
Abstract
We enrolled healthy subjects that received 2 to 4 injections of mRNA-based vaccination to prevent COVID-19 months to a year from the last vaccine boost, and we found numerous SARS-CoV-2 spike-specific regulatory T cell (Treg) that developed T cell memory as effector memory T cells (TEM) and central memory T cells (TCM). CD4+ CD25high Treg expressed the chemokine receptor CCR6 in a considerable percentage, suggesting T cell homing to the vascular endothelium, lung and gut epithelial cells and brain. Treg phenotype was different than peripherally-induced Treg (pTreg) that revert from pro-inflammatory T cells under repeated stimulatory conditions, suggesting that SARS-CoV-2 spike-specific Treg differentiated from naïve T cells in tissues where the SARS-CoV-2 spike proteins were synthetized. Twenty two of 22 subjects studied responded to vaccination developing a spike-specific CD4+ T helper (Th)1 response, and 20 of 22 developing a spike-specific CD8+ cytotoxic T cells (CTL) response. However, in vaccine recipients the expansion of spike-specific pro-inflammatory T cells was less significant than the expansion of spike-specific Treg. Effector (TEM) and central memory (TCM) Treg were numerous as early as after two vaccine doses, with no significant differences following additional vaccine boosts. In co-culture experiments under stimulatory conditions, Treg regulated naïve T cell differentiation toward a pro-inflammatory phenotype and suppressed interferon (IFN)γ production by SARS-CoV-2-specific CD4 + Th1 cells.
Introduction
In the past few years immense research effort has been made to understand the innate and adaptive immune response activated during severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection, and to identify the immune effector mechanisms of severe and lethal COVID-19 [Citation1].
In viral infections, innate immune cells recognize the virus and trigger a type I interferon (IFN) production within few hours of infection. The innate immune response restricts the viral replication and changes the homing of the inflammatory process by recruiting other effector innate immune cells, and importantly, prime B cells and T cells. The SARS-CoV-2 coronavirus avoids or delay intracellular innate responses associated with type I and type III IFN [Citation2,Citation3]. Most recently has been shown that modulation of type I interferon responses potently inhibits SARS-CoV-2 replication and inflammation in rhesus macaques [Citation4], suggesting that the adaptive immunity plays a critical importance in determining the outcome of the infection.
A peculiarity of the T cell response to SARS-CoV-2 is that CD4+ T helper 1 (Th1) responses are more prominent than CD8+ T cell (CTL) responses and that CD4+ Th1 maybe the primary line of adaptive anti-viral defense in SARS-CoV-2 infection (rev. in [Citation5]). We confirm these results in the current study.
The characterization of humoral antibody (Ab) responses and T cell -mediated immunity in COVID-19 has been pursued in many laboratories across the world allowing the generation of vaccines in record time. Of particular interest the T cell responses against SARS-CoV-2 spike protein, contained in almost all vaccine formulations [Citation6]. The humoral and cellular immune response and its memory development after vaccination with Moderna mRNA 1273, Pfizer/BioNTechBNT162b2, Jansen Ad26.COV2.S and Novavax NVX-CoV2373 has been well described [Citation7]. The peculiar aspects of the T cell memory repertoire: where CD8+ CTL were terminally differentiated effector (TMRA) and CD4 + Th1 effector (TEM) and central (TCM) have been discussed [Citation8], with data comparable in many laboratories.
Spike-specific regulatory T cells (Treg) seems very important in determining the severity of the disease [Citation9] and their expansion has been considered a possible tool to mitigate disease severity [Citation10]. Our laboratory determined the expansion of SARS-CoV-2 spike-specific Treg in pregnant women convalescent from COVID-19 [Citation11], suggesting protection via immune regulation of the mother/fetus interface. Treg have multiple functions, including the control of the milieu where the antigen presentation occurs, the polarization of innate cell lineages, limiting the expansion of autoreactive T cells and pro-inflammatory T cells that recognize peptides derived from autoantigens or pathogens [Citation12,Citation13].
Here we are showing that, following mRNA-based vaccination to protect from COVID-19, SARS-CoV-2 spike-specific-Treg precursors were very numerous, fully expanded after two vaccine injections and detectable in circulations several months after vaccination, with little changes following multiple vaccine boosts. Treg developed T cell memory, and expressed CCR6, a chemokine receptor that suggests homing and protection from exuberant inflammation of the vascular endothelium, lung and gut epithelium. CCR6 expression also allows spike-specific Treg to pass through the blood-brain barrier, becoming capable to directly regulate inflammation in the brain and cross-talk with neurons (reviewed in [Citation14]).
Materials and methods
Study population
We enrolled 22 healthy donors, 16 females and 6 males, aged 21 to 70, of different races, that received 2 to 4 vaccine administration within months to a year from blood drawing (UCSD IRB # 101213X).
Blood samples were collected from June 2022 to August 2023. Four subjects reported COVID-19 infection either before or in between vaccine boosts (). mRNA-based vaccines included bivalent formulations of BNT162b2 (Pfizer-BioNTech) and mRNA-1273 (Moderna); these vaccines are based on mRNA encoding the spike protein from the original (ancestral) strain of SARS-CoV-2 and from the B.1.1.529 (Omicron) variants BA.4 and BA.5 (BA.4/BA.5).
Table 1. Characteristics of the healthy donors enrolled in the study.
SARS-CoV-2 spike peptides mega pool
SARS-CoV-2 spike peptide mega pool were used to study the CD4+ and CD8+ T cell responses to SARS-CoV-2 in 22 vaccinated healthy subjects. The peptide mega pools were designed based on the reference genomic sequence of Wuhan-Hu-1 SARS-CoV-2 isolate (GenBank ID:MN908947), as described and validated in acute and convalescent SARS-COV-2 infected patients, as well as unexposed healthy subjects [Citation15,Citation16]. The SARS-CoV-2 spike mega pool contains 253 peptides, 15-amino acid-long, overlapping by 10 amino acids and spanning the entire spike protein. Peptides were synthesized as crude material (T.C. laboratories, San Diego), resuspended in DMSO, pooled according to mega pool design, and re-lyophilized.
AIM assay
Peripheral blood mononuclear cells (PBMC) were separated by Ficoll Hypaque density centrifugation. After count, 1x106 cells were stimulated in 96-well U bottom plates with 1 μg/ml of spike peptide mega pool cultured with 0.1% DMSO, the same concentration of DMSO (solvent) used in the mega pool-stimulated cultures, diluted in complete media served as unstimulated controls. Anti-CD3, (clone UCHT1, mouse IgG1κ, BD Bioscience) and anti-CD28 (clone CD28.2, mouse IgG1κ, BD Biosciences) stimulated PBMC served as a positive control for T cell activation in these experiments.
24h later, cell cultures were harvested and stained with monoclonal antibodies and analyzed by flow cytometry to study T cell activation, CCR6 expression, effector and memory phenotypes: anti-CD4-BV605 (clone RPA-T4, mouse IgG1κ, BD Bioscience), anti-CD8-PE/CF594 (clone RPA-T8, mouse IgG1k, BD Bioscience), anti-4-1BB-allophycocyanin (clone 4B4-1, mouse IgG1κ, Biolegend), anti-OX40-PE/Cy7 (clone Ber-ACT35, mouse IgG1κ, Biolegend), anti-CD69-PE (clone FN50, mouse IgG1κ, BD Bioscience), anti-CCR6-PerCp/Cy5.5 (clone 11A9, mouse IgG1κ, BD Bioscience), anti-CD45RA-APC-H7 (clone HI100, mouse IgG2bk, BD Bioscience), and anti-CCR7-FITC (clone G043H7, mouse IgG2aκ, Biolegend). Data were recorded on BD Canto II) and analyzed with FlowJo software version 10 (Tree Star). Isotype controls for each antibody were tested and showed no staining. Treg were defined by the co-expression of CD4 (anti-CD4-BV605, clone RPA-T4, mouse IgG1κ, BD Bioscience) and CD25high, mean fluorescence approximately 104 (anti-CD25-BV421 (clone M-A251, mouse IgG1κ, BD Biosciences).
Antigen-specific responses were determined by the expression of T cell activation-induced cell markers (AIM) assay by measuring the co-expression of 4-1BB and OX40, two TNF family member co-stimulatory molecules up-regulated following T cell receptor signaling on CD4+ T cells, and by measuring the co-expression of 4-1BB and CD69 (adhesion molecule involved in lymphocyte homing and trafficking) on CD8+ T cells [Citation16]. Many co-stimulatory molecules and selectins are shared between T cell lineages, especially CD4+ T cells, but the markers and the timing chosen define CD4+ Th1 and CD8+ CTL [Citation17]. The expression of the chemokine receptor CCR6 on AIM + CD4+ and CD8+ T cells was also analyzed. TEMRA, CD45RA + CCR7-, TEM, CD45RA − CCR7−, and TCM, CD45RA − CCR7+ were enumerated on AIM + CD4+ and CD8+ T cells.
Statistical analysis
Data were analyzed using Prism software version 9.0 (GraphPad Software). To compare the percentage of AIM + T cells in the unstimulated control and peptide stimulation, data obtained from the SARS-CoV-2 spike peptide megapool-stimulated culture and unstimulated controls in the individual cohort were tested using non-parametric paired tests. A P value ≤ 0.05 was considered significant.
Phenotype of SARS-CoV-2 spike-specific Treg
The generation and phenotypical characterization of spike-specific Treg from PBMC cultures or after FACS sorting from PBMC stimulated cultures was determined by: surface anti-CD4-PerCp/Cy5.5 (clone RPA-T4, mouse IgG1κ, eBioscience), anti-CD25-BV421 (clone M-A251, mouse IgG1κ, BD Biosciences), anti-IL-7Rα-FITC (clone eBioRDR5, mouse IgG1κ, eBioscience), anti-PD-1-BV605 (clone EH12.2H7, mouse IgG1κ, BioLegend), anti-CTLA-4-PE/Dazzle594 (clone BNI3, mouse IgG2aκ, BioLegend), and intracellular anti-FOXP3-PE (clone 259D, mouse IgG1κ, BioLegend). Data were recorded on BD Canto II (BD Bioscience) and analyzed with FlowJo software version 10 (Tree Star).
To obtain SARS-CoV-2 spike-specific T cell lines from PBMC, we stimulated 1×106 PBMC/well with SARS-CoV-2 spike epitopes in duplicate to assess Treg specificity by flow cytometry 24 h after cultures, and to expand in vitro SARS-CoV-2 spike-specific Treg precursors. Briefly, 72 h after peptide stimulation, cell preparation were harvested from the plate, washed, counted, and diluted to obtain 10×104 cells/well prior to restimulation with peptides and autologous irradiated PBMC as source of antigen presenting cells (APC). APC and Treg in the absence of peptides served as unstimulated control.
To address the expression of CTLA-4 and PD-1 on SARS-CoV-2 spike specific Treg depending upon IL-2 regimen, we stimulated PBMC for 72 h with SARS-CoV-2 spike peptides, tested the specificity in duplicate cultures, and FACS-sorted CD4+ CD25high Treg to obtain a pure population. After sorting, SARS-CoV-2 spike-specific Treg were restimulated with agonistic antibodies to CD3 and CD28 in the absence of IL-2, or with media conditioned with different amounts of IL-2, then studied for FOXP3, CTLA-4 and PD-1 expression.
FACS sorting of spike-specific Treg, spike-specific CD4+ Th1 and naïve T cells for in vitro suppression assays
Treg, AIM + CD4 + Th1 cells and naïve T cells have been FACS-sorted for functional experiments in co-cultures. Briefly, PBMC were separated and stimulated with the spike peptide mega pool in 24 well plates at the cell concentration of 4×106/ml. 72 h after cultures, cells were collected and the three T cell populations were stained with a combination of monoclonal antibodies to define: Treg (anti-CD4-BV605, clone RPA-T4, mouse IgG1κ, BD Biosciences and anti-CD25-BV421, clone M-A251, mouse IgG1κ, BD Biosciences), naïve T cells (anti-CD45RA-APC-H7, clone HI100, mouse IgG2bκ, BD Biosciences), CD4+ Th1 cells (anti-OX40-PE/Cy7, clone Ber-ACT35, mouse IgG1κ, BioLegend and anti-4-1BB-APC, clone 4B4-1, mouse IgG1κ, BioLegend). T cell populations were sorted with BD Aria II. Co-cultures were established at the cell ratio of 1:1 and stimulated with agonistic antibodies to CD3, (clone UCHT1, mouse IgG1κ, BD Bioscience) and CD28 (clone CD28.2, mouse IgG1κ, BD Biosciences). The down-regulation of IFNγ, measured by ELISA in culture supernatants, 24 h after Treg/naïve and Treg/AIM + co-cultures, served as a read out in these experiments.
Results
SARS-CoV-2 spike-specific Treg expand and develop memory together with spike-specific CD4+ Th1 and CD8+ CTL in vaccinated subjects
We enrolled 22 healthy subjects (aged 20 to 70, 16 females and 6 males of different ethnicity, ) that received two to four injections of mRNA-based SARS-CoV-2 vaccination (Pfizer or Moderna). Subjects were studied three months to a year from blood drawing, to define their SARS-CoV-2 spike-specific CD4+ Th1, CD8+ CTL and Treg repertoire.
The spike peptides were pooled and the design was carried out on the Wuhan-Hu-1 reference isolated (GenBank ID:MN908947). To ensure a comprehensive assessment of CD4-spike- and CD8-spike- specific reactivity, the main target of vaccine candidates, overlapping 15-mers by 10 spanning the entire protein have been synthetized and pooled. SARS-CoV-2 CD4 and CD8 peptides used in this study have been validated in acute and convalescent samples [Citation15,Citation16,Citation18,Citation19]; n = 253.
PBMC were isolated from heparin-treated blood by Ficoll hypaque, stimulated in vitro 24 h with a pool of SARS-CoV-2 spike peptides, to determine T cell activation by flow cytometry with the AIM assay.
CD4+ cell responses were evaluated by measuring the expression of two co-stimulatory molecules, OX-40 and 4-1BB, CD8+ T cell responses were evaluated by measuring the expression of 4-1BB and CD69, and Treg by measuring the expression of CD4+ CD25high. Co-stimulatory molecules and selectins are in common between many activated T cell lineages, however, careful studies in optimizing the AIM assay suggested that the markers and timing chosen in this study to define CD4+ Th1 and Treg are appropriate [Citation17]. Unstimulated PBMC served as negative control and PBMC stimulated with anti-CD3/anti-CD28 served as a positive control for T cell activation in these experiments.
T cell memory populations were defined as terminally differentiated effector memory (TEMRA, CD45RA + CCR7-), effector memory (TEM, CD45RA- CCR7-), and central memory (TCM, CD45RA- CCR7+).
CD4+ Th1 cells from all the 22 subjects responded to the SARS-CoV-2 spike peptides. Twenty of 22 subjects showed a CD8+ CTL response, and 2 subjects, #10 and #19, did not show expansion of spike-specific CD8+ cytotoxic T cells (, panel A). When we enumerated spike-specific CD4+ CD25high Treg, we found this population numerous in circulation in all vaccine recipients, independently from the number of vaccine boosts or the timing of the blood drawing from vaccination (, panel A and Supplementary Figure 1). COVID-19, contracted by 4 of 22 donors () did not have an impact in the expansion of spike-specific Treg.
Figure 1. Enumeration of SARS-CoV-2 spike-specific CD4+ Th1, CD8+ CTL and CD4+ CD25high Treg and their memory phenotype in 22 healthy vaccinated subjects. (A) PBMC were separated from 22 vaccinated subjects and stimulated in vitro with the SARS-CoV-2 spike peptides pool. Twenty-four hours after stimulation cells were collected and stained with specific monoclonal antibodies to determine their activation state (AIM assay). First gating on CD3+ T cells (anti-CD3-AF700, clone OKT3, mouse IgG2aκ, BioLegend), CD4+ Th1 were studied with anti-CD4-BV605 (clone RPA-T4, mouse IgG1κ, BD Biosciences), anti-4-1BB-allophycocyanin (clone 4B4-1, mouse IgG1κ, BioLegend), and anti-OX40-PE/Cy7 (clone Ber-ACT35, mouse IgG1κ, BioLegend). The percentage of AIM + CD4+ Th1 cells from un-stimulated control and SARS-CoV-2 CD4 S-all-stimulated cells were compared by the Wilcoxon matched-pairs signed rank test. CD8+ CTL were studied with anti-CD8-PE/CF594 (clone RPA-T8, mouse IgG1κ, BD Biosciences), anti-4-1BB-allophycocyanin (clone 4B4-1, mouse IgG1κ, BioLegend), and anti-CD69-PE (clone FN50, mouse IgG1κ, BD Biosciences). The percentage of AIM + CD8+ CTL from unstimulated control and CD4+ Th1 SARS-CoV-2 spike stimulated cells were compared by the Wilcoxon matched-pairs signed rank test. Regulatory T cells (Treg) were defined as CD4+ CD25high (anti-CD4-BV605, clone RPA-T4, mouse IgG1κ, BD Biosciences, and anti-CD25-BV421, clone M-A251, mouse IgG1κ, BD Biosciences). The percentage of CD4+ CD25high Treg from unstimulated control and SARS-CoV-2 spike-stimulated cells were compared by the Wilcoxon matched- pairs signed rank test. CCR6 expression on CD4 + Th1, CD8 + CTL and Treg was determined by staining with anti-CCR6-PerCp/Cy5.5 (clone 11A9, mouse IgG1κ, BD Biosciences). CCR6 expression was found in a large percentage of SARS-CoV-2-spike-specific CD4+ Th1 cells (median: 34.35%) and CD4+ CD25high Treg (median: 27.50%) in most subjects, unless CD8+ CTL where CCR6 was found in a much lower percentage (median: 11.66%). (B) T cell Memory phenotypes. Anti-CD45RA-APC-H7 (clone HI100, mouse IgG2bκ, BD Biosciences), and anti-CCR7- FITC (clone G043H7, mouse IgG2aκ, BioLegend) were used to determine memory populations on gated AIM + CD4+ Th1 and CD8+ CTL and Treg. Each dot shows the percentage of memory populations: terminally differentiated effector T cells (CD45RA + CCR7- TEMRA), median 2.66% on CD4+ Th1; median 44.75% on CD8+ CTL; and median 1.77% on Treg), effector memory T cells (CD45RA- CCR7- TEM), median 62.1% on CD4+ Th1, median 27.75% on CD8+ CTL, median 58.45% on Treg, and central memory T cells (CD45RA- CCR7+ TCM, median) 29.15% on CD4+ Th1, median 4.41% on CD8+ CTL, and median 22.70% on Treg).
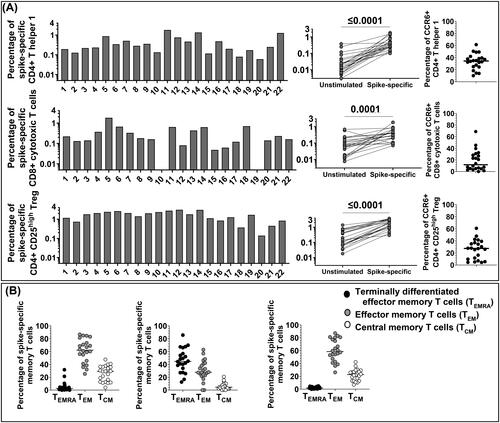
CD4+ Th1 and Treg expressed the chemokine receptor CCR6 in a considerable percentage (, right in panel A), suggesting T cell homing to the vascular endothelium, lung and gut epithelial cells and neurons.
The results were comparable with our previous report on CCR6 expression on spike-specific Treg enumerated in pregnant women convalescent from COVID-19 [Citation20].
Memory TEMRA, TEM, and TCM were analyzed in all the SARS-CoV-2 spike-specific T cell lineages (, panel B). CD4+ Th1 and Treg TEM were numerous and represented the largest spike-specific T cell memory population in circulation. CD4+ Th1 and Treg were also TCM in a significant percentage, unless CD8+ CTL that showed a TEMRA phenotype as previously reported in COVID-19 convalescent adults (reviewed in [Citation1,Citation5]) and by ourselves in multisystem inflammatory syndrome in children (MIS-C) and in COVID-19 convalescent pregnant women [Citation11,Citation20,Citation21].
SARS-CoV-2 spike-specific Treg do not revert from pro-inflammatory T cells as classical pTreg
Next, we asked about the ontogeny of the Treg that so efficiently expanded after mRNA-based vaccination. pTreg that revert from antigen-specific pro-inflammatory T cells are defined by the expression of the IL-7 receptor (IL-7R) [Citation22,Citation23] and revert to a CD45RA + phenotype (reviewed in [Citation13,Citation24]).
SARS-CoV-2 specific Treg lines have been obtained by oligo cloning primary PBMC cultures from subject #14, after addressing Treg specificity (in separate cultures) 24 h after stimulation. Treg have been expanded for 72 h after stimulation with peptides, harvested, washed, and diluted at the concentration of 10x104 cells/ml for oligo cloning. T cell lines have been then re-stimulated at the cell concentration of 10×104/well with the spike peptide mega pool. Autologous, irradiated PBMC served as APC in these experiments.
Three stable SARS-CoV-2 spike-specific oligoclonal Treg lines have been studied with monoclonal antibodies to surface molecules in the context of intracellular staining for FOXP3. The results obtained by gating on CD4+ CD25high T cells revealed a great expansion of SARS-CoV-2 spike-specific Treg upon res-stimulation (, panel A). Treg phenotype and purity was confirmed by intracellular FOXP3 expression on CD4+ CD25high T cells. Treg expressed CTLA-4 and PD-1 at different levels, depending upon Treg line, supporting the oligoclonality of the population. The expression of PD-1 is plastic on Treg, playing a role on the extent of effector regulatory mechanism by Treg during infections [Citation25].
Figure 2. Phenotype of SARS-CoV-2 spike-specific Treg. SARS-CoV-2 spike-specific Treg lines were generated from donor #14 and expanded in vitro from bulk cultures for phenotypical characterizion to determine their ontogeny, possibly reverting from pro-inflammatory T cells (pTreg) or arising from naïve T cells primed in tissues that synthetize spike proteins. (A) We used intracellular staining to detect FOXP3 (anti-FOXP3 PE, clone 259D, mouse IgG1k, Biolegend), and surface CTLA-4 (anti-CTLA-4 PE/Dazzle594, clone BNI3, mouse IgG2ak, Biolegend), PD-1 (anti-PD-1 BV605, clone EH12.2H7, mouse IgG1k, Biolegend), IL7Ra (anti-IL7Ra FITC, eBioRDR5, mouse IgG1k, eBioscience) and CD45RA (anti-CD45RA APC, clone HI100, mouse Ig2bk, eBioscience) on gated CD4+ CD25high Treg. CTLA-4 and PD-1 were measurable on Treg with minor differences between Treg lines. IL-7R and CD45RA were not expressed on SARS-CoV-2-specific Treg suggesting that Treg were not reverting from pro-inflammatory T cells repeatedly stimulated, but they differentiate from a naïve T cell repertoire. (B) To address the role of IL-2 in determining CTLA-4 and PD-1 expression on SARS-CoV-2 spike-specific Treg, we FACS sorted Treg from donor #1 after 72 h in vitro priming with spike peptides, prior to re-stimulation with agonistic antibodies to CD3 and CD28. FACS-sorted Treg were cultured with different regimens of IL-2 for 48 h prior to staining with specific monoclonal antibodies. The results suggested that Treg were pure, FOXP3 positive, and that the expression of Treg markers, CTLA-4 and PD-1 increased depending upon the amount of IL-2 in culture.
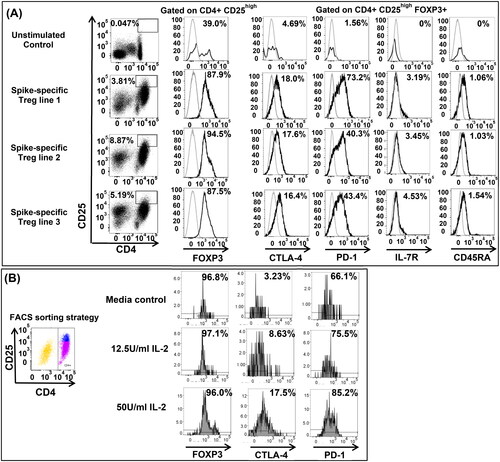
However, IL-7Ra and CD45RA, the markers that define pTreg were negative on Treg (, panel A).
These results suggested that this Treg population, phenotypically resembling thymic-derived nTreg, expanded from naïve T cells primed in tissues where the spike proteins were synthetized, following vaccination.
To address a possible dependency on the IL-2 in the environment in determining the expression of CTLA-4 and PD-1, we FACS sorted spike-specific Treg 72 h after cultures from subject #1, to obtain a pure Treg population, prior to restimulation with agonistic antibodies to CD3 and CD28, in the absence of IL-2, or with different concentrations of IL-2. The results suggested that the expression of both Treg markers, CTLA-4 and PD-1, increased on Treg depending upon IL-2 feeding conditions (, panel B).
SARS-CoV-2 spike-specific Treg memory is fully developed as early as after the second vaccine injection with little changes over time regardless from further vaccine boosts
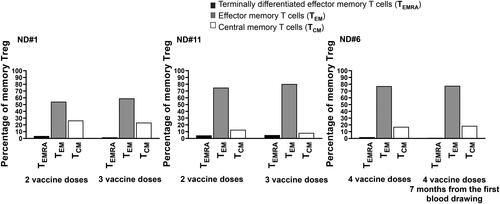
Treg expanded and developed memory as early as after the second vaccine injection with no significant differences in the 22 donors. To define the role of a third or a fourth vaccine dose in the development of Treg memory, we studied three subjects consecutively, subjects #1, #6 and #11 () months apart. After PBMC in vitro priming with the spike peptide mega pool, we looked at Treg memory 24 h after stimulation to be compared with the previous AIM assay, studied months earlier. The results in support that Treg TEM and TCM, already numerous in the first blood drawing, did not further increase with additional vaccine boosts. Moreover, Treg TEM and TCM where not higher in subject #6 that received four injections and has been studied seven months apart.
Figure 3. TEM and TCM Treg develop early on during vaccination with little changes after further vaccine boosts. The SARS-CoV-2 spike-specific memory Treg repertoire has been explored consecutively, months apart, in donors # 1 and #11 studied after the second and third injection, respectively, and #6 that received four vaccine administration and has been studied seven months apart. Numerous TEM and TCM Treg were found in circulation after two vaccine injection in two subjects, #1 and 11, left and middle panels, with little changes after the third dose. Donor 6, studied at two time points after the fourth dose, showed a comparable development of Treg memory as the other two subjects studied after two vaccine injections (right panel).
FACS-sorted SARS-CoV-2 spike-specific Treg down-regulate IFNγ secretion by autologous naïve T cells and SARS-CoV-2 spike-specific CD4+ Th1
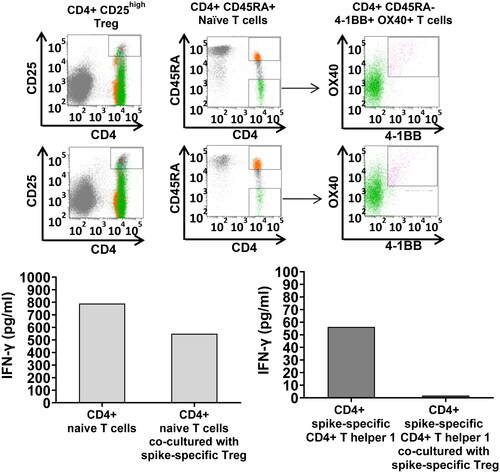
To explore the role of Treg in regulating naïve T cell differentiation and spike-specific pro-inflammatory T cells, we performed co-culture experiments with SARS-CoV-2 spike-specific FACS-sorted Treg, naïve T cells and AIM + T cells. PBMC from two different donors, #1 and #16, have been stimulated 72 h with spike peptide mega pool in 24 well plates at the cell concentration of 4×106 cells/well prior to sorting with specific monoclonal antibodies. Cell numbers was a limitation in these experiments: from subject #1 we FACS-sorted SARS-CoV-2-specific Treg and naïve CD4+ CD45RA + T cells; from subject #16 we FACS sorted SARS-CoV-2 spike-specific Treg and AIM + CD4+ Th1 cells. The cell ratio in co-culture was 1:1 (10,000 T cell/w in the naïve T cell/Treg co-culture, and 1000 T cell/w in the CD4+ Th1 AIM+/Treg co-cultures).
Down-regulation of IFNγ secretion 24 h after stimulation with agonistic anti-CD3 and anti-CD28 antibodies by naïve CD4+ T cells and CD4+ Th1 co-cultured with Treg served as read outs in these experiments.
The results indicated a strong inhibition of IFNγ production by naïve T cells when co-cultured with Treg (left panel) and by CD4+ Th1 when co-cultured with Treg (, right panels).
Figure 4. Treg-mediated down-regulation of IFNγ production by naïve T cells and AIM + T cells in co-culture. SARS-CoV-2 spike-specific Treg, naïve CD4+ T cells, AIM + CD4+ Th1 have been FACS-sorted by flow cytometry with specific monoclonal antibodies after in vitro priming with the spike mega pool (FACS sorting strategy shown in the upper panel) and co-cultured either with autologous CD4+ naïve T cells (left panels) or SARS-CoV-2 spike-specific CD4+ Th1 (right panels) at 1:1 T cell ratio (25,000 T cells in the naïve T cell/Treg co-cultures and 1,000 T cells in the AIM+/Treg co-cultures). IFNγ secretion, measured in culture supernatants by ELISA, served as a read out in these experiments. The results indicated that Treg down-regulate both, naïve T cell polarization toward a pro-inflammatory phenotype, and IFNγ secretion by the AIM + T cells.
Discussion
Little is known about the expansion of antigen specific Treg following an anti-viral vaccination. Depending upon the physiopathology of the pathogen that the vaccine aims to prevent, the expansion of Treg could be highly beneficial and a parameter to evaluate when studying the potency of a vaccine.
The extent of SARS-CoV-2 spike-specific Treg expansion during COVID-19 determines the severity of the disease: a collection of reports emphasized the role of Treg in down-sizing the exuberant inflammation in acute COVID-19 subjects (reviewed in [Citation10]). Moreover, Treg isolated from SARS-CoV-2-infected adults with severe symptomatology showed a peculiar phenotype that jeopardized their immune regulatory functions [Citation9].
In this study, we enumerated SARS-CoV-2 spike-specific Treg in healthy vaccine recipients and we defined their unique phenotype. Treg regulated naïve T cell polarization and down-regulated IFNγ production by pro-inflammatory T cells.
The study population involved 22 vaccine recipients, where we found SARS-CoV-2 spike-specific Treg fully expanded as soon as after two vaccine doses with Moderna mRNA 1273 and/or Pfizer/BioNTechBNT162b2, and in a large percentage with a memory phenotype.
The memory phenotype has been found TEM and TCM, comparable to SARS-CoV-2 spike-specific CD4+ Th1. No significant differences in the development of Treg memory were observed in vaccine recipients that received two or four vaccine boosts, nor in the three subjects that had COVID-19, beside vaccination.
As a validation, we enrolled two times, several months apart, three subjects that we studied after the second and third injection or after the fourth injection.
SARS-CoV-2 spike-specific Treg cells showed a unique phenotype, resembling thymic-derived nTreg. The lack of expression of the IL-7 R and CD45RA, suggested that this Treg population did not arise from pro-inflammatory T cells repeatedly stimulated, as classical pTreg that revert from pro-inflammatory T cells, but were naïve T cells, primed in tissues where the spike synthesis occurred.
It is well-know that the role of Treg is not limited in regulating the immunity: Treg prevent fibrosis, protect tissues homeostasis and, by secreting IL-10, regulate several lymphokines and chemokines [Citation12]. Echoing this concept, a large percent of SARS-CoV-2 spike-specific Treg expressed CCR6, a homing receptor for endothelial cells, lung and gut epithelial cells and neurons: this is why we believe that Treg maintain tissue immune homeostasis in controlling inflammation when vaccinated subjects get exposed to COVID-19.
Another peculiarity of these Treg, phenotypically with high intracellular FOXP3 as classical Treg, was the expression of CTLA-4 and PD-1, that we found up-regulated depending upon IL-2 regimen.
Possibly, a death signal (PD-1/PD-1L) by SARS-Cov-2 specific Treg regulates anti-viral pro-inflammatory T cells only when an over exuberant expansion of IL-2-secreting T cells occurs. This is similar to the murine model of Toxoplasma gondii infection where PD-1/PD-1L interactions modulate Treg functions. Our data are also supported by recent findings on the dynamic transcriptional activity, and chromatin remodeling of Treg depending upon duration of IL-2 signaling that controls not only PD-1 expression, but other effector regulatory functions [Citation26].
Functionally, Treg down-regulated in co-cultures naïve T cell differentiation toward a pro-inflammatory phenotype and IFNγ production by SARS-CoV-2 spike-specific CD4+ Th1, suggesting regulation of inflammation.
Future studies will determine the HLA restriction and the peptides recognized by SARS-CoV-2 spike-specific Treg, that are anticipated to differ from the peptides recognized by pro-inflammatory T cells.
The expansion of our work leads to a new aspect to be evaluated while studying the potency of a vaccine.
In conclusion, the strength of this study is the report of a unique population of Treg that arises in subjects vaccinated for COVID-19 protection, enumerated, phenotypically and functionally characterized in vaccine recipients.
The limitation of the study is the lack of access to human tissues that prevented to conclusively demonstrate the ontogeny of SARS-CoV-2 spike-specific Treg and their crosstalk, not only with naïve and pro-inflammatory T cells, but also with endothelial cells, epithelial cells, and neurons, as suggested by their phenotype.
Supplemental Material
Download MS Word (1.3 MB)Acknowledgments
The authors thank the subjects that participated to the study.
Disclosure statement
The authors declare no conflict of interest. LJI has filed for patent protection for various aspects of vaccine design and identification of specific epitopes.
Data availability statement
The authors confirm that the data supporting the findings of this study are available within the article.
Additional information
Funding
References
- Merad M, Blish CA, Sallusto F, et al. The immunology and immunopathology of COVID-19. Science. 2022;375(6585):1–9.
- Felgenhauer U, Schoen A, Gad HH, et al. Inhibition of SARS-CoV-2 by type I and type III interferons. J Biol Chem. 2020;295(41):13958–13964.
- Kim Y-M, Shin E-C. Type I and III interferon responses in SARS-CoV-2 infection. Exp Mol Med. 2021;53(5):750–760.
- Viox EG, Hoang TN, Upadhyay AA, et al. Modulation of type I interferon responses potently inhibits SARS-CoV-2 replication and inflammation in rhesus macaques. Sci Immunol. 2023;8(85):eadg0033.
- Sette A, Crotty S. Adaptive immunity to SARS-CoV-2 and COVID-19. Cell. 2021;184(4):861–880.
- Krammer F. SARS-CoV-2 vaccines development. Nature. 2020;586(7830):516–527.
- Zhang Z, Mateus J, Coelho CH, et al. Humoral and cellular immune memory to four COVID-19 vaccines. Cell. 2022;185(14):2434–2451.e17.
- Jarjour NN, Masopust D, Jameson SC. T cell memory: understanding COVID-19. Immunity. 2021;54(1):14–18.
- Harb H, Benamar M, Lai PS, et al. Notch4 signaling limits regulatory T-cell-mediated tissue repair and promotes severe lung inflammation in viral infections. Immunity. 2021;54(6):1186–1199.e7.
- Xu Z, Jiang X, Dai X, et al. The dynamic role of FOXP3+ tregs and their potential therapeutic applications during SARS-CoV-2 infection. Front Immunol. 2022;13:916411–916433.
- Hsieh LE, Grifoni A, Dave H, et al. SARS-CoV-2-specific T cell responses and immune regulation in infected pregnant women. J Reprod Immunol. 2022;149:103464–103471.
- Dominguez-Villar M, Hafler DA. Regulatory T cells in autoimmune diseases. Nat Immunol. 2018;19(7):665–673.
- Sakaguchi S, Mikami N, Wing JB, et al. Regulatory T cells and human diseases. Annu Rev Immunol. 2020;38(1):541–566.
- Sallusto F, Impellizzieri D, Basso C, et al. T-cell trafficking in the central nervous system. Immunol Rev. 2012;248(1):216–227.
- Grifoni A, Sidney J, Zhang Y, et al. A sequence homology and bioinformatic approach can predict candidate targets for immune responses to SARS-CoV-2. Cell Host Microbe. 2020;27(4):671–680 e672.
- Grifoni A, Weiskopf D, Ramirez SI, et al. Targets of T cell responses to SARS-CoV-2 coronavirus in humans with COVID-19 disease and unexposed individuals. Cell. 2020;181(7):1489 e1415–1501 e1415.
- Reiss S, Baxter AE, Cirelli KM, et al. Comparative analysis of activation induced marker (AIM) assays for sensitive identification of antigen-specific CD4 T cells. PLoS One. 2017;12(10):e0186998.
- Mateus J, Grifoni A, Tarke A, et al. Selective and cross-reactive SARS-CoV-2 T cell epitopes in unexposed humans. Science. 2020;370(6512):89–94.
- Weiskopf D, Schmitz KS, Raadsen MP, et al. Phenotype and kinetics of SARS-CoV-2-specific T cells in COVID-19 patients with acute respiratory distress syndrome. Sci Immunol. 2020;5(48):eabd2071.
- Hsieh LE, Grifoni A, Shimizu C, et al. T cells in multisystem inflammatory syndrome in children have a predominant CD4+ T helper response to SARS-CoV-2 and numerous virus-specific CD4- CD8- double negative T cells. Int J Mol Sci. 2022;23(13):7219–7233.
- Hsieh L-E, Grifoni A, Sidney J, et al. Characterization of SARS-CoV-2 and common cold coronavirus-specific T-cell responses in MIS-C and Kawasaki disease children. Eur J Immunol. 2022;52(1):123–137.
- Zielinski CE, Mele F, Aschenbrenner D, et al. Pathogen-induced human Th17 cells produce IFN-gamma or IL-10 and are regulated by IL-1beta. Nature. 2012;484(7395):514–518.
- Duhen T, Duhen R, Lanzavecchia A, et al. Functionally distinct subsets of human FOXP3+ treg cells that phenotypically mirror effector Th cells. Blood. 2012;119(19):4430–4440.
- Sakaguchi S, Vignali DA, Rudensky AY, et al. The plasticity and stability of regulatory T cells. Nat Rev Immunol. 2013;13(6):461–467.
- Perry JA, Shallberg L, Clark JT, et al. PD-L1-PD-1 interactions limit effector regulatory T cell populations at homeostasis and during infection. Nat Immunol. 2015;518:197–206.
- Moro A, Gao Z, Wang L, et al. Dynamic transcriptional activity and chromatin remodeling of regulatory T cells after varied duration of interleukin-2 receptor signaling. Nat Immunol. 2022;23(5):802–813.