
ABSTRACT
This study investigated the effect of the odors generated by the glycine/glucose Maillard reaction and the potent odorants 2,3-dimethylpyrazine (3DP) and 2,5-dimethyl-4-hydroxy-3(2H)-furanone (DMHF) on the human mood and integrative physiological activity. The score of certain subjective moods, especially anger-hostility, and tension-anxiety were decreased significantly after inhalation of the Maillard reaction sample and DMHF, and fatigue-inertia mood was also significantly decreased by DMHF, suggesting a sedative effect of these odors on mood, while 3DP had no effect. Miosis rate and fingertip temperature increased significantly following inhalation of the odor from the Maillard reaction sample and both potent odorants, suggesting that the parasympathetic nervous system dominates through suppression of the sympathetic activity. The physiological relaxing effect of these odors was also confirmed by decreased flicker frequency value and decreased oxyhemoglobin in the prefrontal cortex.
Graphical Abstract
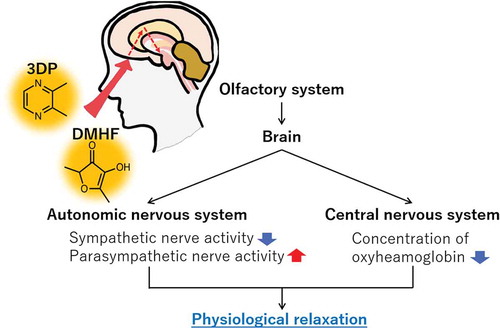
Inhalation of 3DP or DMHF induced parasympathetic dominance by suppression of sympathetic nervous system and led to a decrease in oxyhemoglobin in the prefrontal cortex.
The Maillard reaction is a non-enzymatic reaction between carbonyl and amine functional groups and frequently occurs in thermally processed food like bread, roasted coffee beans, cocoa, and black beer among others, causing browning [Citation1]. The Maillard reaction produces hundreds of low-molecular weight odorous molecules which play an important role in determining the characteristic flavor of processed food.
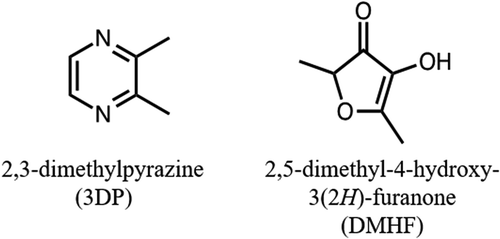
Odors have been demonstrated to affect mood, physiology, and behavior, due to their interaction with the limbic, autonomic, and immune systems [Citation2]. They can exert their effects either through direct activation of olfactory receptor cells which send the olfactory information to various parts of the brain or indirectly through diffusion into the bloodstream through transdermal absorption or digestion, thereby crossing the blood-brain barrier and subsequently affecting brain neural activity. Physiological and psychological properties of essential oils such as lavender [Citation3], rosemary [Citation4], and citrus peels [Citation5–Citation7] have been studied broadly. However, few studies have focused on the odors generated by the Maillard reaction. Suganuma et al. [Citation8] reported that roasted barley tea shortened the passage time of whole blood, with alkylpyrazines possibly affecting blood fluidity. Alkylpyrazines are considered to be odorants generated by the Maillard reaction between maltose and amino acids present in malt, during the roasting of barley. In addition to impacting the physiology through oral administration and digestion, we propose that odors generated by the Maillard reaction may also induce physiological changes directly through the olfactory system. Previously, the effects of odors generated by the glycine/glucose model systems on mood and brainwaves have been investigated in humans. Following inhalation, “tense” and “fatigue” negative moods decreased, while the β brainwave distribution increased [Citation9]. A less depressed mood, greater relaxation, increased β power, and enhanced performance have been reported following exposure to lavender odor [Citation10], suggesting that the odors generated by the glycine/glucose Maillard reaction could have similar sedative effects as lavender odor. Moreover, we identified 2,3-dimethylpyrazine (3DP) and 2,5-dimethyl-4-hydroxy-3(2 H)-furanone (DMHF) as potent odorants in the glycine/glucose Maillard reactant. The chemical structures of 3DP and DMHF are shown in . Each compound inhalation affected brainwave; especially, the β brainwave distribution was found to increase after smelling 3DP. However, more evidence is needed to evaluate these effects. We hypothesized that the odor generated by the glycine/glucose Maillard reaction, 3DP, and DMHF could affect the activity of the autonomic/central nervous system to change a mood state.
Figure 1. The chemical structures of 3DP and DMHF.
The aim of the present study was to investigate the effect of odors generated by the glycine/glucose Maillard reaction and the potent odorants (3DP and DMHF) in humans using integrative physiological methods. Pupil diameter and peripheral skin temperature were used to evaluate the autonomic nervous activity, while the activity of the central nervous system was evaluated by near-infrared spectroscopy and a flicker test.
Materials and methods
Chemicals
Glucose, glycine, and sodium carbonate were obtained from Kanto Chemical Co. (purity: >95%, Tokyo, Japan). 3DP and DMHF were obtained from Sigma Aldrich Japan (purity: >99%, Tokyo, Japan).
Participants
Twenty-six healthy participants (thirteen males and thirteen females, ages 21–24 years) were recruited for this study. All measurements were conducted in a sensory room with a controlled ambient temperature (25°C). The objective and methods used in this study were explained to the participants and their consent obtained. All measurements were performed according to the ethics code of the Kitasato University (ethics code: 15–201).
Preparation of the Maillard reaction sample
Detailed procedures for producing the Maillard reaction sample have been previously described [Citation9]. Briefly, equimolar solutions of glucose (1.0 mol/L) and glycine (1.0 mol/L) were prepared in 0.25% (w/v) sodium carbonate buffer and pH adjusted to 9 using NaOH. Each sample was poured into screw-capped glass tubes and heated at 90°C for 30 min in an oil bath. After heat treatment, samples were immediately cooled in ice water and stored at 4°C until further use.
Ten milliliters of the Maillard reaction sample was poured into an amber vial (opening diameter, 35 mm; height, 73 mm; content, 50 mL) and the lid of the vial was closed. It was left to stand for 30 min, then used for the subsequent experiments.
Preparation of 3DP and DMHF samples
The concentrations of 3DP and DMHF in the Maillard reaction sample were quantified as described in our previous study [Citation9]. The 3DP and DMHF samples were prepared as follows: DMHF was diluted with distilled water to a concentration of 58 mg L−1 and 3DP was diluted with distilled water to a concentration of 0.9 mg L−1. Ten milliliters of each odorant was poured into amber vials (as described above), and the lids of the vials were closed. Then, the samples were left to stand for 30 min.
Presentation of odor samples
Each vial with a sample was presented to each participant’s nose. The distance between the nose and the opening of the vial was approximately 10 mm. The participants were seated comfortably on an armchair during the experiment and breathed the vapor from the sample for 2 min. The participants were presented with control and odor samples in a blind manner. However, they were informed that the odor samples were odorants derived from the Maillard reaction in food.
Study procedure
The participants were seated in a sensory room for 5 min for acclimation to the experimental environment before each measurement. Measurements 1 to 3 (1: POMS 2® examination; 2: pupillary light reflex and fingertip temperature measurements; 3: flicker test and near-infrared spectroscopy measurement) were all performed on different days. The presentation of odor samples was randomized, and the measurement was performed on another day. The methods of each measurement are described below.
Profile of mood states
The Profile of Mood States (2nd edition, POMS 2®) was used to examine the mood changes before and after smelling each odorant from the Maillard reaction sample. The POMS 2® consists of seven sets of mood states: anger-hostility (AH), confusion-bewilderment (CB), depression-dejection (DD), fatigue-inertia (FI), tension-anxiety (TA), and friendliness (F). POMS 2® was standardized in the “past week including today” timeframe. After breathing clean air for 2 min (control), the participants underwent POMS 2® examination; then, they inhaled the vapors of each odor sample for 2 min and underwent POMS 2® examination again. Participants were asked to rate these mood states on a scale from 0 (not at all) to 4 (extremely), control and each odor sample. The total score for each state was calculated by adding the corresponding set of adjectives.
Pupillary light reflex measurement
The pupillary light reflex was measured to examine the state of the autonomic nervous activity. Measurement was performed between 09:00 and 13:00 hours, because pupillary reaction is relatively stable during this time [Citation11]. The participants breathed natural air for 2 min as a control followed by measurement of the miosis rate, which is the ratio of the pupil diameter before and after light stimulation, measured using an Iriscorder Dual C10641 (Hamamatsu Photonics, Shizuoka, Japan). The miosis rate was measured for inhalation of each sample odor as well and compared to the control in order to evaluate the effect of the odorants on the autonomic nervous system.
Fingertip temperature measurement
Fingertip temperature was measured with a thermometer (LT8A, Gram Corporation, Saitama, Japan). Participants were asked to enter the sensory room and settle comfortably in a chair for a certain duration of time. The fingertip temperature was first measured once as a control before odor inhalation. Subsequently, the participants were instructed to inhale the vapor of each sample for 2 min.
Flicker test
Flicker frequency value (FFV) was determined with a green flickering light stimulus of 8.7 mm diameter (Handy flicker HF-II, Neitz Instruments Co., LTD, Tokyo, Japan). The participants were comfortably seated with the light adjusted at about 25 cm away from their eyes. The test started at a frequency of 70 ± 2 Hz, at which the light appeared to be completely steady to all participants. After breathing natural air for 2 min (control) or inhaling the vapors of each sample for 2 min, the participants were instructed to press the button with their dominant thumb to lower the frequency of the light progressively, and to release immediately upon detection of the flickering light. The frequency of the flickering light was used as FFV. The test was repeated five times, and the highest and lowest FFVs were excluded. The remaining three FFVs were averaged and compared with that of the control.
Near-infrared spectroscopy
Changes in the concentration of oxyhemoglobin (oxy-Hb) in the frontal cortex of the brain were assessed by near-infrared spectroscopy (NIRS; Spectratech OEG-16, Spectratech Inc., Tokyo, Japan). Two wavelengths (840 nm and 770 nm) were used to record the changes in the absorption of oxy-Hb. The 16 channels (see ) and the source and detector were separated by 3 cm and the center of the probe holder was located at the frontopolar central position in the International 10/20 System. The sampling interval was set at 650 ms. The oxy-Hb concentration (mM/mm) was calculated using the Modified Lambert–Beer Law. The Spectratech OEG-16 probe holder was attached to the participants’ heads, who were instructed to sit in a relaxed position and look at a cross mark fixed on the wall (2 m from the seat), until the end of the experiment. The concentration of oxy-Hb was recorded twice for a total of 4 min, 2 min before odor inhalation (breathing natural air), and 2 min during odor inhalation. The mean oxy-Hb concentration was calculated before and during odor presentation.
Figure 2. The location design of a measurement channel of a probe holder in near-infrared spectroscopy.
Statistical analysis
All data are expressed as mean ± standard error (SE), and were statistically analyzed using the paired t-test (student’s t-test) after confirming the normal distribution in SPSS [Citation12,Citation13]. A probability value (p) of <0.05 was considered significant.
Results
Subjective evaluation: profile of mood states 2nd edition (POMS)
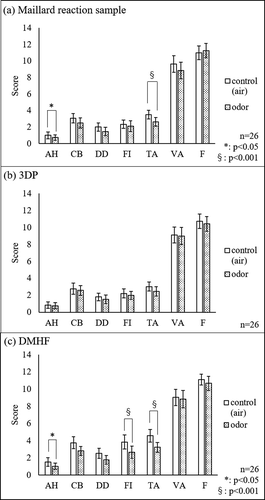
POMS 2® was used in the current study, which is widely used in evaluating the effects of odor on mood and is sensitive to transient mood shifts [Citation14,Citation15]. As shown in , friendliness (F) score tended to increase in the Maillard reaction sample, while the scores of other mood states tended to decrease after odor inhalation. In particular, the anger-hostility (AH) and tension-anxiety (TA) scores decreased significantly (AH: p < 0.05, TA: p < 0.001) after odor inhalation. also shows the changes in score after inhalation of 3DP () and DMHF (). AH (anger-hostility, p < 0.05), FI (fatigue-inertia, p < 0.001), and TA (tension-anxiety, p < 0.001) scores decreased significantly after DMHF inhalation. However, no significant changes were observed after smelling 3DP.
Figure 3. POMS 2® scores for air (control) and odor inhalation.
Effects of odor on autonomic nervous activity
Pupillary light reflex
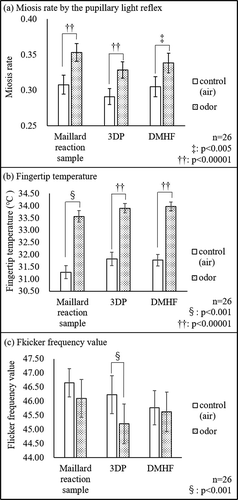
The miosis rate is defined as the ratio of the pupil diameter before and after light stimulation and is measured for both the inhalation of natural air (control) and sample odor (). The miosis rate for the control was 0.31 ± 0.01 and increased significantly up to 0.35 ± 0.01 (p < 0.00001) after odor inhalation from the Maillard reaction sample. Miosis rate increased significantly to 0.328 ± 0.012 after 3DP inhalation (p < 0.00001) compared to 0.291 ± 0.011 for the control and increased significantly to 0.338 ± 0.014 after DMHF inhalation (p < 0.005) compared to 0.305 ± 0.014 for the control.
Figure 4. Effect of the odor of the Maillard reaction sample, 3DP, and DMHF on physiological parameters.
Fingertip temperature
The change in fingertip temperature (FT) of the dominant index finger after odor inhalation is shown in . The average fingertip temperature in the control was 31.28°C, and it increased by 2.28 ± 0.24°C after inhalation of the odor from the Maillard reaction sample for 2 min (p < 0.001). Inhalation of 3DP increased FT by 2.08 ± 0.19°C (p < 0.00001), and inhalation of DMHF increased FT by 2.20 ± 0.19°C (p < 0.00001), in comparison to the control.
Effect of odors on the central nervous system activity
Flicker test
shows the mean ± SD flicker frequency value (FFV). Although a small decrease in FFV was observed after odor inhalation, FFV did not differ significantly between the two conditions. FFV tended to decrease after smelling DMHF, but the changes were not statistically significant, while a significant decrease (−1.04 ± 0.28) was observed after smelling 3DP (p < 0.001).
Oxy-Hb changes measured by NIRS
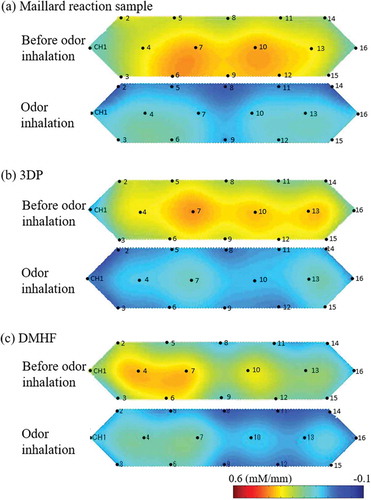
shows the mean changes in oxy-Hb concentration in each channel measured 2 min before odor inhalation and 2 min during odor inhalation. Inhalation of odors of the Maillard reaction sample, 3DP, and DMHF led to a significant decrease in oxy-Hb concentration in all channels. In particular, oxy-Hb concentration at the overall prefrontal cortex located at channels 1, 4, 6, 7, 10, 12, 13, 15, and 16 decreased due to inhalation of 3DP. Decrease in the oxy-Hb concentrations was especially pronounced at the right and left prefrontal cortex located at channels 1, 3, 6, 13, 15, and 16 in the Maillard reaction sample and at the right prefrontal cortex located at channels 1, 4, 6, and 7 in DMHF. depicts the mean changes in oxy-Hb concentration (hemodynamic responses). Inhalation of the odor of the Maillard reaction sample, 3DP, and DMHF led to a decrease in oxy-Hb concentration in the prefrontal cortex.
Table 1. Changes in oxy-Hb concentration (mM/mm) before and during odor inhalation of the Maillard reaction sample, DMHF, and 3DP (n = 26).
Figure 5. Illustration of changes in oxy-Hb concentration (hemodynamic responses) by NIRS.
Discussion
Many studies focused on the physiological and psychological effects of essential oils have shown that potent odorants can produce similar effects as essential oil [Citation3]. We previously observed that the odor generated from the glycine/glucose Maillard reaction has sedative effects in humans, and 3DP and DMHF were identified as the strongest potent odorants in this sample [Citation9]. In order to identify the active odorants in the Maillard reaction sample, the effects of these two odorants on mood and autonomic/central nervous system were investigated in the present study.
We used POMS 2® to evaluate mood changes and observed that the scores in AH and TA moods decreased significantly after inhalation of the Maillard reaction sample odor, suggesting a sedative effect on mood. These data are consistent with our previous findings [Citation9] where the odor significantly decreased the negative “tense” mood. Similar to the Maillard reaction sample, the DMHF odor decreased FI, AH, and TA negative mood scores significantly. DMHF was one of the strongest odorants detected from the Maillard reaction sample and it could be the main causative agent for mood change. Given the direct connection between the olfactory system and the limbic system, odors can produce psychological effects as well. In particular, the hippocampus and amygdala structures have been known to play a critical role in emotional processing, memory functions, and odor preference. On the other hand, no significant changes in POMS 2® score were observed after smelling 3DP, probably due to its low concentration (0.9 mg L−1). The 3DP threshold was reportedly 2500 ppb in water [Citation16]. The subjects who participated in this study answered that the 3DP odor at this concentration was too weak to be perceived and described while answering a simple oral question for the POMS 2®. Although some studies have shown that changes in mood can still be observed even if an odorant is below the detection threshold [Citation17], our results suggest that it is necessary for detectable levels of 3DP to be present in order to alter mood. In contrast, the DMHF threshold was reportedly 60 ppb in water [Citation18]. Since the odors of the glycine/glucose Maillard reaction and DMHF resulted in relaxing and positive effects on mood, sedative effects on the central and autonomic nervous system could also be expected.
Plasma norepinephrine level, mean arterial blood pressure, and heart rate were used to assess autonomic nervous activity in humans as these parameters reflect the activity of the autonomic nervous system, but are also easily influenced by environmental stress caused by the measuring equipment. Noninvasive pupil diameter was measured in order to obtain highly reproducible and reliable data. It is well known that pupillary size is regulated by two antagonistic smooth muscle systems, the sympathetic nervous system which enlarges the pupil by activating the dilator muscle and its opponent the parasympathetic nervous system which regulates the sphincter muscle [Citation19]. Parasympathetic dominance leads to an increase in the miosis rate. In our study, the miosis rate increased significantly after inhalation of the Maillard reaction sample, 3DP, and DMHF, indicating that the odors may affect the autonomic nervous activity, and cause the parasympathetic branch of the autonomic nervous system to become dominant [Citation13,Citation20]. Furthermore, our study showed that the pupillary light reflex measurement is an appropriate experimental technique in evaluating autonomic nervous activity changes caused by food odor stimulations.
The parasympathetic dominance observed in our study could be due to an elevation in the parasympathetic activity, or a suppression of the sympathetic activity. In order to address this question, we investigated the changes in the sympathetic nervous activity through the measurement of FT since arterioles of the fingertip skin have only sympathetic constrictor nerves, which control the skin temperature by constantly maintaining the cutaneous microcirculation [Citation21]. Indeed, sympathetic stimuli such as cold pressure, pain, and mental workload have been reported to cause vasoconstriction and skin temperature drop [Citation22]. In our results, FT increased significantly following the presentation of the Maillard reaction sample, 3DP and DMHF, suggesting that the sympathetic nervous activity was suppressed by these odors. Although the blood flow was not evaluated in this study, improvement in the cutaneous microcirculation and increase in blood fluidity may have occurred in the fingers. The results from the current study show that the odor generated by the Maillard reaction might induce stress-relieving effects similar to green tea [Citation23] and ginger [Citation24], through suppression of the sympathetic nervous activity. Previously, we demonstrated that inhalation of DMHF induced parasympathetic dominance by suppressing renal sympathetic nerve activity in Wistar rats. From the results in this study, it was shown that the similar effects of DMHF on human autonomic nervous system [Citation25].
The sedative effects on the autonomic activity indicate that the odor generated from the Maillard reaction may also suppress cortical activation resulting a sedative effect on the central nervous system. To investigate central nervous activity, we measured the changes in oxy-Hb concentration in the prefrontal cortex using NIRS, and applied the flicker test. Brain activity leads to an increase in oxygen consumption, accompanied by an increase in cerebral blood flow [Citation26,Citation27]. The observed decrease in oxy-Hb concentration during odor presentation indicates that odors of the Maillard reaction sample, 3DP, and DMHF may induce sedation in the central nervous system. Oxy-Hb changes have been considered to be an index of cerebral activity. Data obtained in the present study suggest that dominance in the parasympathetic autonomic nervous system leads to a decline in oxy-Hb in the prefrontal cortex. It has been reported that oxy-Hb concentration changes detected by the NIRS also reflect blood flow and blood oxygenation in the brain tissue and head surface tissue [Citation28]. However, since NIRS generally images brain activity in the skull bones, it is hypothesized that the prefrontal cortex is hardly affected by the muscle tissue and adipose tissue of the head surface layer. Moreover, in the present study, the skin temperature in the central forehead was also detected when measuring fingertip temperature but it confirmedly remained unchanged (data not shown). Thus, it could be concluded that skin tissue-derived blood flow changes were hardly observed during the NIRS measurement. In contrast, breathing could affect NIRS measurements. In this study, pupillary light reflex and fingertip temperature measurements were performed to detect autonomic nerve activity and evaluated even during uncontrolled breathing. In accordance with this condition, NIRS and flicker test were conducted without any strict breathing control (like using a metronome). However, the participants were instructed not to take deep breaths and not to change their way of breathing. The effect of breathing frequency and depth on odor sample inhalation during the NIRS measurements might need further investigation.
A significant decrease in FFV was observed after smelling 3DP. The flicker frequency is considered to reflect the excitation of the brain function (visual field, finger motor function, etc.). It seems likely that the brain function is temporarily recovered by the strong sedative effect of 3DP. The results suggested that 3DP could relieve brain function excitation. Although the differences were not significant, minor FFV decreases were observed following the presentation of other odors from the Maillard reaction sample and DMHF. We hypothesize from the above-described observation and the concentration of 3DP produced by the Maillard reaction that it is possible that 3DP exerted a stronger sedative effect than DMHF.
DMHF decreased negative moods, increased parasympathetic nervous activity, and reduced sympathetic activity. 3DP had similar sedative effects on autonomic and central nervous activity, it had no effects on mood. Comparison with the effects of the odor of the glycine/glucose Maillard reaction showed that DMHF could be the causative factor for mood changes, and both DMHF and 3DP could be the contributors responsible for the sedative effects on the central and autonomic nervous system. In this study, we did not investigate the physiological effects of each potent odorant at different odor intensities. However, it is interesting that 3DP and DMHF both induce a sedative effect despite their different odor qualities (3DP: nutty, cooked rice-like odor; DMHF: caramel-like odor, sweet [Citation9]) and concentrations in the glycine/glucose Maillard reactant. As mentioned in the introduction section, there are only few studies on the physiological effects of odors generated by the Maillard reaction at the food level. However, no report has revealed yet the key odor component. Therefore, to the best of our knowledge, this study clarifies for the first time the aforementioned physiological effects at the single odorant level and demonstrate the sedative effect through an integrative evaluation.
DMHF and 3DP have been detected in various kinds of processed food where the Maillard reaction occurs, and are also commercially available for use as food additives. Data from the present study suggest that apart from increasing favorable sensory properties of foods, mental and physiological relaxing effects can also be expected from the inhalation of these odorants. Moreover, since the Maillard reaction occurs frequently in food processing, such effects can also be expected in behaviors like baking and cooking.
Conclusion
This study reveals that odors generated by the glycine/glucose Maillard reaction can promote mood, and can induce physiological relaxation by suppressing the sympathetic activity. 3DP and DMHF, shown to be primary contributors to the odor profile of the glycine/glucose Maillard reaction sample [Citation9], could be correlated with observed mood or physiological changes.
Authors’ contributions
Motoko Ohata: Conceptualization, Writing original draft, Project administration, Funding acquisition.
Lanxi Zhou: Investigation, Writing original draft.
Yukihiro Yada: Reviewing, Supervision.
Issei Yokoyama: Investigation, Writing original draft.
Keizo Arihara: Reviewing, Supervision.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Morales FJ, Somoza V, Fogliano V. Physiological relevance of dietary melanoidins. Amino Acids. 2012;42:1097–1109.
- Herz RS. Aromatherapy facts and fictions: a scientific analysis of olfactory effects on mood, physiology and behavior. Int J Neurosci. 2009;119:263–290.
- Tanida M, Niijima A, Shen J, et al. Olfactory stimulation with scent of lavender oil affects autonomic neurotransmission and blood pressure in rats. Neurosci Lett. 2006;398:155–160.
- Burnett KM, Solterbeck LA, Strapp CM. Scent and mood state following an anxiety-provoking task. Psychol Rep. 2004;95:707–722.
- Kiecolt-Glaser JK, Graham JE, Malarkey WB, et al. Olfactory influences on mood and autonomic, endocrine, and immune function. Psychoneuroendocrinology. 2008;33:328–339.
- Lehrner J, Marwinski G, Lehr S, et al. Ambient odors of orange and lavender reduce anxiety and improve mood in a dental office. Physiol Behav. 2005;86:92–95.
- Nagai K, Niijima A, Horii Y, et al. Olfactory stimulatory with grapefruit and lavender oils change autonomic nerve activity and physiological function. Auton Neurosci. 2014;185:29–35.
- Suganuma H, Inakuma T, Kikuchi Y. Amelioratory effect of barley tea drinking on blood fluidity. J Nutr Sci Vitaminol. 2002;48:165–168.
- Zhou L, Ohata M, Arihara K. Effects of odor generated from the glycine/glucose Maillard reaction on human mood and brainwaves. Food Funct. 2016;7:2574–2581.
- Field T, Diego M, Hernandez-Reif M, et al. Lavender fragrance cleansing gel effects on relaxation. Int J Neurosci. 2005;115:207–222.
- Yamaguchi S. The effects of acupuncture on pupillary dynamics. J Jpn Soc Balneol Climatol Phys Med. 1995;58:233–240.
- Dayawansa S, Umeno K, Takakura H, et al. Autonomic responses during inhalation of natural fragrance of Cedrol in humans. Auton Neurosci. 2003;108:79–86.
- Yada Y, Sadachi H, Nagashima Y, et al. Overseas survey of the effect of cedrol on the autonomic nervous system in three countries. J Physiol Anthropol. 2007;26:349–354.
- Schiffman SS, Sattely-Miller EA, Suggs MS, et al. The effect of pleasant odors and hormone status on mood of women at midlife. Brain Res Bull. 1995;36:19–29.
- Inoue N, Kuroda K, Sugimoto A, et al. Autonomic nervous responses according to preference for the odor of jasmine tea. Biosci Biotechnol Biochem. 2003;67:1206–1214.
- Burdock GA. Encyclopedia of food and color additives. Boca Raton: CRC Press; 1997. p. 873.
- Yada Y. The physiological analysis for the sedative effect of cedrol. Aroma Res. 2003;15: 213–223. Japanese.
- Buttery RG, Takeoka GR, Krammer GE, et al. Identification of 2,5-dimethyl-4-hydroxy-3(2H)-furanone (furaneol) and 5-methyl-4-hydroxy-3(2H)-furanone in fresh and processed tomato. Lebensm Wiss Technol. 1994;27:592–594.
- Wilhelm B, Giedke H, Lüdtke H, et al. Daytime variations in central nervous system activation measured by a pupillographic sleepiness test. J Sleep Res. 2001;10:1–7.
- Sadachi H, Nagashima Y, Tojo S, et al. Effects of cedrol on the autonomic nervous system and survey of sleep and stress in USA. J Jpn Cosmet Sci Soc. 2007;31:148–152.
- Kistler A, Mariauzouls C, Von Berlepsch K. Fingertip temperature as an indicator for sympathetic responses. Int J Psychophysiol. 1998;29:35–41.
- Or CKL, Duffy VG. Development of a facial skin temperature-based methodology for non-intrusive mental workload measurement. Occup Ergon. 2007;7:83–94.
- Lee E, Lee BJ, Ha J, et al. Efficacy of fermented green tea on peripheral skin temperature: a randomized and placebo-controlled clinical study. J Cosmet Dermatol. 2016;15:226–230.
- Fujisawa F, Nadamoto T, Fushiki T. Effect of intake of ginger on peripheral body temperature. Jpn Soc Nutr Food Sci. 2005;58:3–9.
- Zhou L, Ohata M, Owashi C, et al. Odors generated from the Maillard reaction affect autonomic nervous activity and decrease blood pressure through the olfactory system. J Sci Food Agric. 2018;98:923–927.
- Scholkmann F, Kleiser S, Metz AJ, et al. A review on continuous wave functional near-infrared spectroscopy and imaging instrumentation and methodology. Neuroimage. 2014;85:6–27.
- Aoyama S, Toshima T, Saito Y, et al. Maternal breast milk odour induces frontal lobe activation in neonates: a NIRS study. Early Hum Dev. 2010;86:541–545.
- Caldwell M, Scholkmann F, Wolf U, et al. Modelling confounding effects from extracerebral contamination and systemic factors on functional near-infrared spectroscopy. Neuroimage. 2016;143:91–105.