) A 1 s 30 Hz stimulus train causes a decrease in SE-pHluorin fluorescence with little change in mCherry fluorescence. A 4% decrease in the SEpH/mCherry ratio indicates a transient acidification of the pre-synaptic cytosol. The re-alkalization of the pre-synaptic cytosol extends beyond 1 s post-train cessation and is much slower than the corresponding cleft re-acidification. The stimulus commenced at time = 0. ΔF − change in fluorescence intensity, Δratio − change in the ratio of SEpH/mCherry emission intensities. (H) Pre-synaptic PMCA-kisser-pHerry has no detectable impact on neurotransmission: EJP amplitude is identical to driver control and quantal content is similar as well. (D and H) Box plots show first and third quartiles; error bars represent standard deviation.
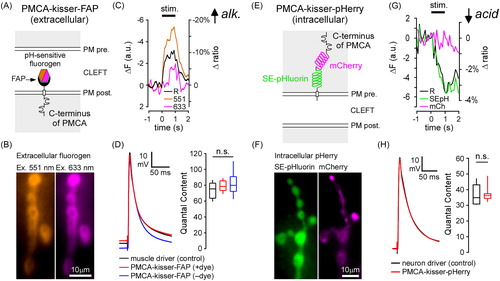
We targeted another pH sensitive probe, PMCA-kisser-pHerry () to the cytosol of the type-Ib boutons. A 30 Hz stimulus train resulted in an immediate decrease in SE-pHluorin fluorescence, which indicates cytosolic acidification (ΔSE-pHluorin =3.28 ± 0.40%%, mean ± SEM; ), while mCherry fluorescence was unchanged (ΔmCherry=–0.15 ± 0.44%, mean ± SEM; ). The stimulus train resulted in a ∼3.5% decrease in the fluorescence intensity ratio of PMCA-kisser-pHerry (ΔR = 3.41 ± 0.65%, mean ± SEM; N = 6, p<.01, Mann–Whitney’s Rank Sum test; ). Re-alkalization of the cytosol () is much slower than the corresponding re-acidification of the synaptic cleft ().
The PMCA-kisser probes have little impact on neurotransmission. Excitatory junction potentials (EJPs) of animals expressing PMCA-kisser-FAP are indistinguishable from those of muscle driver controls (+dye: 65.25 ± 1.18 mV, N = 7; –dye: 65.41 ± 1.60 mV, N = 6; driver control: 64.79 ± 2.62 mV, N = 6; mean ± SEM; EJP amplitudes corrected for non-linear summation; p=.972, one-way ANOVA; left panel). Furthermore, as miniature EJP (mEJP) amplitude is no different between animals (+dye: 0.68 ± 0.03 mV, N = 7; –dye: 0.71 ± 0.04 mV, N = 6; driver control: 0.75 ± 0.05 mV, N = 6; mean ± SEM; p=.544, one-way ANOVA), QC is unchanged between PMCA-kisser-FAP animals and muscle driver controls (+dye: 94.25 ± 3.32, N = 7; –dye: 94.72 ± 6.85, N = 6; driver control: 88.34 ± 6.03, N = 6; mean ± SEM; p=.666, one-way ANOVA; , right panel). Expression of PMCA-kisser-pHerry similarly has little influence on EJP amplitudes (67.37 ± 2.25 mV, N = 7; driver control: 67.64 ± 1.64 mV, N = 6; mean ± SEM; p=.926 Student’s t test; left panel) and mEJP amplitudes (1.60 ± 0.06 mV, N = 7; driver control: 1.67 ± 0.12 mV, N = 6; mean ± SEM; p=.616, Student’s t test) and QC is also unchanged (42.48 ± 2.30, N = 7; driver control: 41.54 ± 2.67 mV, N = 6; mean ± SEM; p=.794, Student’s t test; , right panel).
Discussion
The kisser probes described here are modular proteins that fuse an intracellular domain mimicking the localization-defining portions of an endogenous protein with a generic TM domain and an immunological tag. This approach appears to have a minimal impact on the trafficking and function of endogenous proteins, while enabling kisser probes to mimic their localization. Kisser probes enable the study of the live distribution of untagged proteins in live and fixed larval NMJs ( and , Figures S5–S7 and S10). In theory, kissers also allow analysis of the relative distribution of a single protein in pre- and post-synaptic membranes separated only by a narrow synaptic cleft, something which could not be done with an antibody. However, we were unable to resolve the relative distribution of the protein selected here, PMCA (). A possible explanation for this failure may be due to the FAPtag of pre-synaptic PMCA-kisser-FAP being shed and becoming incorporated in the post-synaptic membrane in the same manner as other membrane-tethered proteins at the larval NMJ (Fuentes-Medel et al., Citation2009). The kisser probes also enable live targeting of functional probes with a chemical or genetically encoded reporter, to membrane-defined subcellular domains. We performed pH imaging at PMCA-defined domains of the PM and found that the pre-synaptic cytosol acidifies in an activity-evoked manner, consistent with Rossano, Chouhan, and Macleod (Citation2013), while the synaptic cleft briefly alkalinizes (described more fully by Stawarski et al., in preparation) (). In a limited electrophysiological examination, we were unable to detect any impact of the kisser probes on neurotransmission ().
The kisser approach is not without limitations. The kisser probes essentially compete with the endogenous protein for the same binding sites on intracellular scaffolds. This competition may result in a partial or complete depletion of the endogenous protein at the site of its presumed action and therefore a disruption of cellular function. If the endogenous protein exists within a multi-protein complex, its replacement with a kisser probe will also lead to unintended physiological consequences. The intracellular domains of kisser probes are modified to remove sites that do not confer localization specificity. These modifications may affect higher order structures of the peptide chain and thus alter the interacting partners of the kisser probe. Functional domains also overlap (as is the case with calpain cleavage sites and CaM binding domains), and therefore such modifications of the peptide chain may result in the removal of protein binding sites and a disrupted localization.
Our ability to assess which Cac-kisser probe was more successful in locating endogenous Cac was compromised by some uncertainty regarding the true distribution of Cac relative to Brp. The consensus view is that there is a high degree of correlation between Cac and Brp-defined active zones. This view is based on data collected from transgenic C-terminal GFP-tagged Cac (Akbergenova et al., Citation2017; Kawasaki et al., Citation2004; Rasse et al., Citation2005), genomic C-terminal GFP-tagged Cac (Matkovic et al., Citation2013), and the CRISPR/Cas9 edited cac locus which yields an N-terminal GFP-tagged Cac (CacsfGFP-N; Gratz et al., Citation2018). Cac-kisser-pHerry shows a strong correlation with Brp, suggesting that it effectively localizes with Cac ( and Figures S4–S6). However, the only antibody for Cac shows a mismatch with Brp (Chang et al., Citation2014), and this mismatch is also observed when PMCA-kisser-FAP is compared to CacsfGFP-N (Figure S7). The crowded environment of the synaptic cleft may decrease the staining efficiency for Cac-kisser-FAP, although this explanation seems strained as the HA-tag antibody is just as ineffective at finding its epitope in fixed and permeabilized tissue, as is the fluorogen in the living tissue (Figure S7), i.e. they both reveal the same incomplete labelling of CacsfGFP-N clusters. Ultimately, we favor the interpretation that all Brp-defined active zones harbor Cac, but Cac-kisser-pHerry is better able to locate all of the Cac clusters with its longer ‘linker’ (two fluorophores) between the TM domain and its C-terminus (Figure S1, and versus Figure S7A).
Although ultimately inconclusive in this example, the data presented in demonstrate the unique advantage kisser probes have over alternate approaches. Immunohistochemical detection of PMCA results in a combined pre- and post-synaptic readout of the endogenous location of PMCA, without any means to distinguish either side. Translational fusions with PMCA, expressed using either neuronal or muscle drivers, would enable differential tagging of pre- and post-synaptic pools of PMCA, but would involve a significantly higher level of active PMCA on one side of the synapse. Overexpression of PMCA would alter the kinetics of Ca2+/H+ exchange and most likely alter synapse physiology, and it would be desirable to disable the activity of overexpressed PMCA. Kisser probes skirt this issue by removing the extracellular and TM portions of PMCA. CRISPR/Cas9 gene editing might be used to insert a tag into the endogenous locus, thus avoiding overexpression issues, but it would not allow pre-synaptic PMCA to be distinguished from post-synaptic PMCA.
Kisser probes also allow for an increased targeting specificity for probes of physiological activity in protein-defined subdomains close to the PM. The experiments described herein deal with pH subdomains at the NMJ, but Ca2+ entry is also spatially heterogeneous and Ca2+-imaging approaches might benefit from the kisser approach by increasing the signal-to-noise ratio and enabling a detailed exploration of Ca2+ subdomains. Kisser probes might also be combined with reagents compatible with super-resolution microscopy techniques such as STORM and STED, to better resolve the subcellular localization of proteins. Such studies are contingent on the success of tagging endogenous proteins or immunological detection. Kisser probes provide a streamlined solution in the form of FAP-based kissers, as the Malachite Green fluorogen is compatible with STED (Fitzpatrick et al., Citation2009).
Overall, the kisser probes represent a practical contribution to the molecular biological toolkit. The approach described here should be applicable to any PM protein, but its success requires that intracellular tails mimicking target proteins be stripped of peptide sequences that might interfere with the localization of the probe to its target, or that cause unintended interactions between the probe and its target.
Conclusions
Kisser probes offer a simple but novel approach to resolving the distribution of endogenous membrane proteins in either live or fixed tissues. They are a fusion of an intracellular terminus mimicking the intracellular anchoring motifs of the target protein with a generic TM domain and a reporter tag. We demonstrated that (i) Cac-kisser and Sh-kisser probes recapitulate the localization of endogenous Cac and Dlg, respectively; (ii) PMCA-kisser can be used to investigate the relative distribution of pre-synaptically versus post-synaptically expressed PMCA at the Drosophila larval NMJ; and (iii) kisser probes can be used to live target functional probes (chemical and protein fluorescent pH reporters) to protein-defined subcellular domains of the PM.
Supplemental Material
Download PDF (2.7 MB)Acknowledgements
This article is dedicated to Harold Atwood, an inspirational, sage and patient mentor.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Akbergenova, Y., Zhang, Y.V., Weiss-Sharabi, S., Cunningham, K.L., & Littleton, J.T. (2017). Examining molecular determinants underlying heterogeneity of synaptic release probability using optical quantal imaging. bioRxiv, doi:10.1101/240549
- Benetti, F., Micetic, I., Carsughi, F., Spinozzi, F., Bubacco, L., & Beltramini, M. (2011). Insights into the oligomerization process of the C-terminal domain of human plasma membrane Ca(2)+-ATPase. Archives of Biochemistry and Biophysics, 506, 194–200. doi:10.1016/j.abb.2010.11.017
- Bolbat, A., & Schultz, C. (2017). Recent developments of genetically encoded optical sensors for cell biology. Biology of the Cell, 109, 1–23. doi:10.1111/boc.201600040
- Brown, M., Stafford, L.J., Onisk, D., Joaquim, T., Tobb, A., Goldman, L., … Chambers, R. (2013). Snorkel: An epitope tagging system for measuring the surface expression of membrane proteins. PLoS One, 8, e73255. doi:10.1371/journal.pone.0073255
- Chang, J.C., Hazelett, D.J., Stewart, J.A., & Morton, D.B. (2014). Motor neuron expression of the voltage-gated calcium channel cacophony restores locomotion defects in a Drosophila, TDP-43 loss of function model of ALS. Brain Research, 1584, 39–51. doi:10.1016/j.brainres.2013.11.019
- Costantini, L.M., Baloban, M., Markwardt, M.L., Rizzo, M., Guo, F., Verkhusha, V.V., & Snapp, E.L. (2015). A palette of fluorescent proteins optimized for diverse cellular environments. Nature Communications, 6, 7670. doi:10.1038/ncomms8670
- Crivat, G., & Taraska, J.W. (2012). Imaging proteins inside cells with fluorescent tags. Trends in Biotechnology, 30, 8–16. doi:10.1016/j.tibtech.2011.08.002
- Desai, S.A., & Lnenicka, G.A. (2011). Characterization of postsynaptic Ca2+ signals at the Drosophila larval NMJ. Journal of Neurophysiology, 106, 710–721. doi:10.1152/jn.00045.2011
- Di Leva, F., Domi, T., Fedrizzi, L., Lim, D., & Carafoli, E. (2008). The plasma membrane Ca2+ ATPase of animal cells: Structure, function and regulation. Archives of Biochemistry and Biophysics, 476, 65–74. doi:10.1016/j.abb.2008.02.026
- Falchetto, R., Vorherr, T., & Carafoli, E. (1992). The calmodulin-binding site of the plasma membrane Ca2+ pump interacts with the transduction domain of the enzyme. Protein Science, 1, 1613–1621. doi:10.1002/pro.5560011209
- Fitzpatrick, J.A.J., Yan, Q., Sieber, J.J., Dyba, M., Schwarz, U., Szent-Gyorgyi, C., … Bruchez, M.P. (2009). STED nanoscopy in living cells using fluorogen activating proteins. Bioconjugate Chemistry, 20, 1843–1847. doi:10.1021/bc900249e
- Fritze, C.E., & Anderson, T.R. (2000). Epitope tagging: General method for tracking recombinant proteins. Methods in Enzymology, 327, 3–16. doi:10.1016/S0076-6879(00)27263-7
- Fuentes-Medel, Y., Logan, M.A., Ashley, J., Ataman, B., Budnik, V., & Freeman, M.R. (2009). Glia and muscle sculpt neuromuscular arbors by engulfing destabilized synaptic boutons and shed presynaptic debris. PLOS Biology, 7, e1000184. doi:10.1371/journal.pbio.1000184
- Graf, E.R., Valakh, V., Wright, C.M., Wu, C., Liu, Z., Zhang, Y.Q., & DiAntonio, A. (2012). RIM promotes calcium channel accumulation at active zones of the Drosophila neuromuscular junction. Journal of Neuroscience, 32, 16586–16596. doi:10.1523/JNEUROSCI.0965-12.2012
- Gratz, S.J., Bruckner, J.J., Hernandez, R.X., Khateeb, K., Macleod, G., & O'Connor-Giles, K.M. (2018). Calcium channel levels at single synapses predict release probability and are upregulated in homeostatic potentiation. bioRxiv, doi:10.1101/240051
- Gronwald, R.G., Grant, F.J., Haldeman, B.A., Hart, C.E., O'Hara, P.J., Hagen, F.S., … Murray, M.J. (1988). Cloning and expression of a cDNA coding for the human platelet-derived growth factor receptor: evidence for more than one receptor class. Proceedings of the National Academy of Sciences of the United States of America, 85(10), 3435–3439. doi:10.1073/pnas.85.10.3435
- Guerrero, G., Reiff, D.F., Agarwal, G., Ball, R.W., Borst, A., Goodman, C.S., & Isacoff, E.Y. (2005). Heterogeneity in synaptic transmission along a Drosophila larval motor axon. Nature Neuroscience, 8, 1188–1196. doi:10.1038/nn1526
- Hibino, H., Pironkova, R., Onwumere, O., Vologodskaia, M., Hudspeth, A.J., & Lesage, F. (2002). RIM binding proteins (RBPs) couple Rab3-interacting molecules (RIMs) to voltage-gated Ca(2+) channels. Neuron, 34, 411–423. doi:10.1016/S0896-6273(02)00667-0
- Jing, C., & Cornish, V.W. (2011). Chemical tags for labeling proteins inside living cells. Accounts of Chemical Research, 44, 784–792. doi:10.1021/ar200099f
- Kaeser, P.S., Deng, L., Wang, Y., Dulubova, I., Liu, X., Rizo, J., & Südhof, T.C. (2011). RIM proteins tether Ca2+ channels to presynaptic active zones via a direct PDZ-domain interaction. Cell, 144, 282–295. doi:10.1016/j.cell.2010.12.029
- Kawasaki, F., Zou, B., Xu, X., & Ordway, R.W. (2004). Active zone localization of presynaptic calcium channels encoded by the cacophony locus of Drosophila. Journal of Neuroscience, 24, 282–285. doi:10.1523/JNEUROSCI.3553-03.2004
- Kittel, R.J., Wichmann, C., Rasse, T.M., Fouquet, W., Schmidt, M., Schmid, A., … Sigrist, S.J. (2006). Bruchpilot promotes active zone assembly, Ca2+ channel clustering, and vesicle release. Science, 312, 1051–1054. doi:10.1126/science.1126308
- Lahey, T., Gorczyca, M., Jia, X.X., & Budnik, V. (1994). The Drosophila tumor suppressor gene dlg is required for normal synaptic bouton structure. Neuron, 13, 823–835. doi:10.1016/0896-6273(94)90249-6
- Lnenicka, G.A., Grizzaffi, J., Lee, B., & Rumpal, N. (2006). Ca2+ dynamics along identified synaptic terminals in Drosophila larvae. Journal of Neuroscience, 26, 12283–12293. doi:10.1523/JNEUROSCI.2665-06.2006
- Lopreiato, R., Giacomello, M., & Carafoli, E. (2014). The plasma membrane calcium pump: New ways to look at an old enzyme. Journal of Biological Chemistry, 289, 10261–10268. doi:10.1074/jbc.O114.555565
- Matkovic, T., Siebert, M., Knoche, E., Depner, H., Mertel, S., Owald, D., … Sigrist, S.J. (2013). The Bruchpilot cytomatrix determines the size of the readily releasable pool of synaptic vesicles. Journal of Cell Biology, 202, 667–683. doi:10.1083/jcb.201301072
- Maue, R.A. (2007). Understanding ion channel biology using epitope tags: Progress, pitfalls, and promise. Journal of Cellular Physiology, 213, 618–625. doi:10.1002/jcp.21259
- McLachlan, E.M., & Martin, A.R. (1981). Non-linear summation of end-plate potentials in the frog and mouse. Journal of Physiology (London), 311, 307–324. doi:10.1113/jphysiol.1981.sp013586
- Menon, K.P., Carrillo, R.A., & Zinn, K. (2013). Development and plasticity of the Drosophila larval neuromuscular junction. Wiley Interdisciplinary Reviews-Developmental Biology, 2, 647–670. doi:10.1002/wdev.108
- Peled, E.S., & Isacoff, E.Y. (2011). Optical quantal analysis of synaptic transmission in wild-type and rab3-mutant Drosophila motor axons. Nature Neuroscience, 14, 519–526. doi:10.1038/nn.2767
- Rasse, T.M., Fouquet, W., Schmid, A., Kittel, R.J., Mertel, S., Sigrist, C.B., … Sigrist, S.J. (2005). Glutamate receptor dynamics organizing synapse formation in vivo. Nature Neuroscience, 8, 898–905. doi:10.1038/nn1484
- Rimessi, A., Coletto, L., Pinton, P., Rizzuto, R., Brini, M., & Carafoli, E. (2005). Inhibitory interaction of the 14-3-3{epsilon} protein with isoform 4 of the plasma membrane Ca(2+)-ATPase pump. Journal of Biological Chemistry, 280, 37195–37203. doi:10.1074/jbc.M504921200
- Rossano, A.J., Chouhan, A.K., & Macleod, G.T. (2013). Genetically encoded pH-indicators reveal activity-dependent cytosolic acidification of Drosophila motor nerve termini in vivo. Journal of Physiology, 591, 1691–1706. doi:10.1113/jphysiol.2012.248377
- Rossano, A.J., Kato, A., Minard, K.I., Romero, M.F., & Macleod, G.T. (2017). Na(+)/H(+) exchange via the Drosophila vesicular glutamate transporter mediates activity-induced acid efflux from presynaptic terminals. Journal of Physiology, 595, 805–824. doi:10.1113/JP273105
- Shastry, B.S. (1995). Overexpression of genes in health and sickness. A bird's eye view. Comparative Biochemistry and Physiology – Part B: Biochemistry & Molecular Biology, 112, 1–13. doi:10.1016/0305-0491(95)00055-D
- Szent-Gyorgyi, C., Schmidt, B.F., Creeger, Y., Fisher, G.W., Zakel, K.L., Adler, S., … Waggoner, A. (2008). Fluorogen-activating single-chain antibodies for imaging cell surface proteins. Nature Biotechnology, 26, 235–240. doi:10.1038/nbt1368
- Szent-Gyorgyi, C., Schmidt, B.F., Fitzpatrick, J.A., & Bruchez, M.P. (2010). Fluorogenic dendrons with multiple donor chromophores as bright genetically targeted and activated probes. Journal of the American Chemical Society, 132, 11103–11109. doi:10.1021/ja9099328
- Tejedor, F.J., Bokhari, A., Rogero, O., Gorczyca, M., Zhang, J., Kim, E., … Budnik, V. (1997). Essential role for dlg in synaptic clustering of Shaker K + channels in vivo. Journal of Neuroscience, 17, 152–159. doi:10.1523/JNEUROSCI.17-01-00152.1997
- Thomas, R.C. (2009). The plasma membrane calcium ATPase (PMCA) of neurones is electroneutral and exchanges 2 H + for each Ca2+ or Ba2+ ion extruded. Journal of Physiology, 587, 315–327. doi:10.1113/jphysiol.2008.162453
- von Heijne, G. (2007). The membrane protein universe: What's out there and why bother? Journal of Internal Medicine, 261, 543–557. doi:10.1111/j.1365-2796.2007.01792.x
- Wang, K.K., Roufogalis, B.D., & Kuo, T.H. (1991). Structure–function relationship of the human erythrocyte plasma membrane Ca(2+)-ATPase revealed by V8 protease treatment. Biochemical Journal, 279, 537–544. doi:10.1042/bj2790537
- Wang, S., Yang, J., Tsai, A., Kuca, T., Sanny, J., Lee, J., … Krieger, C. (2011). Drosophila adducin regulates Dlg phosphorylation and targeting of Dlg to the synapse and epithelial membrane. Developmental Biology, 357, 392–403. doi:10.1016/j.ydbio.2011.07.010
- Watschinger, K., Horak, S.B., Schulze, K., Obermair, G.J., Koschak, A., Sinnegger-Brauns, M.J., … Striessnig, J. (2008). Functional properties and modulation of extracellular epitope-tagged Ca(V)2.1 voltage-gated calcium channels. Channels (Austin), 2, 461–473. doi:10.4161/chan.2.6.6793
- Zhang, F., Gu, W., Hurles, M.E., & Lupski, J.R. (2009). Copy number variation in human health, disease, and evolution. Annual Review of Genomics and Human Genetics, 10, 451–481. doi:10.1146/annurev.genom.9.081307.164217
- Zito, K., Fetter, R.D., Goodman, C.S., & Isacoff, E.Y. (1997). Synaptic clustering of Fascilin II and Shaker: Essential targeting sequences and role of Dlg. Neuron, 19, 1007–1016. doi:10.1016/S0896-6273(00)80393-1