Abstract
Trypanosomes contain unique peroxisome-like organelles designated glycosomes which sequester enzymes involved in a variety of metabolic processes including glycolysis. We identified three ABC transporters associated with the glycosomal membrane of Trypanosoma brucei. They were designated GAT1–3 for Glycosomal ABC Transporters. These polypeptides are so-called half-ABC transporters containing only one transmembrane domain and a single nucleotide-binding domain, like their homologues of mammalian and yeast peroxisomes. The glycosomal localization was shown by immunofluorescence microscopy of trypanosomes expressing fusion constructs of the transporters with Green Fluorescent Protein. By expression of fluorescent deletion constructs, the glycosome-targeting determinant of two transporters was mapped to different fragments of their respective primary structures. Interestingly, these fragments share a short sequence motif and contain adjacent to it one – but not the same – of the predicted six transmembrane segments of the transmembrane domain. We also identified the T. brucei homologue of peroxin PEX19, which is considered to act as a chaperonin and/or receptor for cytosolically synthesized proteins destined for insertion into the peroxisomal membrane. By using a bacterial two-hybrid system, it was shown that glycosomal ABC transporter fragments containing an organelle-targeting determinant can interact with both the trypanosomatid and human PEX19, despite their low overall sequence identity. Mutated forms of human PEX19 that lost interaction with human peroxisomal membrane proteins also did not bind anymore to the T. brucei glycosomal transporter. Moreover, fragments of the glycosomal transporter were targeted to the peroxisomal membrane when expressed in mammalian cells. Together these results indicate evolutionary conservation of the glycosomal/peroxisomal membrane protein import mechanism.
| Acronyms | ||
| glycosomal ABC transporter | = | Abbreviations: GAT |
| green fluorescent protein | = | GFP |
| peroxisomal ABC transporter | = | PAT |
| phosphate-buffered saline | = | PBS |
| peroxin, PMP, peroxisome-membrane protein | = | PEX |
| peroxisome-targeting signal | = | PTS |
| transmembrane domain | = | TMD |
| transmembrane segment | = | TMS |
Introduction
Glycosomes are single membrane-bounded organelles which contain part of the glycolytic pathway. They are only found in organisms belonging to the protozoan order Kinetoplastida, such as Trypanosoma and Leishmania species Citation[1], Citation[2]. Nevertheless, glycosomes were shown to belong to the same organelle family as peroxisomes and plant glyoxysomes Citation[3]. They all contain enzymes of metabolic pathways like peroxide metabolism, fatty-acid β-oxidation or ether-lipid biosynthesis. Moreover, the mechanism of biogenesis appears largely conserved among these organelles. Several trypanosomal proteins homologous to peroxins, the proteins involved in peroxisome biogenesis, have been identified and characterized Citation[3].
The import of proteins into the matrix of peroxisomes/glycosomes has been extensively studied Citation[3–5]. Proteins are synthesized in the cytosol and recognized through their peroxisome-targeting signal (PTS) by either of two soluble receptors. Complexes of other peroxins are then responsible for the docking of the receptor loaded with the PTS-protein at the membrane, membrane translocation and receptor recycling. Fewer details are known about the mechanism by which proteins are inserted into the peroxisomal membrane, although recent publications have shed some light on different aspects of the process Citation[6–9]. Targeting determinants have been described for several membrane proteins but, contrary to the situation for matrix proteins, no consensus sequences have been definitely established Citation[10]. In some cases a cluster of basic amino acids was shown to be important Citation[11–13]. The necessity for the presence of one or more transmembrane segments was also described Citation[10], Citation[14–16]. Regarding the mechanism by which proteins are inserted into the peroxisomal membrane, only a few peroxins were shown to be essential: PEX19, PEX3 and PEX16. PEX19 is a predominantly cytosolic, partially peroxisomally-associated protein Citation[17], Citation[18] that has the specificity to interact with all peroxisomal membrane proteins (PMPs) tested so far Citation[17], Citation[19–22]. This property makes it the best candidate as PMP-import receptor. Several recent studies provided conclusive evidence that PEX19 is indeed able to stabilize PMPs in the cytosol and to subsequently target them to the peroxisomal membrane Citation[6], Citation[7], Citation[9]. Besides this main activity, other functions have also been attributed to PEX19, for example a regulatory role in the formation of membrane-associated protein complexes Citation[23]. PEX3 is thought to act as the docking factor for the PMP-loaded PEX19 at the peroxisomal membrane Citation[8]. Human PEX16 has also been shown to participate in the early steps of peroxisomal membrane assembly but its exact role remains unclear Citation[24], Citation[25]. No kinetoplastid homologues of PEX3, and PEX16 have been identified so far, but a report on T. brucei PEX19 was published while our paper was under review Citation[26].
A large amount of information is available about the metabolic processes occurring within peroxisomes, glyoxysomes and glycosomes, and about the import of proteins into the organelles. However, we have little knowledge about the transport of metabolites through the boundary membrane of these organelles and thus about the details how cytosolic and organellar metabolism are coordinated. There is considerable evidence that peroxisomal/glycosomal membranes are poorly permeable for most metabolites, implying the need for transporter molecules (reviewed in Citation[3], Citation[27]). For yeast and mammalian cells two classes of solute transporters have been identified in peroxisomal membranes. In yeast, the ANT1 ATP/ADP exchanger Citation[28] and the long-chain fatty acid transporter PAT1/PAT2 Citation[29] have been described and only ANT1 has been reconstituted in proteoliposomes. PAT1 and PAT2 are homologous proteins, belonging to the ABC-transporter superfamily. However, they are so called ‘half ABC transporters’, i.e., containing only one membrane domain and a single nucleotide-binding domain. They have been shown to act as heterodimers thus constituting a functional full-size transporter molecule Citation[29]. Four of such half ABC transporters are present in mammalian peroxisomes: the adrenoleukodystrophy protein (ALDP or ABCD1) Citation[30], the adrenoleukodystrophy-related protein (ALDRP or ABCD4) Citation[31], the 70-kDa peroxisomal membrane protein (PMP70 or ABCD3) Citation[32] and the 69-kDa peroxisomal membrane protein (PMP69, P70R or ABCD4) Citation[33]. The functions of these transporters have not been elucidated to date.
In this study, we used the sequences of known peroxisomal ABC transporters to identify the first putative metabolite transporters of T. brucei glycosomes. Three members of this family (called GAT, for glycosomal ABC transporter) could be identified and for two of them (GAT1 and GAT2) a minimal glycosome-targeting domain was determined. We also identified a putative T. brucei PEX19 that was able to interact with the targeting domain in a bacterial two-hybrid system. This interaction is also observed between trypanosome GAT2 and human PEX19, arguing in favour of evolutionary conservation of PEX19 function.
Materials and methods
Nomenclature
Slightly different systems are being used for abbreviations of proteins and genes in research on peroxisomes/glyoxysomes/glycosomes in mammals, plants, yeasts and protozoa. Throughout this paper, we have used the official nomenclature system for trypanosomatid protozoa (upper case for proteins, italicized upper case for genes) Citation[34].
Nucleotide sequences
The new nucleotide sequences reported in this paper are available in the GenBank/EMBL/DDJB databases under the accession numbers DQ065772 (TbGAT1), DQ065773 (TbGAT2) and DQ065774 (TbPEX19).
Cloning of genes and construction of plasmids for trypanosome transfections
The detailed procedures used for the cloning of the T. bruceiGAT1, GAT2, GAT3 and PEX19 genes, and for the preparation of the large number of DNA constructs coding for fusions of GAT (fragments) and Green Fluorescent Protein are available on the Internet as Supplementary Materials to this paper.
Multiple alignments of amino-acid sequences, and an analysis of the relationships of the sequences by dendogram construction were performed using the program ClustalX Citation[35].
Parasite culturing and transfection
Procyclic (insect-form) T. brucei 427 cells (cell line 449, constitutively expressing the tetracycline repressor) were cultured in normal SDM79 medium Citation[36] supplemented with 15% foetal bovine serum (Gibco/Invitrogen) and 1 µg/ml phleomycin in an incubator at 28°C under water-saturated air with 5% CO2. Transfected cells were selected with 10 µg/ml blasticidin.
Transfection was performed as described Citation[37]. Briefly, 2–3 x 107 cells were centrifuged, washed once in ice-cold Zimmerman Post-Fusion Medium and resuspended in 500 µl of this medium. 10 µg of DNA was restricted overnight with the enzyme NotI (Fermentas), precipitated with ethanol and resuspended in 20 µl of sterile water. DNA and cells were incubated together for 10 min on ice, in a 4 mm electroporation cuvette. Cells were then subjected to a single pulse by a BTX ECM 630 electroporator set for a peak discharge of 1.8 kV, 25 Ohm and 50 µF and directly diluted in 4.5 ml of SDM79 medium. Selection was applied after one day by addition of 4.5 ml of SDM79 supplemented with 10 µg/ml blasticidin.
Immunofluorescence studies
Induction of expression was done by addition of 2 µg/ml tetracycline. The trypanosomes were allowed to grow overnight before immunofluorescence analysis. Cells were fixed with 4% formaldehyde in phosphate-buffered saline (PBS), permeabilized with 1% Triton X-100 and spread on poly-L-lysine coated slides. Cells were then incubated for 45 min in PBS containing 5% BSA, followed by incubation in PBS with 2% BSA and the primary antiserum (i.e. rabbit polyclonal anti-T.brucei hexokinase as glycosomal marker). After washing with PBS, cells were allowed to react with 5 µg/ml Alexa 568 anti-rabbit IgG conjugate (Molecular Probes), washed again and mounted in Mowiol. Cells were visualized using a Zeiss Axiovert microscope coupled to an MRC-1024 confocal scanning laser imaging system (BioRad).
Two-hybrid analysis
The yeast and bacterial two hybrid constructs coding for human peroxin fusion proteins have been described elsewhere Citation[21], Citation[22]. Yeast two-hybrid plasmids encoding GAL4BD or GAL4AD fused to fragments encoding T. brucei GAT1(1–207), GAT1(74–207), GAT1(116–207), GAT2(1–90), GAT2(1–126) and GAT2(68–126) were prepared by amplification of the corresponding sequences with oligonucleotides adding EcoRI and BamHI restriction sites. Amplified DNA was inserted into the pGBT9 and pGAD424 vectors (Clontech). The inserts of the bacterial two-hybrid plasmids were also generated by PCR (templates: the corresponding yeast two-hybrid plasmids; primers: 5′-GGGCGCGCGGCCGCCATAAGAAATTCGCCCGGAAT-3′ and 5′-GGGCTCGAGGTTGACTGTATCGCCGGAATTC-3′). The amplified DNA fragments restricted by XhoI/NotI were subsequently ligated to the modified bacterial two-hybrid vectors pMF413 (restricted by XhoI/NotI) and pMF424 (restricted by SalI/NotI), whose constructions have been described elsewhere Citation[21]. The construction of the HsPEX19 pentapeptide mutant library has been described by Fransen et al. Citation[38]. Bacterial (E. coli strain BTH101) and yeast (S. cerevisiae strain SFY526) two-hybrid analyses were performed as described elsewhere Citation[21], Citation[22].
Expression of TbPEX19 in mammalian cells
The complete TbPEX19 sequence was amplified using oligonucleotides which added EcoRI and KpnI restriction sites at the termini of the gene. The PCR product was treated with these restriction enzymes and subsequently ligated to a EcoRI/KpnI-digested pEGFP-C1 vector (Clontech). Pex5+/- mouse fibroblasts were cultured as described Citation[39], Citation[40]. After transfer to coverslips, the cells were transiently transfected with the plasmid encoding the GFP-TbPEX19 fusion protein by employing the Magnetofection transfection methodology (OZ Biosciences), and processed for direct fluorescence as described Citation[39]. Human PEX19-deficient fibroblasts Citation[18], obtained from Prof. R. Wanders (University of Amsterdam), were transiently transfected with the GFP-TbPEX19 plasmid using the lipofectamine™ Plus Reagent (Invitrogen). The peroxisomal localization of the GFP-fusion proteins was confirmed by co-localization studies with the peroxisome-targeted DsRed-SKL reporter protein Citation[22]. Fluorescence was observed under a Leica DMR microscope equipped with FITC/RSGFP/Bodipy/Fluo3/DIO and Texas red filters. Chinese Hamster Ovary (CHO) cells were cultured, transiently transfected, and processed for indirect immunofluorescence as described elsewhere Citation[22].
Results
Cloning and sequence analysis of ABC transporter genes
Searches were performed in the database of the T. brucei (strain TREU947/4) genome project to identify proteins homologous to known peroxisomal transporters. Sequences derived from overlapping sheared DNA fragments enabled us to assemble parts of genes for two different putative glycosomal ABC transporters. These proteins were named GAT1 and GAT2 for Glycosomal ABC Transporter 1 and 2, respectively. Clones of the two genes were isolated from a T. brucei (strain 427) genomic library and sequenced (). The encoded GAT1 and GAT2 amino-acid sequences are 683 and 641 residues long, corresponding to proteins with a calculated Mr of 75978 and 71247 Da, respectively. The proteins have a predicted structural organization similar to that reported for other peroxisomal ABC transporters: a transmembrane domain (TMD) fused to a nucleotide-binding domain containing canonical Walker A and B motifs as well as the ABC signature. The overall identity between trypanosomal and human, yeast or plant homologues is around 25%. If only the ABC domains are compared, the identity increases to about 40%. By contrast, the identity of the TMD is less than 20%. Despite this low degree of similarity between the glycosomal and peroxisomal ABC transporter TMDs, a relatively well-conserved motif could be recognized comprising the second predicted transmembrane segment (TMS) as well as the sequence just in front of it (). This region has already been postulated to play a role in the targeting of the mammalian ALDP protein to the peroxisomal membrane Citation[41].
Figure 1. Alignment of the amino-acid sequences of kinetoplastid glycosomal ABC transporters. Sequences of putative Leishmania major (Lm) and Trypanosoma cruzi (Tc) orthologues were found by homology searches in the GeneDB database, using the T. brucei (Tb) sequence as a query. LmGAT1 (GeneDB systematic name LmjF31.0540), LmGAT2 (LmjF33.1860), LmGAT3 (LmjF27.0470), TcGAT1 (Tc00.1047053509237.30), TcGAT2 (Tc00.1047053510431.150) and TcGAT3 (Tc00.1047053506925.530). The sequences were aligned using the ClustalW 1.8 program (http://searchlauncher.bcm.tmc.edu/multi-align/Options/clustalw.html). Identical amino acids are given against a black background, conserved residues (similar in more than 50% of the sequences) against grey shading. Hatched bars above the alignment represent approximate positions of predicted TMSs as identified in GAT1 and GAT2 sequences using prediction programs available on the ExPASy server (http://www.expasy.org): TMHMM, HMMTOP and TopPred. Amino acids in bold-italic correspond to the region essential for targeting as revealed by deletion analysis (see text). The conserved RVVGLL motif identified in these regions is underlined. Boxed regions represent positions of the predicted PEX19-binding sites identified using the S. cerevisiae prediction matrix developed by Rottensteiner et al. Citation[46]. Asterisks indicate the region suggested by Landgraf et al. Citation[41] to bear targeting determinants of HsALDP.
![Figure 1. Alignment of the amino-acid sequences of kinetoplastid glycosomal ABC transporters. Sequences of putative Leishmania major (Lm) and Trypanosoma cruzi (Tc) orthologues were found by homology searches in the GeneDB database, using the T. brucei (Tb) sequence as a query. LmGAT1 (GeneDB systematic name LmjF31.0540), LmGAT2 (LmjF33.1860), LmGAT3 (LmjF27.0470), TcGAT1 (Tc00.1047053509237.30), TcGAT2 (Tc00.1047053510431.150) and TcGAT3 (Tc00.1047053506925.530). The sequences were aligned using the ClustalW 1.8 program (http://searchlauncher.bcm.tmc.edu/multi-align/Options/clustalw.html). Identical amino acids are given against a black background, conserved residues (similar in more than 50% of the sequences) against grey shading. Hatched bars above the alignment represent approximate positions of predicted TMSs as identified in GAT1 and GAT2 sequences using prediction programs available on the ExPASy server (http://www.expasy.org): TMHMM, HMMTOP and TopPred. Amino acids in bold-italic correspond to the region essential for targeting as revealed by deletion analysis (see text). The conserved RVVGLL motif identified in these regions is underlined. Boxed regions represent positions of the predicted PEX19-binding sites identified using the S. cerevisiae prediction matrix developed by Rottensteiner et al. Citation[46]. Asterisks indicate the region suggested by Landgraf et al. Citation[41] to bear targeting determinants of HsALDP.](/cms/asset/6e25077e-0b8a-490a-b7f1-9d1d5ba42a76/imbc_a_145995_f0001_b.gif)
As all peroxisomal ABC transporters, both GATs were predicted to have six TMSs (). However, some significant differences were found with regard to their positions. The sixth TMS usually proposed for peroxisomal ABC transporters was not observed in any of the predictions for GATs. In contrast, a strongly predicted TMS was found at the extreme N-terminal end of the two Trypanosoma proteins.
While this work was ongoing, the trypanosomatid genome sequencing projects reached almost completion and the full set of sequence data became available through the GeneDB database. New careful searches led to the surprising finding of a third homologous putative glycosomal ABC transporter, called GAT3. The T. brucei 427 gene was therefore also cloned and analyzed. The overall identity of GAT3 with peroxisomal homologues is even lower than it is for GAT1 and GAT2. TMD identity drops in this case to around 15%. Moreover several unique insertions were observed, some of them found in the very well conserved nucleotide-binding domain of the proteins (see ).
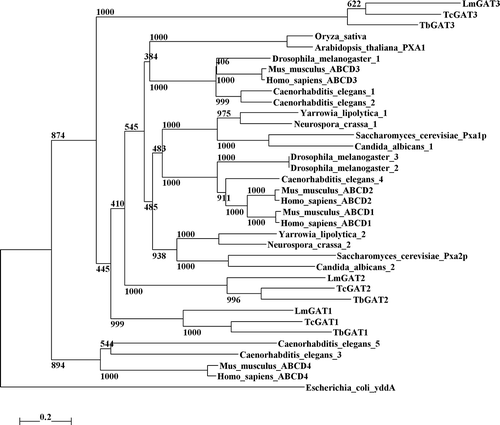
Putative orthologous of the three T. brucei GAT sequences were found in the databases of the related organisms Leishmania major and Trypanosoma cruzi (). Details about the similarities between the encoded proteins and a dendogram showing the relationships among them and with peroxisomal ABC transporters are presented as Supplementary Material online.
Subcellular localization
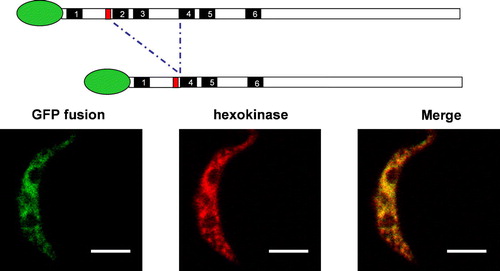
Subcellular targeting of the three GAT proteins was investigated by confocal microscopy using GFP-fusion constructs. For that purpose, GFP-expression plasmids were developed for stable integration into the trypanosome genome. Expression was assessed by fluorescence emission. Subsequently, cell lines were constructed expressing the full-length GAT proteins fused either N- or C-terminally to the GFP and the subcellular localization of the fluorescence was analysed. In all six cases, a discrete punctuate emission pattern was observed (). This pattern was completely superimposable with that observed for the glycosomal enzyme hexokinase.
Figure 2. Subcellular localization of GAT1, GAT2, and GAT3 variants. Procyclic trypanosomes were stably transfected with chromosomally-integrated plasmids designed for inducible expression of GFP-fusion proteins. After overnight induction, cells were processed for immunofluorescence with a polyclonal anti-hexokinase antibody as glycosomal marker. Full-length GAT1, full-length GAT3, full-length GAT2, GAT2(1–270) and GAT2(1–126) showed a punctuate fluorescence staining pattern completely colocalizing with hexokinase and clearly different from the pattern observed for GFP alone. GAT1(1–132) is mislocalized to the cytoplasm. In trypanosomes transfected with GAT2(1–270) only one or two enlarged glycosomes or aggregates of the organelles per trypanosome could be seen. The resolution did not permit an unambiguous distinction between these two possibilities, but aggregation seems most likely. Organelle aggregation has previously been described for the filamentous fungus Podospora anserina when its peroxisomal ABC transporter was overexpressed Citation[55]. Bars, 5 µm.
![Figure 2. Subcellular localization of GAT1, GAT2, and GAT3 variants. Procyclic trypanosomes were stably transfected with chromosomally-integrated plasmids designed for inducible expression of GFP-fusion proteins. After overnight induction, cells were processed for immunofluorescence with a polyclonal anti-hexokinase antibody as glycosomal marker. Full-length GAT1, full-length GAT3, full-length GAT2, GAT2(1–270) and GAT2(1–126) showed a punctuate fluorescence staining pattern completely colocalizing with hexokinase and clearly different from the pattern observed for GFP alone. GAT1(1–132) is mislocalized to the cytoplasm. In trypanosomes transfected with GAT2(1–270) only one or two enlarged glycosomes or aggregates of the organelles per trypanosome could be seen. The resolution did not permit an unambiguous distinction between these two possibilities, but aggregation seems most likely. Organelle aggregation has previously been described for the filamentous fungus Podospora anserina when its peroxisomal ABC transporter was overexpressed Citation[55]. Bars, 5 µm.](/cms/asset/9de92341-9db2-4f00-b3a8-c86b7edb307c/imbc_a_145995_f0002_b.jpg)
Targeting determinants of the GAT proteins
To identify determinants required to target proteins to the T. brucei glycosomal membrane, we selected various corresponding fragments of GAT1 and GAT2 based on a multiple sequence alignment, and expressed them as GFP-fusion constructs in trypanosomes. The localization phenotypes are summarized in .
Figure 3. Schematic representation of T. brucei deletion mutant constructs and their glycosomal localization. The ellipse indicates the position of the GFP and the black boxes represent the positions of the predicted transmembrane segments. The subcellular localization as concluded from fluorescence is indicated as follows:+complete glycosomal localization;±partial glycosomal localisation; Cytopl. Cytoplasmic localization;-absence of detectable fluorescence.
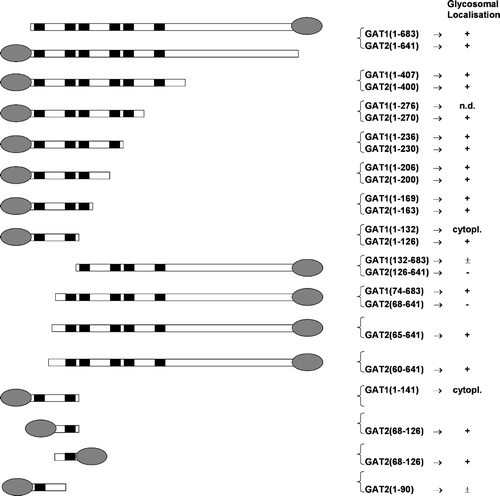
First, deletion of the C-terminal half containing the nucleotide-binding domain of the proteins had no effect on targeting efficiency, showing that important information for glycosomal localization is present within the N-terminal half containing the integral membrane part of the GATs. Additional constructs were designed, by progressively shortening the TMD of each GAT from the C- to the N-terminal side. Deleting the sixth TMS and the loop upstream of it did not affect the targeting efficiency [construct GFP-GAT2(1–270)]. All successive deletion constructs retained a perfect glycosomal localization until the third predicted TMS was reached. A different GFP localization pattern was obtained when this TMS was deleted from either GAT1 or GAT2. Proper localization was only retained in the case of GFP-GAT2(1–126). For the equivalent GAT1 deletion, GFP-GAT1(1–132), we observed a shift of the fluorescence to the cytoplasmic compartment. To confirm that the fluorescence shift was not due to a folding problem, we also expressed a slightly longer fragment of GAT1, GFP-GAT1(1–141). This fragment too retained a cytoplasmic localization, showing that the targeting determinants of the two proteins are not present within the corresponding region of their aligned sequences.
To confirm the presence of a single targeting signal within each GAT protein, we also made N-terminal deletions with GFP fused at the C-terminus. The construct GAT1(132–683)-GFP appeared to be partially located in the cytoplasm and partially associated with glycosomes. The GAT2(126–683)-GFP protein was not detectable at all under those conditions; it may have been unstable and thus rapidly degraded. A shorter N-terminal deletion led to a complete glycosomal localisation for GAT1 [GAT1(74–683)-GFP], but the corresponding GAT2 fragment remained undetectable. We therefore designed two additional GAT2 deletions [GAT2(65–641)-GFP and GAT2(60–641)-GFP)]. These two constructs were detected and colocalized perfectly with the glycosomal marker.
It should be noted that two N-terminal deletion constructs with GFP fused at the N-terminal side, GFP-GAT1(132–683) and GFP-GAT2(126–641), showed a cytoplasmic expression pattern (not shown). In view of the position of the routing determinants as located in all experiments described above, this was expected for GAT2, but was rather surprising for GAT1. A possible explanation is that the presence of the GFP protein just upstream of the routing signal interferes with the efficiency of targeting.
The GAT2 targeting region was further narrowed down by designing a combination of N- and C-terminal truncations, leading to constructs GFP-GAT2(68–126) and GAT2(68–126)-GFP. Both recombinant proteins were well expressed and showed a complete glycosomal localization. They contain the GAT2 fragment with TMS2 and the relatively well-conserved motif in front of it. To determine whether or not the second predicted TMS (residues 92–109) itself was important for targeting, we expressed the fragment GFP-GAT2(1–90). This construct retained its glycosomal localization but was also partially found in the cytoplasm.
Summarized, our data suggest that the amino acids 132–169 of GAT1 and 68–90 of GAT2 are essential for glycosome targeting. However, note that we currently can not exclude the possibility that GAT1 and GAT2 variants lacking these amino acids are defective in protein folding and/or have impaired assembly into the membrane. When comparing the primary structure of the two regions responsible for targeting of T. brucei GAT 1 and 2, the only apparent similarity consists in a block of six conserved amino acids, RVVGLL. However, this block was not found in any other related sequence presented in .
We finally analyzed the importance of the TMS2 sequence for the targeting efficiency by making an internal deletion. The result obtained with this construct demonstrated convincingly that the TMS is not required for the targeting itself of GAT2, but rather for its correct insertion into the glycosomal membrane (data shown in Supplementary Material online).
Identification of a T. brucei PEX19 homologue
To further elucidate the mechanism by which the glycosomal ABC transporters GAT1 and GAT2 are inserted into the glycosomal membrane, we pursued our research with the identification of proteins that bind the domains essential for glycosomal localization. For the glycosome-related organelle, the peroxisome, evidence has recently been provided that a single protein, PEX19, acts as a chaperone and import receptor for newly synthesized PMPs Citation[6–19]. We therefore performed searches in the genome project databases to identify a putative T. brucei homologue. A good candidate was found with orthologues in L. major and T. cruzi (). The TbPEX19 found in the database for strain TREU927/4 has a calculated Mr of 31413 Da and shows 25 and 35% identity with its L. major and T. cruzi homologues, respectively. The identity drops to 15% and 16%, respectively, for the human and Saccharomyces cerevisiae PEX19. The PEX19 gene of T. brucei strain 427 was amplified, cloned and sequenced. It shows eight nucleotide changes compared to the database TbPEX19, resulting in four amino-acid substitutions (GenBank accession number DQ065774).
Figure 4. Alignment of kinetoplastid putative PEX19s with human and yeast homologues. TbPEX19 (GeneDB systematic name Tb09.211.3300), LmPEX19 (LmjF35.3260) and TcPEX19 (Tc00.1047053510737.100) were identified in the GeneDB database by homology searches with human and yeast sequences. Identical amino acids are given against a black background, conserved residues (similar in more than 50% of the sequences) against grey shading.

Human PEX19 has been shown to interact with several PMPs including HsPEX3, HsPEX11β, HsPEX12, HsPEX13, HsPEX14, HsPEX16, and HsPEX26 in yeast and/or bacterial two-hybrid systems. We were interested in determining if any of these interactions would also be found when replacing HsPEX19 by its T. brucei homologue. By employing the bacterial (s in Supplementary Material online) and yeast (data not shown) two-hybrid systems, we could show that TbPEX19p can interact with HsPEX3. Interactions of TbPEX19 with other human PMPs could not be demonstrated (data not shown). The strong interaction between human PEX3 and trypanosome PEX19 offered a first good indication that the cloned gene is indeed an authentic, functional PEX19. To analyze this further, we expressed GFP-TbPEX19 in mammalian cells. Transfected cells showed a fluorescence staining pattern varying from completely cytoplasmic to partially cytoplasmic, partially peroxisomal (). This phenotype is similar to what was reported for human PEX19 Citation[17], Citation[18]. We therefore conclude that TbPEX19, when expressed in a heterologous system such as human cells, is able to dock at the peroxisomal membrane. Although no interactions were observed between TbPEX19 and several other human peroxins (see above), we also investigated whether or not TbPEX19 was able to restore peroxisome biogenesis in a PEX19-deficient human cell line. As expected this appeared not to be the case (data not shown).
Figure 5. Intracellular localization of TbPEX19 in mouse fibroblasts. Mouse fibroblasts transfected transiently with plasmids expressing the peroxisomal marker protein DsRed-SKL and GFP-TbPEX19 were examined for direct fluorescence 48 h after transfection. The two extreme observed phenotypes are presented. (A) TbPEX19 is found exclusively in the cytoplasm. (B) TbPEX19 is partially cytoplasmic and partially peroxisomal as shown by its colocalization with DsRed-SKL. Bars, 10 µm.
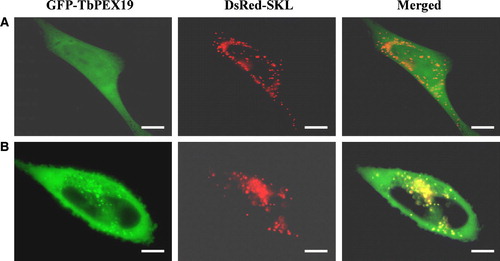
Table I. Binding properties of GAT1 and GAT2 fragments to human and trypanosomal PEX19s
Interaction between PEX19 and glycosomal ABC transporters
PEX19 is thought to act both as a chaperone for newly synthesized PMPs and also as a receptor for their membrane insertion. Indeed, this peroxin has been shown to interact with a considerable number of PMPs in several organisms Citation[17], Citation[19–22]. We therefore checked if the putative TbPEX19 was also able to bind to the PMPs GAT1 and GAT2, and if that would be the case, whether or not the PEX19 binding-regions of the GATs correlated with the regions essential for glycosomal localization. To this end, fragments GAT1(1–207), GAT1(74–207), GAT1(116–207), GAT2(1–126), GAT2(1–90) and GAT2(68–126) were analyzed for binding to TbPEX19 as well as HsPEX19 in the bacterial two-hybrid system. β-Galactosidase-based two-hybrid assays were performed with liquid cultures and the results are given in .
All GAT2 deletion constructs, except T25-GAT2(1–90), showed a similar, considerable level of binding to trypanosome PEX19, but also to its human homologue. The apparent lack of binding of construct T25-GAT2(1–90) to T18-TbPEX19, contrary to the situation observed with the reciprocal constructs and its binding to T18-HsPEX19 is at present not understood. Concerning GAT1, construct GAT1(1–207) showed only a low extent of binding when fused to the T25 part of the system, whereas GAT1(74–207), GAT1(116–207) showed a stronger affinity for both HsPEX19 and TbPEX19. This should, most likely, be attributed to the fact that GAT1(1–207) contains three putative TMSs that may interfere with detection of interactions in two-hybrid experiments Citation[42]. The shorter GAT1 constructs displaying a stronger affinity for both PEX19s, contain either only one or two TMSs. Together, these results indicate that: (i) TbPEX19 is able to interact with fragments of the trypanosome PMPs GAT1 and GAT2 which contain the glycosomal localization determinant, and (ii) the PMP–PEX19-binding is evolutionary conserved between trypanosomes and human, as the GAT signals are recognized by both PEX19s.
Human PEX19 has been shown to consist of a C-terminal core domain and a flexible N-terminal region Citation[9]. In addition, the first 50 amino acids of the protein have been shown to be necessary for interaction with the PEX3 protein of several organisms Citation[43–45]. Recently, pentapeptide scanning mutagenesis studies also revealed the presence of a PEX14-binding box in the N-terminal part of human PEX19 Citation[38]. These studies also showed that the C-terminal core region of the protein is required for binding to other PMPs such as PEX11β, PEX12, PEX13, PEX16 and PEX26. In view of the conserved interaction observed between human PEX19 and trypanosomal GATs, we were interested to see which pentapeptide mutants of HsPEX19 would be affected in their binding affinity to GAT2. The complete set of available HsPEX19 mutants (87 independent clones) was tested for interaction with constructs GAT2(1–126) and GAT2(68–126). Results obtained for GAT2(68–126) are presented in . Similar results were obtained with GAT2(1–126), except for differences seen for insertions at the positions 190, 192 and 258 (data not shown). In both cases, all affected mutants clustered in the C-terminal core domain. GAT2 is the first non-peroxin PMP tested against this HsPEX19 mutant library.
Figure 6. Schematic representation of the binding capacity of human PEX19 variants containing random peptide insertions to TbGAT2(68–126), as observed using a bacterial two-hybrid system. The amino-acid sequence of human PEX19 is shown. The pentapeptide insertions are indicated by vertical arrows, and named by the position number of the amino-acid residue to the N-terminal side of the insertion followed by the sequence of the insertion. Pentapeptide insertions retaining the same binding properties as wild-type PEX19 are indicated with normal characters. Insertions resulting in a strongly diminished binding affinity for GAT2(68–126) are given in italic and are marked with a dot. Insertions that resulted in a diminished binding affinity for GAT2(68–126) but retained a strong affinity for the human membrane peroxins HsPEX11β, HsPEX16 and HsPEX26 Citation[38] are in normal characters and marked by underlining. Boxed, grey-shaded regions indicate α-helices as predicted by the PSA-server (probability R0.85).
![Figure 6. Schematic representation of the binding capacity of human PEX19 variants containing random peptide insertions to TbGAT2(68–126), as observed using a bacterial two-hybrid system. The amino-acid sequence of human PEX19 is shown. The pentapeptide insertions are indicated by vertical arrows, and named by the position number of the amino-acid residue to the N-terminal side of the insertion followed by the sequence of the insertion. Pentapeptide insertions retaining the same binding properties as wild-type PEX19 are indicated with normal characters. Insertions resulting in a strongly diminished binding affinity for GAT2(68–126) are given in italic and are marked with a dot. Insertions that resulted in a diminished binding affinity for GAT2(68–126) but retained a strong affinity for the human membrane peroxins HsPEX11β, HsPEX16 and HsPEX26 Citation[38] are in normal characters and marked by underlining. Boxed, grey-shaded regions indicate α-helices as predicted by the PSA-server (probability R0.85).](/cms/asset/82b37389-93a4-4e0e-b72e-a2ced6386c40/imbc_a_145995_f0006_b.gif)
To confirm that this observed heterologous interaction is indeed relevant for the understanding of the glycosome assembly mechanism in T. brucei, TbPEX19 was separated in two parts, TbPEX19(1–60) and TbPEX19(61–285), that were tested for interaction with GAT2(1–126) in the bacterial two-hybrid system. As expected, based on studies with human PEX19 Citation[38], only the C-terminal domain of PEX19 showed a significant level of binding. On the contrary, only the N-terminal half of TbPEX19 presented a very strong affinity for human PEX3 (see s in Supplementary Material online). Our results thus confirm the role of the C-terminal domain of PEX19 in binding and probably stabilizing a number of distinct PMPs in the cytoplasm. This binding capacity is not only retained between PMPs from the same organism but is clearly evolutionarily conserved, again arguing in favour of a crucial role for PEX19 in the biogenesis of the peroxisomal/glycosomal membrane.
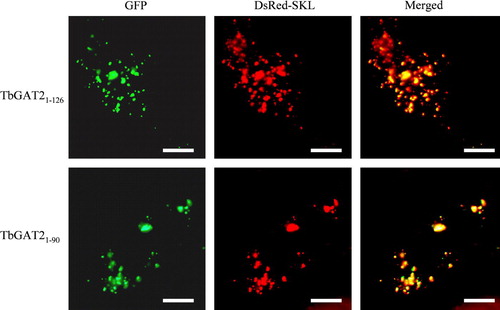
To determine whether or not the ability of HsPEX19 to bind to the TbGAT2 constructs containing the glycosome-targeting determinants would also result in their routing to peroxisomes in vivo in mammalian cells, we expressed GFP-fusions of the fragments GAT2(1–126) and GAT2(1–90) in CHO cells (). A peroxisomal localization was clearly demonstrated by their colocalization with the DsRed-SKL marker.
Figure 7. Intracellular localization of GAT2(1–126) and GAT2(1–90) in CHO cells. CHO cells transfected transiently with plasmids expressing the peroxisomal marker protein DsRed-SKL and GFP-GAT2(1–126) or GFP-GAT2(1–90) were examined for direct fluorescence 48 h after transfection. In both cases the peroxisomal localization was demonstrated by the perfect colocalization with DsRed-SKL. Bars, 10 µm.
Recently the PEX19-binding sites of two S. cerevisiae PMPs (PEX13 and PEX11) could be determined and characterized by using synthetic peptide scan analyses Citation[46]. A prediction matrix was developed that enabled the in silico identification of PEX19-binding sites in other PMPs. In view of the observed conservation of the PEX19 binding affinity between trypanosome and human, we applied the S. cerevisiae prediction matrix to the trypanosomal GAT sequences. Several putative PEX19-binding peptides could be detected for each protein, all clustering in the N-terminal half (see ). Among these, two are in complete agreement with those obtained in the experimental localization analysis: GAT1(132–146) and GAT2(68–82). This provides another argument in favour of a conservation of the PMP insertion mechanism during evolution.
Discussion
The compartmentalization of the major part of the glycolytic pathway within glycosomes is a characteristic of all protozoa belonging to the order Kinetoplastida. If the origin and specific role of this unique compartmentation are, at least to some extent, a matter of conjecture Citation[2], Citation[47], Citation[48], the absolute requirement for proper enzyme localization inside the organelles has been experimentally proven Citation[49–51]. Nevertheless, an important gap in our comprehension of how glycosomes function in the overall cellular metabolism concerns the mechanism by which metabolites are translocated across the membrane of the organelle. Such questions concerning the translocation of metabolites do not only pertain to glycosomes, but also to the related peroxisomes. However, until now only two different peroxisomal metabolite transport systems have been characterized in some detail. These are the yeast hetero-dimeric PAT1/PAT2 transporter for long-chain fatty acids Citation[29] and the yeast (PMP47 or ANT1) Citation[28] and human (PMP34) Citation[52] ATP/ADP exchangers.
In this study, we used the sequences of known yeast and human peroxisomal ABC transporters to identify T. brucei orthologues as putative transporters in the glycosomal membrane. In fine, three homologous proteins were identified for which the glycosomal localization was confirmed by immunofluorescence. This family of three proteins is different from what has been observed in other phyla. Four family members were found in peroxisomal membranes of mammalian cells and only two in yeast peroxisomes. In the latter case an exclusive requirement for the assembly of a heterodimeric complex has been demonstrated Citation[29]. No function can be inferred for the T. brucei polypeptides merely on the basis of a primary structure analysis. Yet, preliminary complementation experiments with yeast mutants suggest that GAT1 and GAT2 are involved in fatty-acid transport (CY, PM and B. Distel – University of Amsterdam, unpublished). The fact that the third protein (GAT3) harbours specific insertions and seems to have evolved more rapidly than any other related glycosomal/peroxisomal ABC transporter, makes it of special interest. In fact, the variety of metabolic capacities of glycosomes Citation[2], Citation[53] requires the presence of multiple specific and still undiscovered metabolite transporters in the membrane, and it would not be surprising if those would have arisen from pre-existing proteins.
The import of peroxisomal matrix proteins relies on an evolutionarily conserved mechanism involving two distinct, well-defined PTS1 and PTS2 consensus sequences. Despite numerous attempts in recent years to identify a similar targeting signal for PMPs, no consensus information could be definitely established. The targeting determinants seem to vary from one protein to the other: a single or multiple signals, the presence of a cluster of basic residues and one or several transmembrane segments Citation[10–13], Citation[15], Citation[16]. To shed some more light in this field, we decided to take advantage of working with two different but closely related proteins. This enabled us to specify similar deletions based on a multiple alignment with the objective to identify critical determinants affecting localisation. Surprisingly, the targeting regions that were thus identified in GAT1 and GAT2 did not match at all when the sequences were aligned. The GAT1 targeting signal was located in the region encompassing amino acids 132–169, while the GAT2 signal was found between amino acids 68 and 90. At first sight, the only common feature that can be distinguished is a conserved six amino-acid block: RVVGLL. The specific relevance of this motif will be subject of future investigation. Furthermore, a single predicted TMS was present in the GAT fragments containing the targeting determinant, TMS3 for GAT1 and TMS2 for GAT2. Deletion studies with GAT2 in which TMS2 was removed while bringing TMS4 adjacent to the targeting motif indicates the importance of a TMS for the insertion of the protein into the membrane rather than for its subcellular targeting.
Recently, convincing evidence has been put forward for a crucial role of PEX19 in the mechanism by which PMPs are inserted into the peroxisomal membrane Citation[6], Citation[7], Citation[9], Citation[46]. This peroxin would act as both a chaperone and import receptor for newly synthesized proteins. We were interested to determine if the same mechanism operates in trypanosomes and if the minimal glycosome-targeting regions that we delineated correspond to putative PEX19 binding areas. Candidate trypanosomatid PEX19s were found in the databases by homology searches. Strong support for the identity of the T. brucei protein as a PEX19 was provided by showing that it has the ability to strongly interact with HsPEX3 in yeast and bacterial two-hybrid systems, as well as to dock at the peroxisomal membrane of mouse fibroblasts. Moreover, the TbPEX19 identity is further confirmed by results published by Banerjee and colleagues while our submitted paper was under review Citation[26]. The ability of TbPEX19 to interact with the shortest GAT1 and GAT2 fragments with glycosome-targeting information was investigated using a non-transcription based bacterial two-hybrid system. A substantial extent of interaction was detected in all cases except for fragment GAT1(1–207), but this negative result is most probably due to the presence of three TMSs. It has already previously been observed that the presence of TMSs may interfere with the interaction detection in classical two-hybrid systems Citation[42]. Interestingly, our experiments clearly showed that all GAT1 and GAT2 fragments with the targeting motif not only interacted with TbPEX19 but also with HsPEX19, indicating an evolutionary conservation of the PEX19 binding capacity.
The functional domains of human PEX19 have recently been analysed in great detail using a large pentapeptide mutant library of the protein Citation[38]. This analysis permitted the identification of three main domains involved in interactions: a N-terminal domain strongly interacting with PEX3, followed by a previously uncharacterized PEX14-binding motif and a C-terminal region constituting the binding domain for several distinct PMPs. To characterize in more detail the binding detected between human PEX19 and the glycosomal ABC transporters, we tested the interaction of fragments GAT2(1–126) and GAT2(68–126) with the whole panel of available HsPEX19 pentapeptide variants. The results obtained for the two GAT2 fragments were almost identical, and similar to those previously obtained for human PMPs. This indicates that the binding domains of the interacting partners, TbPEX19 and PMPs, as revealed in our analysis may indeed have biological relevance. This notion is reinforced by the observation that GAT2(1–126) and GAT2(1–90), upon their expression in mammalian cells, were perfectly targeted to the peroxisomes. The additional conclusion drawn from these data, i.e., the evolutionary conservation of PEX19 function, was corroborated by the in silico identification of several putative PEX19-binding sites in GATs using a prediction matrix developed for S. cerevisiae PMPs. In fact, two of such sites predicted for GAT1 and GAT2 correspond completely with those revealed by the experimental studies on GAT localisation and PEX19 binding. Very recently, and independently of our work, a conservation of PEX19 function in PMP targeting has also been concluded from studies with yeast and human peroxisomes Citation[54].
In this paper we presented the first characterization of solute transporters of the glycosomal membrane of T. brucei. We anticipate that the information obtained in this study about targeting and membrane insertion of the GATs, together with the fact that the genome sequencing projects of various trypanosomatids have been essentially completed, will facilitate the discovery and characterization of other solute carriers as well. Notably, information about glycosomal transporters of glycolytic intermediates will be essential for getting a better understanding about the precise functioning of these organelles in energy and carbohydrate metabolism of these parasites.
Supplementary material
Cloning of genes
The database of the T. brucei (strain TREU927/4) genome project, when still at a stage of partial completion, was searched for sequences homologous to yeast and mammalian peroxisomal ABC transporters. Fragments obtained by DNA shearing with sequences of two different putative candidates were identified. The corresponding fragments were amplified from genomic DNA of T. brucei strain 427, cloned using the vector pGEM-T (Promega) and used as hybridization probes to screen a T. brucei 427 genomic library in λGEM11. Positive phage clones were analysed by DNA restriction followed by Southern blot analysis. A 5 kb EcoRI fragment containing the GAT1 gene and a 9 kb SacI fragment containing GAT2 were then subcloned into plasmid pZErO-2 (Invitrogen). Both strands of the two genes were sequenced using a Beckman CEQ2000 sequencer.
Full-length GAT3 and PEX19 sequences were found in the completed GeneDB database (http://www.genedb.org) by homology searches. The accession codes are Tb11.03.0030 and Tb09.211.3300, respectively. Both genes were amplified from T. brucei 427 genomic DNA. The GAT3 gene was amplified with primers 5′-GGGCCCCATGACAACGCATTGGGTTCGTGAC-3′ (sense) and 5′-GGATCCCCGAAACCCCTGTGCAAATCAACTC-3′ (antisense), partially sequenced and subcloned in the T. brucei localisation vectors (see below). PEX19 was amplified with 5′-AGGGCTCGAGGGAATTCATGTCTCATCCCGACAATGAC-3′ and 5′-GGAGCTCGAGGTACCCTACACTGATGGTTGCACATCGGC-3′ as sense and antisense primers, respectively, and inserted into the yeast and bacterial two-hybrid vectors pGAD424 (y-2HS, Clontech), pKT25 (b-2HS, Hybrigenics), and pUT18C (b-2HS, Hybrigenics). The inserts of the pGAD424 and pKT25-derived plasmids were sequenced (Agowa).
Construction of plasmids for trypanosome transfection
Vector pHD1336 (a gift by Prof. C. Clayton, Heidelberg) was used as starting material for the construction of a plasmid suitable for the expression of Green Fluorescent Protein (GFP) -fusion proteins in trypanosomes. It contains the T. brucei PARP promoter under the control of the Escherichia coli tetracycline operator, 5′ and 3′ UTRs of a T. brucei actin gene, and the blasticidin resistance gene. The complete ORF of GFP combined with the adjacent multiple cloning site region was amplified by PCR from the plasmids pEGFP-N1 and pEGFP-C1 (Clontech). Plasmid pHD1336 was doubly restricted by enzymes HindIII and BamHI and the 5′ overhanging ends were filled by incubation with Taq DNA polymerase (TaKaRa) in appropriate buffer supplied with 0.2 mM dNTPs. The resulting blunt-end vectors were used for ligation with the amplified GFP fragments, resulting in the formation of the plasmids pGC1 and pGN1, with the possibility to insert additional DNA fragments in the still available BamHI and HindIII restriction sites to make genes coding for respectively C- and N-terminal fusion constructs of GFP.
Genes encoding full-length proteins were amplified using primers adding restriction sites ApaI and BamHI for GAT1 and restriction sites HindIII and BamHI for GAT2. The corresponding fragments were first cloned using vector pGEM-T (Promega), sequenced and subsequently inserted into the plasmids pGC1 and pGN1. C- and N-terminal deletion constructs of GATs were made by PCR amplification with specific oligonucleotides adding restriction sites ApaI and BamHI for GAT1 and HindIII and BamHI for GAT2. PCR was performed using a Pfu DNA polymerase (Fermentas), the fragments were cloned in vector pGEM-T and sequenced. Fragments with correct sequences were then transferred to the pGC1 or pGN1 GFP-fusion construct expression vectors. Constructs coding for proteins with internal deletions, GFP-GAT1(Δ97–209) and GFP-GAT2(Δ91–203), were made using the QuickChange™site Directed Mutagenesis Kit (Stratagene) according to the manufacturer's instructions. GAT1 primers used were 5′-GTCCATTCGTTCACGGGAATCCAGAAACGTCGCGGGG-3′ and its exact complement. GAT2 primers used were 5′-CAGTGTTTTCTCCCCTGAAAGCAGAGAACTTGCGGTTTTG-3′ and its exact complement.
Comparison of the GAT sequences from T. brucei, T. cruzi and L. major
The percentage of amino-acid identity between the orthologues sequences varies between 40 and 55%. By contrast, the identity between the GAT1 and GAT2 proteins within each parasite species is about 27–28%, very similar to the values observed (25–30%) when comparing trypanosomatid GAT sequences with their homologues of mammalian, plant and yeast peroxisomes. The GAT3 sequences are less conserved and have only about 15% identity with the homologues of peroxisomes. The prediction of six transmembrane segments (see in paper) and their positions in all GAT proteins was rather similar.
A dendrogram showing the relationships between amino-acid sequences of described and putative peroxisomal ABC transporters is shown in . The tree shows that the GAT3 sequences of the three trypanosomatids form a separate cluster. The GAT3 sequences are highly divergent from the other family members and seem to have evolved more rapidly than the others, suggesting the acquisition of a new function.Importance of the TMS2 sequence for the targeting efficiency
Figure 1s. Dendrogram showing the relationships between amino-acid sequences of described and putative peroxisomal ABC transporters. The tree was created using the ClustalX program. Positions with gaps were excluded and the tree was corrected for multiple substitutions. The closest related ABC transporter sequence of Escherichia coli was used to root the tree. Drosophila melanogaster, Caenorhabditis elegans, Yarrowia lipolytica, Neurospora crassa and Candida albicans sequences were the highest scoring hits obtained when the three GAT sequences were used in a BLAST search against the nr database at the NCBI website (http://www.ncbi.nlm.nih.gov). Numbers at the nodes represent the support for each node as obtained by 1000 bootstrap samplings. The horizontal bar represents 20 accepted amino-acid changes/100 residues.
Importance of the TMS2 sequence for the targeting efficiency
Since GFP fluorescence was not completely located in glycosomes in trypanosomes transfected with construct GFP-GAT2(1–90) (see of the article), the role of the TMS2 of GAT2 in the targeting was studied in more detail. To that end, a GAT2 construct was designed with an internal deletion removing the region encompassing amino acids 91 to 203 [GAT2(Δ91–203)-GFP] (). This brought the putative glycosomal localization domain (amino acids 68-90) in a similar context but in front of a TMS with a different amino-acid sequence (predicted TMS4). As a control, we made the alignment-based corresponding deletion construct of GAT1, GAT1(Δ97–209)-GFP. The internal deletion in GAT2 did not affect the targeting efficiency, strongly suggesting that the presence of a TMS behind the targeting motif is most important for the correct insertion of the protein into the glycosomal membrane rather than for the targeting itself. As expected, only a very weak and cytoplasmic signal could be observed in the case of GAT1, confirming the previous observation of a targeting signal present in a different alignment region for the two polypeptides.
Figure 9. Importance of TMS2 for glycosome-targeting efficiency. GAT1 and GAT2 constructs with similar deletions as inferred from sequence alignment were expressed in procyclic trypanosomes. The cells were processed for immunofluorescence with anti-hexokinase polyclonal antibody. The punctuate fluorescence pattern typical of glycosomal localization was only observed for the GAT2(Δ91–203) deletion construct. Bars, 5 µm.
Mapping of the protein-protein interaction domains of TbPEX19
T25-TbPEX19 deletion mutants were tested for interaction with T18, T18-HsPEX3(1–373) and T18-TbGAT2(1–126) in the bacterial two-hybrid system. The values are given in s.
Table Is. Mapping of the TbGAT2 and HsPEX3 binding domain of TbPEX19
We are grateful to Prof. Christine Clayton (ZMBH, Universität Heidelberg) for providing the procyclic T. brucei cell line 449 and plasmid pHD1336; to Dr Daniel Ladant (Institut Pasteur and Hybrigenics, Paris) for the gift of the bacterial two-hybrid vectors pKT25 and pUT18C; to Prof. Ron Wanders (University of Amsterdam) for making available the human PEX19-deficient fibroblasts; to Dr Myriam Baes (Katholieke Universiteit Leuven, Belgium) for the immortalized Pex5+/--fibroblasts; and to Prof. Fred Opperdoes for critical comments on this manuscript. This research was supported through grants from the ‘Fonds de la Recherche Scientifique Médicale’ (FRSM) of the ‘Communauté française de Belgique’ and the Interuniversity Attraction Poles – Belgian Federal Office for Scientific, Technical and Cultural Affairs. CY acknowledges a PhD scholarship from the ‘Fonds pour la formation à la Recherche dans l'Industrie et dans l'Agriculture’ (FRIA) and the Université catholique de Louvain. MF and CB were supported through grants from the Flemish Government (‘Geconcerteerde Onderzoeksacties GOA/2004/08)’ and the ‘Fonds voor Wetenschappelijk Onderzoek-Vlaanderen (Onderzoeksproject G.0237.04)’.
References
- Opperdoes FR, Borst P. Localization of nine glycolytic enzymes in a microbody-like organelle in Trypanosoma brucei: the glycosome. FEBS Lett 1977; 80: 360–364
- Hannaert, V, Bringaud, F, Opperdoes, FR, Michels, PAM. Evolution of energy metabolism and its compartmentation in Kinetoplastida. Kinetoplastid Biol Dis 2003;2:11. http://www.kinetoplastids.com/content/2/1/11
- Moyersoen J, Choe J, Fan E, Hol WG, Michels PAM. Biogenesis of peroxisomes and glycosomes: trypanosomatid glycosome assembly is a promising new drug target. FEMS Microbiol Rev 2004; 28: 603–643
- Eckert JH, Erdmann R. Peroxisome biogenesis. Rev Physiol Biochem Pharmacol 2003; 147: 75–121
- Subramani S, Koller A, Snyder WB. Import of peroxisomal matrix and membrane proteins. Annu Rev Biochem 2000; 69: 399–418
- Schliebs W, Kunau WH. Peroxisome membrane biogenesis: the stage is set. Curr Biol 2004; 14: R397–R399
- Jones JM, Morrell JC, Gould SJ. PEX19 is a predominantly cytosolic chaperone and import receptor for class 1 peroxisomal membrane proteins. J Cell Biol 2004; 164: 57–67
- Fang Y, Morrell JC, Jones JM, Gould SJ. PEX3 functions as a PEX19 docking factor in the import of class I peroxisomal membrane proteins. J Cell Biol 2004; 164: 863–875
- Shibata H, Kashiwayama Y, Imanaka T, Kato H. Domain architecture and activity of human Pex19p, a chaperone-like protein for intracellular trafficking of peroxisomal membrane proteins. J Biol Chem 2004; 279: 38486–38494
- Baerends RJ, Faber KN, Kiel JA, van der Klei IJ, Harder W, Veenhuis M. Sorting and function of peroxisomal membrane proteins. FEMS Microbiol Rev 2000; 24: 291–301
- Dyer JM, McNew JA, Goodman JM. The sorting sequence of the peroxisomal integral membrane protein PMP47 is contained within a short hydrophilic loop. J Cell Biol 1996; 133: 269–280
- Biermanns M, von Laar J, Brosius U, Gartner J. The peroxisomal membrane targeting elements of human peroxin 2 (PEX2). Eur J Cell Biol 2003; 82: 155–162
- Pause B, Saffrich R, Hunziker A, Ansorge W, Just WW. Targeting of the 22 kDa integral peroxisomal membrane protein. FEBS Lett 2000; 471: 23–28
- Wang X, Unruh MJ, Goodman JM. Discrete targeting signals direct Pmp47 to oleate-induced peroxisomes in Saccharomyces cerevisiae. J Biol Chem 2001; 276: 10897–10905
- Honsho M, Fujiki Y. Topogenesis of peroxisomal membrane protein requires a short, positively charged intervening-loop sequence and flanking hydrophobic segments. Study using human membrane protein PMP34. J Biol Chem 2001; 276: 9375–9382
- Jones JM, Morrell JC, Gould SJ. Multiple distinct targeting signals in integral peroxisomal membrane proteins. J Cell Biol 2001; 153: 1141–1150
- Sacksteder KA, Jones JM, South ST, Li X, Liu Y, Gould SJ. PEX19 binds multiple peroxisomal membrane proteins, is predominantly cytoplasmic, and is required for peroxisome membrane synthesis. J Cell Biol 2000; 148: 931–944
- Matsuzono Y, Kinoshita N, Tamura S, Shimozawa N, Hamasaki M, Ghaedi K, Wanders RJ, Suzuki Y, Kondo N, Fujiki Y. Human PEX19: cDNA cloning by functional complementation, mutation analysis in a patient with Zellweger syndrome, and potential role in peroxisomal membrane assembly. Proc Natl Acad Sci USA 1999; 96: 2116–2121
- Snyder WB, Koller A, Choy AJ, Subramani S. The peroxin Pex19p interacts with multiple, integral membrane proteins at the peroxisomal membrane. J Cell Biol 2000; 149: 1171–1178
- Gloeckner CJ, Mayerhofer PU, Landgraf P, Muntau AC, Holzinger A, Gerber JK, Kammerer S, Adamski J, Roscher AA. Human adrenoleukodystrophy protein and related peroxisomal ABC transporters interact with the peroxisomal assembly protein PEX19p. Biochem Biophys Res Commun 2000; 271: 144–150
- Fransen M, Brees C, Ghys K, Amery L, Mannaerts GP, Ladant D, Van Veldhoven PP. Analysis of mammalian peroxin interactions using a non-transcription-based bacterial two-hybrid assay. Mol Cell Proteomics 2002; 1: 243–252
- Fransen M, Wylin T, Brees C, Mannaerts GP, Van Veldhoven PP. Human pex19p binds peroxisomal integral membrane proteins at regions distinct from their sorting sequences. Mol Cell Biol 2001; 21: 4413–4424
- Fransen M, Vastiau I, Brees C, Brys V, Mannaerts GP, Van Veldhoven PP. Potential role for Pex19p in assembly of PTS-receptor docking complexes. J Biol Chem 2004; 279: 12615–12624
- South ST, Gould SJ. Peroxisome synthesis in the absence of preexisting peroxisomes. J Cell Biol 1999; 144: 255–266
- Honsho M, Hiroshige T, Fujiki Y. The membrane biogenesis peroxin Pex16p. Topogenesis and functional roles in peroxisomal membrane assembly. J Biol Chem 2002; 277: 44513–44524
- Banerjee SK, Kessler PS, Saveria T, Parsons M. Identification of trypanosomatid PEX19: functional characterization reveals impact on cell growth and glycosome size and number. Mol Biochem Parasitol 2005; 142: 47–55
- Wanders RJ, Tager JM. Lipid metabolism in peroxisomes in relation to human disease. Mol Aspects Med 1998; 19: 69–154
- Palmieri L, Rottensteiner H, Girzalsky W, Scarcia P, Palmieri F, Erdmann R. Identification and functional reconstitution of the yeast peroxisomal adenine nucleotide transporter. EMBO J 2001; 20: 5049–5059
- Hettema EH, van Roermund CW, Distel B, van den Berg M, Vilela C, Rodrigues-Pousada C, Wanders RJ, Tabak HF. The ABC transporter proteins Pat1 and Pat2 are required for import of long-chain fatty acids into peroxisomes of Saccharomyces cerevisiae. EMBO J 1996; 15: 3813–3822
- Mosser J, Douar AM, Sarde CO, Kioschis P, Feil R, Moser H, Poustka AM, Mandel JL, Aubourg P. Putative X-linked adrenoleukodystrophy gene shares unexpected homology with ABC transporters. Nature 1993; 361: 726–730
- Lombard-Platet G, Savary S, Sarde CO, Mandel JL, Chimini G. A close relative of the adrenoleukodystrophy (ALD) gene codes for a peroxisomal protein with a specific expression pattern. Proc Natl Acad Sci USA 1996; 93: 1265–1269
- Kamijo K, Taketani S, Yokota S, Osumi T, Hashimoto T. The 70-kDa peroxisomal membrane protein is a member of the Mdr (P-glycoprotein)-related ATP-binding protein superfamily. J Biol Chem 1990; 265: 4534–4540
- Shani N, Jimenez-Sanchez G, Steel G, Dean M, Valle D. Identification of a fourth half ABC transporter in the human peroxisomal membrane. Hum Mol Genet 1997; 6: 1925–1931
- Clayton C, Adams M, Almeida R, Baltz T, Barrett M, Bastien P, Belli S, Beverley S, Biteau N, Blackwell J, et al. Genetic nomenclature for Trypanosoma and Leishmania. Mol Biochem Parasitol 1998; 97: 221–224
- Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 1997; 25: 4876–4882
- Brun, R, Schonenberger. Cultivation and in vitro cloning or procyclic culture forms of Trypanosoma brucei in a semi-defined medium. Short communication. Acta Trop 1979;36:289–292.
- Biebinger S, Rettenmaier S, Flaspohler J, Hartmann C, Pena-Diaz J, Wirtz LE, Hotz HR, Barry JD, Clayton C. The PARP promoter of Trypanosoma brucei is developmentally regulated in a chromosomal context. Nucleic Acids Res 1996; 24: 1202–1211
- Fransen M, Vastiau I, Brees C, Brys V, Mannaerts GP, Van Veldhoven PP. Analysis of human Pex19p's domain structure by pentapeptide scanning mutagenesis. J Mol Biol 2005; 346: 1275–1286
- Amery L, Sano H, Mannaerts GP, Snider J, Van Looy J, Fransen M, Van Veldhoven PP. Identification of PEX5p-related novel peroxisome-targeting signal 1 (PTS1)-binding proteins in mammals. Biochem J 2001; 357: 635–646
- Baes M, Gressens P, Baumgart E, Carmeliet P, Casteels M, Fransen M, Evrard P, Fahimi D, Declercq PE, Collen D, et al. A mouse model for Zellweger syndrome. Nat Genet 1997; 17: 49–57
- Landgraf P, Mayerhofer PU, Polanetz R, Roscher AA, Holzinger A. Targeting of the human adrenoleukodystrophy protein to the peroxisomal membrane by an internal region containing a highly conserved motif. Eur J Cell Biol 2003; 82: 401–410
- Auerbach D, Thaminy S, Hottiger MO, Stagljar I. The post-genomic era of interactive proteomics: facts and perspectives. Proteomics 2002; 2: 611–623
- Muntau AC, Mayerhofer PU, Paton BC, Kammerer S, Roscher AA. Defective peroxisome membrane synthesis due to mutations in human PEX3 causes Zellweger syndrome, complementation group G. Am J Hum Genet 2000; 67: 967–975
- Ghaedi K, Tamura S, Okumoto K, Matsuzono Y, Fujiki Y. The peroxin pex3p initiates membrane assembly in peroxisome biogenesis. Mol Biol Cell 2000; 11: 2085–2102
- Soukupova M, Sprenger C, Gorgas K, Kunau WH, Dodt G. Identification and characterization of the human peroxin PEX3. Eur J Cell Biol 1999; 78: 357–374
- Rottensteiner H, Kramer A, Lorenzen S, Stein K, Landgraf C, Volkmer-Engert R, Erdmann R. Peroxisomal membrane proteins contain common Pex19p-binding sites that are an integral part of their targeting signals. Mol Biol Cell 2004; 15: 3406–3417
- Bakker BM, Mensonides FI, Teusink B, van Hoek P, Michels PAM, Westerhoff HV. Compartmentation protects trypanosomes from the dangerous design of glycolysis. Proc Natl Acad Sci USA 2000; 97: 2087–2092
- Michels PAM, Moyersoen J, Krazy H, Galland N, Herman M, Hannaert V. Peroxisomes, glyoxysomes and glycosomes. Mol Membr Biol 2005; 22: 133–145
- Blattner J, Helfert S, Michels P, Clayton C. Compartmentation of phosphoglycerate kinase in Trypanosoma brucei plays a critical role in parasite energy metabolism. Proc Natl Acad Sci USA 1998; 95: 11596–11600
- Furuya T, Kessler P, Jardim A, Schnaufer A, Crudder C, Parsons M. Glucose is toxic to glycosome-deficient trypanosomes. Proc Natl Acad Sci USA 2002; 99: 14177–14182
- Moyersoen J, Choe J, Kumar A, Voncken FG, Hol WGJ, Michels PAM. Characterization of Trypanosoma brucei PEX14 and its role in the import of glycosomal matrix proteins. Eur J Biochem 2003; 270: 2059–2067
- Visser WF, van Roermund CW, Waterham HR, Wanders RJ. Identification of human PMP34 as a peroxisomal ATP transporter. Biochem Biophys Res Commun 2002; 299: 494–497
- Michels PAM, Hannaert V, Bringaud F. Metabolic aspects of glycosomes in Trypanosomatidae-new data and views. Parasitol Today 2000; 16: 482–489
- Halbach A, Lorenzen S, Landgraf C, Volkmer-Engert R, Erdmann R, Rottensteiner H. Function of the PEX19-binding site of human ALDP as targeting motif in man and yeast: PMP targeting is evolutionarily conserved. J Biol Chem 2005; 280: 21176–28182
- Boisnard S, Zickler D, Picard M, Berteaux-Lecellier V. Overexpression of a human and a fungal ABC transporter similarly suppresses the differentiation defects of a fungal peroxisomal mutant but introduces pleiotropic cellular effects. Mol Microbiol 2003; 49: 1287–1296