
Abstract
Background
Six Homo naledi early juveniles were recovered from U.W. 101 (Dinaledi Chamber), U.W. 102 (Lesedi Chamber), and U.W. 110 in the Rising Star cave system.
Aim
This paper develops the information for the H. naledi early juvenile life stage, as defined by a combination of deciduous and permanent dentition, and the eruption of the first permanent molar.
Subjects and methods
The growing number of young individuals recovered from the Rising Star cave system allows us to gain a better understanding of their variation, or lack thereof, and provides a basis to estimate broad ranges for age at death of the individuals. The individuals are identified and described through craniodental remains and spatial associations.
Results and conclusion
Our results show that the teeth are remarkably consistent across the localities in their metric and non-metric traits, and our analyses refine previous estimations on dental eruptions with the first permanent molar erupting first in the sequence among permanent teeth.
Introduction
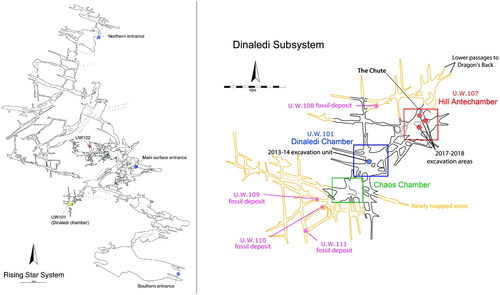
Homo naledi is known from the Rising Star cave system, South Africa, where its remains have been reported from three localities: U.W. 101 (Dinaledi Chamber), U.W. 102 (Lesedi Chamber), and U.W. 110 (Berger et al. Citation2015; Hawks et al. Citation2017; Brophy et al. Citation2021) (). Continued exploration of the cave system led to the discovery of additional localities that are current targets of investigation, U.W. 107–111 (see ) (Elliott et al. Citation2021). A geological date estimate for the Dinaledi Chamber hominin remains places them between 335,000 and 241,000 years ago (Dirks et al. Citation2017; Robbins et al. Citation2021). No estimate of age is yet available for the Lesedi Chamber or other areas (Hawks et al. Citation2017; Brophy et al. Citation2021). To date, this species is the best-documented later Middle Pleistocene hominin from Africa, and may represent a sister-species to our own (Dembo et al. Citation2016; Berger et al. Citation2017).
Figure 1. (A) Overview of the Rising Star cave System, showing the two main chamber locations of Homo naledi material; (B) Detail of the Dinaledi subsystem, showing the multiple fossil deposits (from Elliott et al. Citation2021).
Anatomically, Homo naledi is remarkable for its retention of a number of australopith features in a late-surviving hominin. These traits include a small brain and body size, laterally flared ilia, and slight curvature of the digits in the hands and feet (Berger et al. Citation2015; Harcourt-Smith et al. Citation2015; Kivell et al. Citation2015; Garvin et al. Citation2017; VanSickle et al. Citation2018). The species also shares derived features with later Middle and Late Pleistocene Homo, such as human-like dentition (tooth sizes, absence of accessory traits such as protostylids on the lower M2s, and lack of accessory cusps on mandibular permanent dentition and deciduous upper and lower m2), large spinal canal, wrist morphology, long, thin lower limbs, and human-like brain organisation (Berger et al. Citation2015; Feuerriegel et al. Citation2017; Hawks et al. Citation2017; Marchi et al. Citation2017; Williams et al. Citation2017; Holloway et al. Citation2018; Irish et al. Citation2018; Bailey et al. Citation2019). Homo naledi expresses several species-specific anatomies, such as robust, heavily muscled first metacarpals (Berger et al. Citation2015; Kivell et al. Citation2015). Multiple Homo naledi individuals have been recovered across all life stages, including infants, early juveniles, late juveniles, sub-adults, young and old adults (Berger et al. Citation2015; Dirks et al. Citation2015). Age classes for the Rising Star paleodeme are primarily defined based on dentition (Berger et al. Citation2015, Citation2023; Hawks et al. Citation2017; Bolter et al. Citation2020; Brophy et al. Citation2021). Preservation of this range of life stages within an assemblage is rare in the fossil hominin record (see also Bermúdez de Castro et al. Citation2004).
This paper provides additional evidence for one of the Homo naledi life stages: early juvenile (E-JUV). This life stage is defined by the presence of a combination of deciduous and permanent dentition, marked by the eruption of the first permanent molar (Bolter et al. Citation2018). Bolter et al. (Citation2018) distinguish between six age classes: infant, early juvenile, late juvenile, subadult, young adult and old adult (see same for full descriptions). Briefly, E-JUVs are considered older than infants due to the existence of erupted permanent molars, but younger than late juveniles due to the lack of erupted M2s.
Three E-JUV individuals were originally identified in the Dinaledi Chamber from the 2013–2014 field seasons through dental associations. Two other E-JUV individuals were subsequently identified from the Lesedi Chamber and U.W. 110 through craniodental remains and spatial associations (Hawks et al. Citation2017; Brophy et al. Citation2021; Elliott et al. Citation2021). Although an additional four individuals are represented in the en bloc feature recovered in 2017 from the Hill Antechamber, U.W. 107, none of these specimens have been identified as an early juvenile (Berger et al. under review). Here, we further assess spatial data from the Dinaledi Chamber, describe the early juveniles’ context of discovery within the Rising Star cave system, and discuss the distribution of these young individuals within the cave sub-system.
Materials and methods
Dental and associated cranial material in the early juvenile life/maturity stage recovered from sites U.W. 101, U.W. 102, and U.W. 110 were used in this study. Dental specimens were assessed based on wear, interproximal and occlusal facets, eruption/developmental ages, size, and spatial associations, when available. The eruption of teeth serves as biomarkers of life stages and is standardly used to estimate dental associations and age classes in fossil assemblages (e.g. Weidenreich Citation1937; Schultz Citation1940; Wolpoff Citation1979; Smith Citation1989; Bermúdez de Castro et al. Citation2004; Rosas et al. Citation2013; cf. Bolter and Cameron Citation2020). All individuals discussed in this paper fall into the early juvenile age class as they either have (1) associated first molars erupted, but no second molars yet erupted, or (2) other teeth recovered (e.g. deciduous canines) that are at wear stages consistent with the eruption of the first molars (after Bolter et al. Citation2018). Cranial remains were included with an individual if they were in the same developmental stage and unambiguously associated spatially (that is, not commingled with material that clearly represents other individuals). In this paper, we expand and modify the dental associations from Bolter et al. (Citation2018) to incorporate spatial data from the excavation records, and updated dental identifications. The excavation records, provenience, and photographic documentation from recovery are assessed to provide additional context and spatial association information. The excavation levels were divided into five depth levels: Surface (surface to 40 mm), Level 1 (41–70 mm), Level 2 (71–100 mm), Level 3 (101–130 mm), and Level 4 (131–160 mm) (Kruger Citation2017; Bolter et al. Citation2020).
Results
lists the hypothesised individuals. Below, each individual is defined by its elements and location.
Table 1. Early juvenile individuals from Rising Star localities U.W. 101, U.W. 102, and U.W. 110.
Associations
U.W. 101: Dinaledi Chamber
Early Juvenile #1
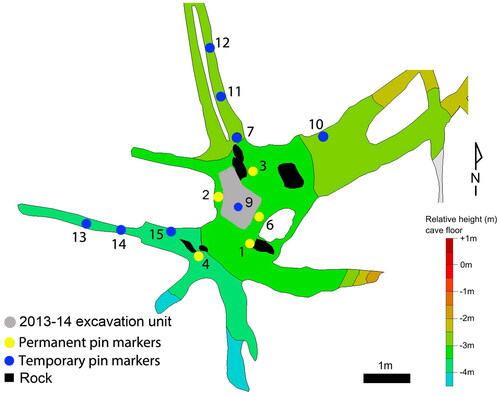
This juvenile is represented by six permanent molars: two (erupted) M1s and four (unerupted) M2s. The mandibular permanent teeth include right M1 (U.W. 101–285), left M1 (U.W. 101–582) (antimeres); and left M2 (U.W. 101–145) and right M2 (U.W. 101–507) (antimeres). The maxillary permanent teeth are right M2 (U.W. 101–593) and left M2 (U.W. 101–505) (antimeres). The erupted molars all have minimal or light wear. There are no deciduous molars present (, ). All specimens were found in the Surface level (surface to 40 mm): four at Pin 4, one at Pin 13, and one at Pin 2/9 ().
Figure 2. E-JUV individual #1. Top row (left to right): Left maxillary M2; right M2. Bottom row: mandibular left M2, M1; right M1, M2.
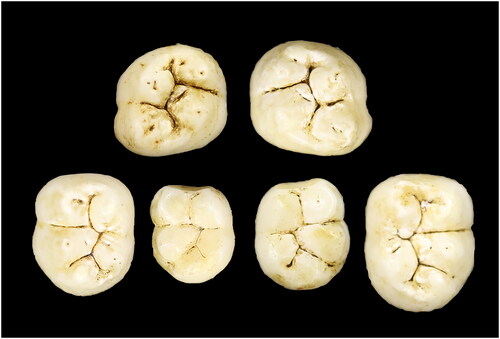
Figure 3. Dinaledi Chamber (U.W. 101), with excavation areas noted by pin numbers. Area 2/9 (shaded grey) represents the primary excavation unit for the 2013–2014 field seasons.
Early Juvenile #2
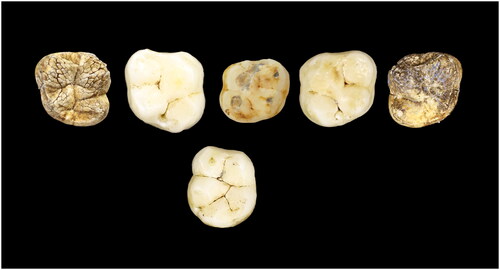
This juvenile is represented by five permanent molars and two permanent canines: three (erupted) M1s and two (unerupted) M2s. The permanent mandibular teeth are a left M1 (U.W. 101–814), right C1 (U.W. 101–886), and left C1 (U.W. 101–1126). The maxillary permanent teeth include a right M1 (U.W. 101–999) and left M1 (U.W. 101–445) (antimeres) (all have unworn or minimal wear); and the crowns of the right M2 (U.W. 101–1135) and left M2 (U.W. 101–1063) (antimeres) (, ). One deciduous tooth with moderate wear is also considered to be part of this individual: a right maxillary dm2 (U.W. 101–384). All of these specimens were recovered in U.W. 101 from the primary excavation unit, Pin 2 ( and ).
Figure 4. E-JUV individual #2. Top row (left to right): Left maxillary M2, M1; right dm2, M1, M2. Bottom row: mandibular left M1.
Early Juvenile #3
The designation of Juvenile #3 as E-JUV is based on a deciduous left canine with moderate wear, ldc U.W. 101–824 (Bolter et al. Citation2018) (). This canine is more worn than that recovered with the partial mandible U.W. 101–1400, which represents an individual of the Infant life history stage, with no erupted permanent dentition and relatively unworn first and second deciduous molars () (Berger et al. Citation2015). This specimen was recovered on the Surface level (surface to 40 mm) of 4SC ( and ).
Figure 5. Infant hemi-mandible U.W. 101–1400 (left). E-JUV individual #3 Ldc1 (right).

Figure 6. Pull-out section of Area 4, showing sub-divided surface collection areas.
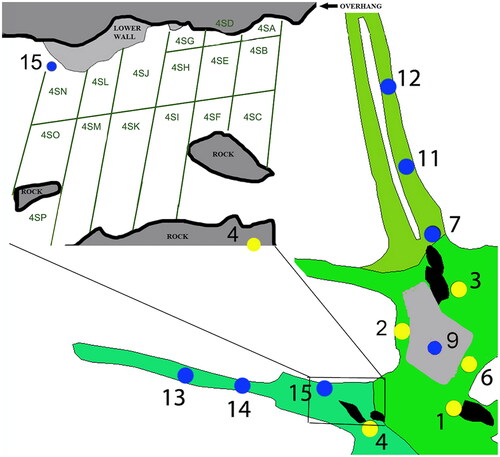
Early Juvenile #4
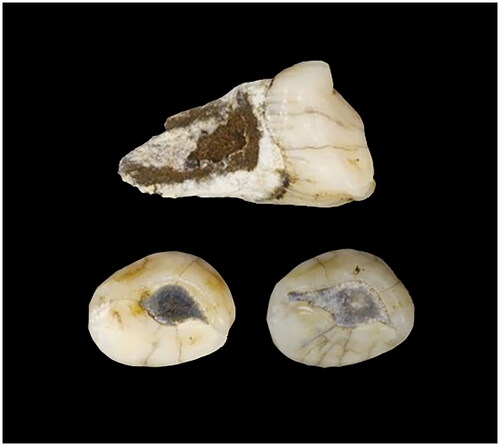
Juvenile #4 is designated as E-JUV based on an additional deciduous left canine, ldc1 U.W. 101–1571. Like U.W. 101–824, U.W. 101–1571 also has moderate wear (). This specimen was recovered from sediments collected in the Surface level (surface-40 mm), also from Area 4 (SB) ( and ).
Figure 7. E-JUV individual #4. U.W. 101–1571 (top) and occlusal of U.W. 101–1571 (left) and U.W. 101–824 (right).
U.W. 102: Lesedi Chamber
Early Juvenile #5
The early juvenile from Lesedi U.W. 102b is represented by five teeth, a partial right mandibular fragment with an immature tooth in the crypt, and cranial fragments (). The dental remains include one mandibular deciduous tooth with moderate wear, left dm2 (102b–437), three unerupted mandibular permanent teeth crowns, including a right P4 in crypt (102b–438), a left C (U.W. 102b–511), a left I2 (U.W. 102b–178), and two maxillary unerupted permanent teeth crowns: left I2 (U.W. 102b–515) and right P4 (U.W. 102b–503). The cranial fragments include: U.W. 102b–502, U.W. 102b–506, U.W. 102b–507, U.W. 102b–509, U.W. 102b–514, U.W. 102b–516. No growth remodelling has been noted on these fragments. The bone fragments are as follows: U.W. 102b–505, U.W. 102b–508, U.W. 102b–510, U.W. 102b–512, U.W. 102b–513.
Figure 8. E-JUV individual #5. From left to right (top): maxillary LI2, RP4; (bottom): mandibular LI2, LC, ldm2, partial mandible with RP4.
This individual is assigned as E-JUV by the presence of an alveolar hole for the developed roots of an erupted right M1, although the molar tooth itself was not recovered. All of the craniodental remains were recovered from Area 2 except for U.W. 102b–178. U.W. 102b–178 left second incisor tooth was found ex situ on the floor, in Area 4 ( and ). The elements were not deep in the sediment. No hominin remains were collected in Area 1, although an opening in the west wall connected this area to Area 2. No hominin remains were recovered in Area 3. The cranial material includes six cranial fragments, none of which are complete bone elements. All of the fragments are compatible in thickness and appearance with hominin cranial fragments though they are too small to allow a more definitive identification than “cranium” (Hawks et al. Citation2017).
Figure 9. Lesedi Chamber (U.W. 102) in the Rising Star cave.
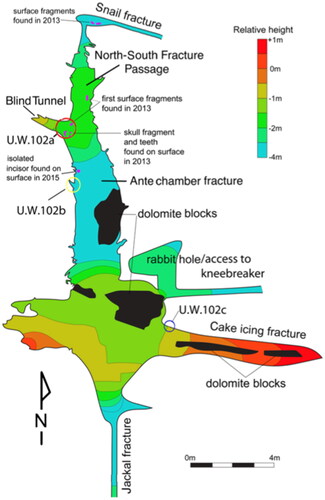
Figure 10. A (left). Schematic of the west wall region of the Lesedi Chamber, designated U.W. 102b showing Area 4, where U.W. 102b–178 was recovered and Area 2, where the remaining U.W. 102b hominin fossils were found. 10B (right). Image of west wall of Lesedi Chamber. White arrow points to where the U.W. 102b E-JUV hemi-mandible, teeth, and cranial fragments were recovered.
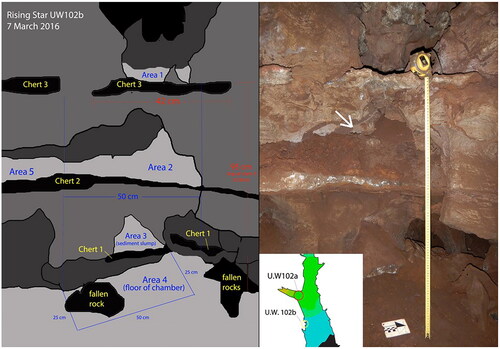
U.W. 110
Early Juvenile #6
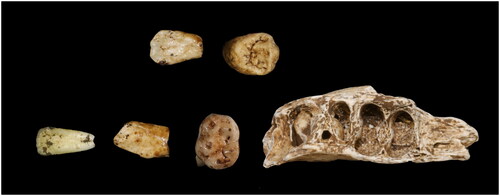
This early juvenile consists of 28 cranial fragments and six maxillary teeth (, ). The individual is assigned an early juvenile by the presence of one erupted, unworn permanent maxillary molar. All of the remains were found in a single, remote locality, at the southern edge of the Dinaledi subsystem, U.W. 110 ( and ). Two maxillary deciduous antimere teeth with moderate wear were recovered: right dm2 (U.W. 110–1) and left dm2 (U.W. 110–17). The maxillary permanent teeth include: right M1 crown (U.W. 110–5); right P4 crown germ (U.W. 110–2); right I1 crown (U.W. 110–14) and right I2 crown (U.W. 110–15). The frontal and cranial fragments are as follows: U.W. 110–3, U.W. 110–4, U.W. 110–6, U.W. 110–7, U.W. 110–8, U.W. 110–9, U.W. 110–10, U.W. 110–11, U.W. 110–12, U.W. 110–13, and U.W. 110–16.
Figure 11. From right to left: Ldm2, Rdm2, RI1, RI2, RP4, RM1.
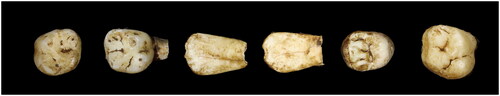
Figure 12. Sediment rise at back of passage U.W. 110 where elements of E-JUV #6 were recovered.

Depositional context
U.W. 101: Dinaledi Chamber
The context of the H. naledi remains in U.W. 101 has been extensively described (Berger et al. Citation2015; Dirks et al. Citation2015, Citation2016, Citation2017; Elliott et al. Citation2018, Citation2021; Tucker et al. Citation2018). Briefly, the E-JUV materials were part of a large assemblage that likely resulted from a complex depositional process (Dirks et al. Citation2015). The materials excavated during the 2013–2014 seasons include an MNI of 3 E-JUV, while an additional E-JUV individual is reported in this paper, bringing the current Dinaledi Chamber MNI to 21 individuals ().
Table 2. Homo naledi individuals and their life stages by the fossil site area within the Rising Star cave system.
U.W. 102: Lesedi Chamber
The U.W. 102 context has been explained in detail in Hawks et al. (Citation2017). The fossils from U.W. 102 are spatially separated into three sub-contexts: 102a, 102b, and 102c. U.W. 102b is approximately 3.8 m south and 1.8 m west of U.W. 102a (Hawks et al. Citation2017). The Juvenile #5 craniodental remains were found in U.W. 102b and represent an MNI of 1. Juvenile #5 is the only hominin recovered in U.W. 102b. Small faunal remains were also found, including those of several rodents, although their association with the hominin material is unknown. The first E-JUV specimen from U.W. 102 (LI2, U.W. 102b–178) was recovered in 2015, having been found ex situ on the surface floor leading into U.W. 102a area ( and ). Upon inspection, the specimen appeared to have tumbled down from sediments lying on the chert shelf above. In 2016, this chert shelf was re-examined and additional early juvenile material was identified. The area was then designated U.W. 102b and excavations of the chert shelf (designated Chert 2), as well as shelves above and below it (Cherts 1 and 3), commenced. All of the identifiable E-JUV material recovered after this point, including the cranial fragments, hemi-mandible and teeth, were recovered from sediments on Chert 2 (). The presence of maxillary and mandibular remains indicates a depositional event that included both the cranium and mandible.
A partial ilium consistent with the E-JUV developmental stage was recovered from the 102a locality, approximately 3 m from the 102b material (Cofran et al. Citation2022). Although this element represents the same developmental stage as the cranial and dental material from 102b, it is not clear whether they represent the same individual (Hawks et al. Citation2017). All other hominin material from 102a is attributable to an adult developmental stage and is interpreted as representing the LES1 individual (Hawks et al. Citation2017).
U.W. 110
Elliott et al. (Citation2021) and Brophy et al. (Citation2021) describe the U.W. 110 locality. Briefly, U.W. 110 is in a difficult-to-access fissure, approximately 13 m southwest of the 2013–2014 excavation area in the Dinaledi Chamber and, therefore, deeper into the subsystem. The fossil remains were collected from the surface of a short slope of sediment within the crevice (). The constricted nature of the space makes excavation very challenging, and none have been undertaken to date (Elliott et al. Citation2021). The cranial remains are highly fragmented and include discontiguous portions such as the maxillary teeth, a large supraorbital and interorbital fragment, sphenoid, frontal, and parietal fragments. The absence of intervening portions, and extreme state of fragmentation, both suggest that the skull was deposited here and subsequently fragmented with the loss of fragile elements such as the maxillary bones. No other individuals or non-hominin fauna are currently associated with E-JUV #6.
Discussion
Association refinements
In this paper, we expanded and modified the dental associations from Bolter et al. (Citation2018) which incorporates spatial data and updated dental identifications. One tooth (U.W. 101–020, M1) was disassociated from E-JUV #1 (Dinaledi Hominin 8), as it was recovered in what is now recognised as a separate fossil deposit, U.W. 107, the Hill Antechamber, adjacent to U.W. 101 (Elliott et al. Citation2021). Excavations here continue to uncover specimens, and we hypothesise this molar will associate with other material from this locality (Elliott et al. Citation2018, Peixotto et al. Citation2020; Tucker et al. Citation2018; Berger et al. under review). Dinaledi E-JUV #2 (Dinaledi Hominin 12) now includes U.W. 101–1063 M2 based on developmental congruity.
An additional E-JUV has been proposed based on a deciduous left canine (U.W. 101–1571), which is consistent in wear with another left deciduous canine already incorporated in the original MNI count from U.W. 101 (U.W. 101–824).
We assessed the Dinaledi Chamber collection for permanent canines that were relatively unworn, and for all deciduous canines, to locate any canines in the assemblage that were developmentally consistent with this early juvenile stage. Two permanent canines matched this description. U.W. 101–886 is a permanent, unworn right mandibular canine with approximately ¼ root formation. U.W. 101–1126 is a left permanent mandibular canine with no wear facet and incomplete root formation. We recognise these teeth as antimeres. U.W. 101–1126 was recovered from Pin 2, Level 1 and U.W. 886 from Pin 2 (Surface level). Given their spatial locations and developmental stage, we hypothesise that these canines associate with Juvenile #2.
Dental eruption schedule
The dental maturity of Homo naledi combines a mosaic of ancestral and derived conditions (Cofran and Walker Citation2017; Mahoney et al. Citation2024). The deciduous dental eruption schedule for Homo naledi is [di1–di2]–dm1–dc–dm2 (Cofran and Walker Citation2017; Bailey et al. Citation2019; this study), a human-like sequence. In chimpanzees, the deciduous canines typically emerge after the deciduous second molars (Kuykendall et al. Citation1992).
The permanent tooth eruption pattern for Homo naledi can be slightly refined from Cofran and Walker (Citation2017) to denote that the M1 erupts first in sequence: M1 [I1-I2] [P3-P4] [C-M2] M3. In the permanent dentition, Homo naledi premolars erupt in sequence before the second molars, a trait shared with Homo sapiens (Cofran and Walker Citation2017). M2 root formation at emergence is less developed, an ape-like trait (Cofran and Walker Citation2017). Canines erupt later than typical for modern humans, a Homo naledi trait shared with chimpanzees and early hominins (Cofran and Walker Citation2017). However, canine enamel growth is more human-like in Homo naledi (Mahoney et al. Citation2024).
Age estimates
Six E-JUV H. naledi individuals are represented thus far at Rising Star, classified by the eruption of their first permanent molars. These individuals also have erupted and worn deciduous teeth, including canines and second molars, along with unerupted incisor, canine and fourth premolar crowns. When considering the first permanent molar, one Homo naledi individual exhibits an enamel cusp formation time greater than the average time for modern humans (1187 days versus 1088 days) (Mahoney et al. Citation2024; see also Smith et al. Citation2015). Modern humans from sub-Saharan Africa generally erupt first molars between 5.0 and 6.3 years old (means combined for mandibular and maxillary teeth and for females and males) (Esan et al. Citation2018). In a more refined analysis of a population of South African Blacks, lower first molars erupted by 6.4 years (n = 639; mean = 6.1, SD = 0.3) and upper first molars erupted by 6.6 years (n = 639; mean = 6.3, SD = 0.3) (Esan et al. Citation2018). Comparatively, free-living Pan troglodytes chimpanzees erupt the first lower molars no later than 4.6 years old (Kelley et al. Citation2020). Given the totality of the mosaic nature of Homo naledi dental development and the human-like enamel formation timing for the first molar, we hypothesise a human-like timing of maturity and estimate the early juvenile Homo naledi are at least about 5.0 − 6.6 years of age.
The Infant H. naledi partial mandible U.W. 101–1400 () provides evidence for wear patterns among the canines and deciduous molars in the species. The first deciduous molars wear first (score higher for wear) compared to the deciduous canines, which also score higher for wear than the second deciduous molars (see also Molnar Citation1971; Bailey et al. Citation2019). This wear pattern conforms to the sequence of emergence of deciduous teeth: dm1–dc–dm2 (Cofran and Walker Citation2017). This wear comparison allows an age estimation for the early juveniles represented only by moderately worn deciduous canines, Juvenile #3 and #4 from the Dinaledi Chamber. These two individuals represent the younger members of this age class. Juveniles #2, #5, and #6 each have second deciduous molars with moderate wear, consistent with canines that would exhibit more than moderate wear (i.e. heavy wear). Juveniles #2, #5, and #6 are therefore developmentally older than Juveniles #3 and #4. Although Juvenile #1 does not presently have deciduous canines or molars associated with it, the wear of the first permanent mandibular molars are consistent with Juvenile #2, and we estimate these two to be of similar age. In sum, the early juveniles fall into two sub-age categories, with the younger #3 and #4 from the Dinaledi Chamber, and the older #1, #2, #5, and #6 early juveniles from Dinaledi, Lesedi, and the remote fissure U.W. 110 area ().
Table 3. Relative wear patterns of deciduous first molar, canine ,and second molar.
A third left deciduous canine, U.W. 101–595, was recovered on the surface of Area 4 (SJ). This maxillary canine has minimal wear, is younger than the other two left mandibular canines, and is developmentally consistent with a right deciduous first molar U.W. 101–823 with light wear also found in Area 4SC. These two deciduous teeth are developmentally consistent with an individual that has no permanent dentition erupted, which would be in the Infant age class (Bolter et al. Citation2018). We note that at least four immature individuals have been recovered from the surface of Area 4 in the Dinaledi Chamber: three early juveniles and one infant.
Dental attributes
Morphologically, the revised H. naledi E-JUV dental associations are consistent with each other, and with the other individuals across age classes in the assemblage. The mesiodistal (or labiolingual) crown dimensions of the teeth all fall within the range of the H. naledi dental sample (Hawks et al. Citation2017; Brophy et al. Citation2021; Delezene et al. Citation2023). Some of the shared characteristics include evincing Carabelli’s traits and dental chipping, and similar crown outline shape. These findings support homogeneity of the Rising Star cave dental assemblage as documented in other studies (Towle et al. Citation2017; Irish et al. Citation2018; Bailey et al. Citation2019; Brophy et al. Citation2021; Delezene et al. Citation2024).
Summary and conclusions
Six individuals of E-JUV life stage were recovered in the Rising Star cave system, and we hypothesise that they are at least 5.0 − 6.6 years old based on human standards. These individuals are all consistent in morphology with the H. naledi paleodeme as initially described (Berger et al. Citation2015; Dirks et al. Citation2015), and subsequent discoveries have reinforced the homogeneity of the dental assemblage (Hawks et al. Citation2017; Brophy et al. Citation2021). The refinement of dental eruptions, developmental stages and wear patterns of these early juveniles strengthen the foundation for associations of deciduous and permanent isolated teeth within the larger assemblage. The Homo naledi assemblage now represents at least 29 individuals: 21 from Dinaledi, at least three from Lesedi, one from U.W. 110, and at least four from the Hill Antechamber (), and excavations continue to uncover new fossil deposits within the subsystem. Future analyses are ongoing to reintegrate fossil remains in all six localities around U.W. 101 (U.W. 107–111), as well as the three regions of U.W. 102 (U.W. 102a–c).
The early juveniles presented here differ in their depositional context, with U.W. 101 being a multi-depositional event and U.W. 102b and U.W. 110 as separate, single events (although not necessarily primary depositions). Further investigation of the patterning of early juvenile remains may test the extent of additional evidence to support hominin agency and intentional disposal of individuals in the Rising Star cave system.
Acknowledgements
We would like to thank Bernhard Zipfel for access to the fossils at the University of the Witwatersrand.
Disclosure statement
The authors have no relevant conflicts of interest to disclose. This article was submitted to the Human Origins & Survival Section of Annals of Human Biology (AHB) for which one of the authors is the Section Editor. This Section Editor reclused themselves from handling this article and it was assigned to another AHB Editor for independent and anonymous peer-review. In no way was this author involved in the editorial process for this article.
Additional information
Funding
References
- Bailey SE, Brophy JK, Moggi-Cecchi J, Delezene LK. 2019. The deciduous dentition of Homo naledi: a comparative study. J Hum Evol. 136:1. doi: 10.1016/j.jhevol.2019.102655.
- Berger LR, Hawks J, de Ruiter DJ, Churchill SE, Schmid P, Delezene LK, Kivell TL, Garvin HM, Williams SA, DeSilva JM, et al. 2015. Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa. eLife. 4:e09560. doi: 10.7554/eLife.09560.
- Berger LR, Hawks J, Dirks PH, Elliott M, Roberts EM. 2017. Homo naledi and Pleistocene hominin evolution in subequatorial Africa. eLife. 6:e24234.
- Berger LR, Makhubela T, Molopyane K, Kruger A, Randolph-Quinney P, Elliott M, Peixotto B, Fuentes A, Tafforeau P, Beyrand V, et al. 2023. Evidence for deliberate burial of the dead by Homo naledi. eLife. 12:RP89106. doi: 10.7554/eLife.89106.1.
- Bermúdez de Castro JM, Martinón-Torres M, Lozano M, Sarmiento S, Muela A. 2004. Paleodemography of the Atapuerca: Sima De Los Huesos hominin sample: a revision and new approaches to the paleodemography of the European Middle Pleistocene population. Journal of Anthropological Research. 60(1):5–12. doi: 10.1086/jar.60.1.3631006.
- Bolter DR, Elliott MC, Hawks J, Berger LR. 2020. Immature remains and the first partial skeleton of a juvenile Homo naledi, a late Middle Pleistocene hominin from South Africa. PLoS One. 15(4):e0230440. doi: 10.1371/journal.pone.0230440.
- Bolter DR, Cameron N. 2020. Utilizing auxology to understand ontogeny of extinct hominins: a case study on Homo naledi. Am J Phys Anthropol. 173(2):368–380. doi: 10.1002/ajpa.24088.
- Bolter DR, Hawks J, Bogin B, Cameron N. 2018. Palaeodemographics of individuals in Dinaledi Chamber using dental remains. S Afr J Sci. 114(1/2):6. doi: 10.17159/sajs.2018/20170066.
- Brophy JK, Elliott MC, De Ruiter DJ, Bolter DR, Churchill SE, Walker CS, Hawks J, Berger LR. 2021. Immature hominin craniodental remains from a new locality in the Rising Star cave system, South Africa. PaleoAnthropology. 1:1–14. 10.48738/2021.iss1.64.
- Cofran Z, Garvin H, Radovčić D, Elliott M, Peixotto B, Chinamitira G, García-Martínez D, Williams SA, Hawks J, Berger L. In prep. Infant Homo naledi skeleton reveals the evolution of growth and development.
- Cofran Z, VanSickle C, Valenzuela R, García-Martínez D, Walker CS, Hawks J, Zipfel B, Williams SA, Berger LR. 2022. The immature Homo naledi ilium from the Lesedi Chamber, Rising Star Cave, South Africa. American Journal of Biological Anthropology. 179(1):3–17. doi: 10.1002/ajpa.24522.
- Cofran Z, Walker CS. 2017. Dental development in Homo naledi. Biol Lett. 13(8):20170339. doi: 10.1098/rsbl.2017.0339.
- Delezene LK, Skinner MW, Bailey S, Brophy JK, Elliott ME, Gurtov A, Irish JD, Moggi-Cecchi J, de Ruiter DJ, Hawks J, et al. 2023. Descriptive catalog of Homo naledi dental remains from the 2013 to 2015 excavations of the Dinaledi Chamber, site U.W. 101, within the Rising Star cave system, South Africa. J Hum Evol. 180:103372. doi: 10.1016/j.jhevol.2023.103372.
- Delezene LK, Scott JE, Irish JD, Villaseñor A, Skinner MM, Hawks J, Berger LR. 2024. Sex-biased sampling may influence Homo naledi tooth size variation. J Hum Evol. 187:103490. doi: 10.1016/j.jhevol.2023.103490.
- Dembo M, Radovčić D, Garvin HM, Laird MF, Schroeder L, Scott JE, Brophy J, Ackermann RR, Musiba CM, De Ruiter DJ, et al. 2016. The evolutionary relationships and age of Homo naledi: An assessment using dated Bayesian phylogenetic methods. J Hum Evol. 97:17–26. doi: 10.1016/j.jhevol.2016.04.008.
- Dirks PHGM, Berger LR, Hawks J, Randolph-Quinney PS, Backwell LR, Roberts EM. 2016. Comment on “Deliberate body disposal by hominins in the Dinaledi Chamber, Cradle of Humankind, South Africa?” [J. Hum. Evol. 96 (2016) 145–148]. J Hum Evol. 96:149–153.
- Dirks PHGM, Berger LR, Roberts EM, Kramers JD, Hawks J, Randolph-Quinney PS, Elliott M, Musiba CM, Churchill SE, de Ruiter DJ, et al. 2015. Geological and taphonomic context for the new hominin species Homo naledi from the Dinaledi Chamber, South Africa. eLife. 4:e09561. doi: 10.7554/eLife.09561.
- Dirks PHGM, Roberts EM, Hilbert-Wolf H, Kramers JD, Hawks J, Dosseto A, Duval M, Elliott M, Evans M, Grün R, et al. 2017. The age of Homo naledi and associated sediments in the Rising Star Cave, South Africa. eLife. 6:e24231. doi: 10.7554/eLife.24231.
- Elliott MC, Makhubela TV, Brophy JK, Churchill SE, Peixotto B, Feuerriegel EM, Morris H, Van Rooyen D, Ramalepa M, Tsikoane M, Kruger A, Spandler C, Kramers J, Roberts E, Dirks PHGM, Hawks J, Berger LR. 2021. Expanded explorations of the Dinaledi Subsystem, Rising Star Cave System, South Africa. Palaeoanthropology. 1:15−22. 10.48738/2021.iss1.68.
- Elliott MC, Peixotto B, Morris H, Feuerriegel EM, Tucker S, Hunter R, Ramalepa M, Tsikoane M, Roberts EM, Spandler C, et al. 2018. Hominin material recovered from the base of the chute in the hill antechamber, in the Dinaledi Chamber System of the Rising Star Cave. Am J Phys Anthropol. 165:76–76.
- Esan TA, Mothupi KA, Schepartz LA. 2018. Permanent tooth emergence: timing and sequence in a sample of Black Southern African children. Am J Phys Anthropol. 167(4):827–839. doi: 10.1002/ajpa.23714.
- Feuerriegel EM, Green DJ, Walker CS, Schmid P, Hawks J, Berger LR, Churchill SE. 2017. The upper limb of Homo naledi. J Hum Evol. 104:155–173. doi: 10.1016/j.jhevol.2016.09.013.
- Garvin HM, Elliott MC, Delezene LK, Hawks J, Churchill SE, Berger LR, Holliday TW. 2017. Body size, brain size, and sexual dimorphism in Homo naledi from the Dinaledi Chamber. J Hum Evol. 111:119–138. doi: 10.1016/j.jhevol.2017.06.010.
- Harcourt-Smith WE, Throckmorton Z, Congdon KA, Zipfel B, Deane AS, Drapeau MS, Churchill SE, Berger LR, DeSilva JM. 2015. The foot of Homo naledi. Nat Commun. 6(1):8432. doi: 10.1038/ncomms9432.
- Hawks J, Elliott M, Schmid P, Churchill SE, de Ruiter DJ, Roberts EM, Hilbert-Wolf H, Garvin HM, Williams SA, Delezene LK, et al. 2017. New fossil remains of Homo naledi from the Lesedi Chamber, South Africa. eLife. 6:e24232. doi: 10.7554/eLife.24232.
- Holloway RL, Hurst SD, Garvin HM, Schoenemann PT, Vanti WB, Berger LR, Hawks J. 2018. Endocast morphology of Homo naledi from the Dinaledi chamber, South Africa. Proc Natl Acad Sci USA. 115(22):5738–5743. doi: 10.1073/pnas.1720842115.
- Irish JD, Bailey SE, Guatelli-Steinberg D, Delezene LK, Berger LR. 2018. Ancient teeth, phenetic affinities, and African hominins: another look at where Homo naledi fits in. J Hum Evol. 122:108–123. doi: 10.1016/j.jhevol.2018.05.007.
- Kelley J, Schwartz GT, Smith TM. 2020. Age at first molar emergence in Pan troglodytes verus and variation in the timing of molar emergence among free-living chimpanzees. J Hum Evol. 145:102823. doi: 10.1016/j.jhevol.2020.102823.
- Kivell TL, Deane AS, Tocheri MW, Orr CM, Schmid P, Hawks J, Berger LR, Churchill SE. 2015. The hand of Homo naledi. Nat Commun. 6(1):8431. doi: 10.1038/ncomms9431.
- Kruger A. 2017. Site formation processes at Rising Star: taphonomy and 3D spatial analyses of the Homo naledi assemblage [PhD thesis]. University of Witwatersrand, South Africa.
- Kuykendall KL, Mahoney CJ, Conroy GC. 1992. Probit and survival analysis of tooth emergence ages in a mixed‐longitudinal sample of chimpanzees (Pan troglodytes). Am J Phys Anthropol. 89(3):379–399. doi: 10.1002/ajpa.1330890310.
- Laird MF, Schroeder L, Garvin HM, Scott JE, Dembo M, Radovčić D, Musiba CM, Ackermann RR, Schmid P, Hawks J, et al. 2017. The skull of Homo naledi. J Hum Evol. 104:100–123. doi: 10.1016/j.jhevol.2016.09.009.
- Mahoney P, McFarlane G, Taurozzi AJ, Madupe PP, O’Hara MC, Molopyane K, Cappellini E, Hawks J, Skinner M, Berger L. 2024. Human‐like enamel growth in Homo naledi. Am J Biol Anthropol. 2024:1–14. doi: 10.1002/ajpa.24893.
- Marchi D, Walker CS, Wei P, Holliday TW, Churchill SE, Berger LR, DeSilva JM. 2017. The thigh and leg of Homo naledi. J Hum Evol. 104:174–204. doi: 10.1016/j.jhevol.2016.09.005.
- Molnar S. 1971. Human tooth wear, tooth function and cultural variability. Am J Phys Anthropol. 34(2):175–189. doi: 10.1002/ajpa.1330340204.
- Peixotto B, Elliott MC, Hawks J, Berger L. 2020. A closer look at the spatial distribution of hominin fossils at Pin 8, Hill Antechamber, Rising Star Cave. Am J Phys Anthropol Supp. 171:213.
- Robbins JL, Dirks PH, Roberts EM, Kramers JD, Makhubela TV, Hilbert-Wolf HL, Elliott M, Wiersma JP, Placzek CJ, Evans M, et al. 2021. Providing context to the Homo naledi fossils: constraints from flowstones on the age of sediment deposits in rising Star Cave, South Africa. Chem Geol. 567:120108. doi: 10.1016/j.chemgeo.2021.120108.
- Rosas A, Estalrrich A, García-Vargas S, García-Tabernero A, Huguet R, Lalueza-Fox C, de la Rasilla M. 2013. Identification of Neandertal individuals in fragmentary fossil assemblages by means of tooth associations: the case of El Sidrón (Asturias, Spain). CR Palevol . 12(5):279–291. doi: 10.1016/j.crpv.2013.06.003.
- Schultz A. 1940. Growth and development of the chimpanzee. Contr. Embryol. 28:1–63.
- Smith BH. 1989. Dental development as a measure of life history in primates. Evolution. 43(3):683–688. doi: 10.2307/2409073.
- Smith TM, Tafforeau P, Le Cabec A, Bonnin A, Houssaye A, Pouech J, Moggi-Cecchi J, Manthi F, Ward C, Makaremi M, et al. 2015. Dental ontogeny in Pliocene and early Pleistocene hominins. PLoS One. 10(2):e0118118. doi: 10.1371/journal.pone.0118118.
- Towle I, Irish JD, De Groote I. 2017. Behavioral inferences from the high levels of dental chipping in Homo naledi. Am J Phys Anthropol. 164(1):184–192. doi: 10.1002/ajpa.23250.
- Tucker S, Kruger A, Elliott M, Peixotto B, Van Rooyen D, Tsikoane M, Ramalepa M, Hunter R, Roberts E, Dirks P. 2018. 3D Mapping of the Hominin-bearing deposits and associated passages and chambers of the Rising Star Cave System, South Africa. Am J Phys Anthropol Supp. 165:278.
- VanSickle C, Cofran Z, García-Martínez D, Williams SA, Churchill SE, Berger LR, Hawks J. 2018. Homo naledi pelvic remains from the Dinaledi Chamber, South Africa. J Hum Evol. 125:122–136. doi: 10.1016/j.jhevol.2017.10.001.
- Weidenreich F. 1937. The dentition of Sinanthropus pekinensis: a comparative odontography of the hominids. Natl Geol Survey China, Pall Sinica. 1:143–146.
- Williams SA, García-Martínez D, Bastir M, Meyer MR, Nalla S, Hawks J, Schmid P, Churchill SE, Berger LR. 2017. The vertebrae and ribs of Homo naledi. J Hum Evol. 104:136–154. doi: 10.1016/j.jhevol.2016.11.003.
- Wolpoff M. 1979. The Krapina dental remains. Am J Phys Anthropol. 50(1):67–114. doi: 10.1002/ajpa.1330500110.