Abstract
The cationic amphipathic α-helical antibiotic peptide, pleurocidin, from the winter flounder Pleuronectes americanus associates strongly with anionic membranes where it is able to translocate across the membrane, cause dye leakage from vesicles and induce pore like channel conductance. To investigate the mechanism of pleurocidin antibiotic activity in more detail we have applied a variety of spectroscopic techniques to study the interaction of pleurocidin with model membranes. At neutral pH the peptide inserts into membranes containing anionic lipids and, as shown by proton-decoupled 15N solid-state NMR spectroscopy of macroscopically oriented samples, is aligned parallel to the membrane surface. 2H solid-state NMR spectroscopy of chain deuterated phosphatidylethanolamine (PE) and phosphatidylglycerol (PG) lipids in mixed membranes shows that pleurocidin interacts with both the zwitterionic PE and anionic PG but disrupts the lipid acyl chain order of the anionic PG lipids more effectively. At acidic pH the three histidine residues of pleurocidin become protonated and positively charged which does not alter the membrane disrupting effect nor the location of the peptide in the membrane. The results are interpreted in terms of a structural model for pleurocidin inserted into anionic lipid membranes and the implications of our data are discussed in terms of a general mechanism for the antibiotic activity.
Introduction
Pleurocidin is a cationic amphipathic helical Citation[1] peptide antibiotic found in the skin Citation[2] and intestine Citation[3] of the winter flounder Pleuronectes americanus. Pleurocidin shares sequence homology with other antibiotic peptides from diverse sources such as dermaseptin Citation[4] and maculatin Citation[5] from different species of tree frog and ceratotoxins from the female Mediterranean fruit fly Citation[6] (). Pleurocidin is similarly capable of causing dye leakage from liposomes Citation[7], translocating across model membranes Citation[7] and demonstrates pore forming activity in planar lipid bilayers Citation[8]. All of these properties are greatly enhanced when the membranes are partly composed of anionic lipids. Some potentially interesting comparisons can also be made with antibiotic peptides discovered from other fish (). Pleurocidin is approximately the same length as moronecidin or piscidins from hybrid striped bass Citation[9–11] and chrysophsin from red sea bream Citation[12]. A helical wheel diagram of these peptides predicts an amphipathic helix with charged residues on one face with hydrophobic residues clustered on the other, but of particular note, in contrast with the other cationic peptides mentioned, is the prevalence of histidine residues in the sequence of all three peptides which would lead to a dramatic increase in charge at acidic pH.
Table I. Sequences of selected antibiotic helical peptides. Residues that are positively charged at pH 7 or pH 5 are highlighted in bold whilst negatively charged residues are marked in grey. *Moronecidin is also known as piscidin 2.
Comparative studies of cationic antimicrobial peptides from a variety of structural classes have given further insights into the mechanism of action Citation[13]. Though cationic antimicrobial peptides are structurally diverse, the secondary structure is usually composed of both a hydrophobic and hydrophilic surface which has been taken as evidence that some form of permeabilization of the bacterial membrane is the most likely mechanism of action Citation[13]. However, it has been shown that, for a number of cationic peptides, membrane depolarization is incomplete even when 90% or more bacteria have been killed and hence, although the membrane interactions of cationic antibiotic peptides are an important factor, it may be important for entry into the cell but not constitute the main killing mechanism among many classes of peptide Citation[14], Citation[15].
Nonetheless, cationic amphipathic α-helical peptides, including pleurocidin, have been shown to be capable of depolarizing membranes and a strong interaction between the positively charged residues and the anionic lipid head groups is thought to be involved in this process Citation[8]. The number and position of histidine residues in derivatives of a model peptide, LAH4, have been shown, using solid-state NMR methods, to be important in determining the membrane disruption effect at acidic pH and the ability of the peptides to cross membranes with DNA cargo Citation[16]. Since the localization of pleurocidin expression reveals the peptide to be distributed in both mucin skin granules and intestinal goblet cells Citation[3], it may function over a range of pH in vivo. Hence, by varying the pH whilst conducting a range of biophysical experiments, the role of histidine residues in pleurocidin activity can be determined and the potential importance of membrane disruption in the mechanism of pleurocidin antibiotic activity assessed. We have used intrinsic fluorescence and solid-state NMR methods to study the interaction of pleurocidin with model bilayers mimicking bacterial membranes to determine the mechanism of action of pleurocidin as an antibiotic. Our results show that pleurocidin interacts strongly with, and inserts into, anionic phosphatidylglycerol containing membranes. It is oriented parallel to the membrane surface and capable of disrupting lipid chain order in anionic deuterated lipids to a greater degree to that of similarly deuterated zwitterionic lipids. Protonation of the histidine residues at acidic pH however, does not increase the disruption of membrane order by pleurocidin. These results are interpreted in terms of a structural model which has important implications for the mechanism of antibiotic action of pleurocidin and other cationic amphipathic peptides which are discussed in the context of existing models.
Material and methods
Peptide and lipids
Pleurocidin amide () containing three 15N labels (Ala10, Ala19 and Leu21) was synthesized using standard FMOC solid-state chemistry on a Millipore 9050 synthesizer. In crude peptide preparations a predominant peak was observed when analysed by HPLC with acetonitrile/water gradients. During HPLC purification the main peak was collected and the identity of the product confirmed by matrix assisted laser desorption ionization (MALDI) mass spectrometry. The lipids 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphatidylcholine (POPC), 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphatidylglycerol (POPG), 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphatidylethanolamine (POPE), 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphatidyserine (POPS), 1-palmitoyld31-2-oleoyl-sn-glycero-3-phosphatidylcholine (POPC-d31) and 1-palmitoyld31-2-oleoyl-sn-glycero-3-phosphatidylglycerol (POPG-d31) were obtained from Avanti Polar Lipids, Inc. (Alabaster, AL) and used without further purification. All other reagents were analytical grade or better.
Sample preparation for solid-state NMR
For solid-state NMR, samples with different lipid compositions were prepared for 2H-echo NMR; POPE/POPE-d31/POPG and POPE/POPG-d31. The molar ratios of POPE/POPE-d31/POPG and POPE/POPG-d31 were 50:25:25 and 75:25 respectively. A total of around 5 mg lipids per sample were dissolved and mixed in chloroform and dried under rotor-evaporation at room temperature. In order to remove all organic solvent, the lipid films were exposed to vacuum overnight. The films were then rehydrated with 5 ml of a suspension of pleurocidin in 0.1 M Tris, 0.1 M KCl buffer at pH 7.5 at room temperature. For wideline 2H NMR experiments pleurocidin was added at 2.5% by mol. The samples were briefly sonicated in a bath sonicator to improve exposure of all lipids to the peptide. Samples were subjected to five rapid freeze-thaw cycles for further sample homogenization and then centrifuged at 21000 g for 20 min at room temperature. The pellets, containing lipid vesicles and associated pleurocidin were transferred to 4 mm MAS rotors for NMR measurements (Bruker, Karlsruhe, Gemany). Lipid vesicles were also prepared in this way in the absence of pleurocidin. After solid-state NMR measurements were recorded on these samples at pH 7.5 the pellets were resuspended in a large excess of buffer (0.1 M Tris-HCl, 0.1 M KCl) at pH 5. The samples were then sonicated in a bath sonicator for 5 min and again centrifuged as above and loaded into MAS rotors for further NMR measurements.
Static aligned samples were prepared using a procedure whereby a sublimable solid is co-dissolved with the lipids and peptide to aid alignment during dehydration and rehydration Citation[17]. Then 4 mg of pleurocidin, carrying 15N labels at position Ala10, Ala19 and Leu21 was co-dissolved with POPE/POPG (75:25) in chloroform at a molar ratio of 2.5 mole%. The solvent was evaporated under a stream of nitrogen gas and re-dissolved in a solution of chloroform containing naphthalene. The molar ratio of naphthalene to lipid was 1:1. The solution was layered over 15 thin glass plates (9×22 mm, Marienfeld GmbH, Lauda-Königshofen, Germany) and the plates were then exposed to high vacuum overnight. The samples were rehydrated at 93% humidity and at 37°C.
Solid-state NMR
2H quadrupole experiments Citation[18] for samples containing either POPE-d31 or POPG-d31 were performed at 46.10 MHz on a Bruker Avance 300 NMR spectrometer using a 4 mm MAS probe, spectral width of 200 KHz and with recycle delay, echo delay, acquisition time and 90° pulse lengths of 0.3 s, 42 µs, 2 ms and 5 µs respectively. The temperature was maintained at 303 K to keep the bilayers in their liquid-crystalline phase. During processing the first 40 points were removed in order to start Fourier-transformation at the echo maximum. Spectra were zero filled to 8 k points and 50 Hz exponential line-broadening was applied. Smoothed deuterium order parameter profiles were obtained from symmetrized and dePaked 2H-NMR powder spectra of POPE-d31 and POPG-d31 using published procedures Citation[19–21].
15N cross polarization (CP) spectra of static aligned samples were acquired at 40.54 MHz for 15N on a Bruker Avance 400 NMR spectrometer equipped with a double resonance flat-coil probe. A simple adiabatic CP pulse sequence was used with a spectral width, acquisition time, CP contact time and recycle delay time of 25 kHz, 10 ms, 800 µs and 3 s respectively. The 1H π/2 pulse and spinal64 heteronuclear decoupling field strengths were 42 kHz. 40 k scans were accumulated and the spectra were zero filled to 4 k points and 100 Hz exponential line-broadening was applied during processing. Spectra were externally referenced to 15NH4Cl at 41.5 ppm.
31P spectra with proton-decoupling were acquired at 161.953 MHz for 31P on a Bruker Avance 400 NMR spectrometer equipped with a double resonance flat-coil probe. Proton decoupled 31P spectra were acquired using one 4 µs 90° pulse, followed by continuous wave heteronuclear 1H decoupling at a field strength of 25 kHz. The spectral sweep width was 75 kHz and the recycle delay and acquisition times were 5 s and 6.8 ms respectively. The temperature was maintained at 303 K. Spectra were zero filled to 16 k points and 50 Hz exponential line broadening was applied during processing. Spectra were referenced externally to 85% H3PO4 at 0 ppm.
Fluorescence spectroscopy
Emission spectra of the intrinsic fluorescence of Trp2 or Tyr24 were acquired using a Jasco FP-6300 spectrometer at room temperature. Vesicle suspensions were prepared as for solid-state NMR experiments above except the freeze thaw cycles were omitted such that multi lamellar vesicles were prepared. Vesicles containing POPC, POPC/POPG (75:25) and POPC/POPS (75:25) were prepared in addition to the POPE/POPG vesicles described above. 150 µl of these suspensions containing pleurocidin at a concentration of 0.2 mg/ml, was added to 0.85 ml of 0.1 M Tris, 0.1 M KCl buffer at pH 7.5 to produce a final peptide concentration of about 0.03 mg/ml. Tryptophan emission spectra of the lipid/peptide suspension were acquired by scanning from 310–450 nm using an excitation wavelength of 295 nm with a spectral bandwidth of 5 nm for both excitation and emission. A spectrum of the aqueous peptide was acquired at a peptide concentration of 0.2 mg/ml in the same buffer. For fluorescence quenching experiments, 40% acrylamide solution was added stepwise to a final concentration of 0.24 M at each step, equilibration of the sample was ensured and all spectra are an average of three scans.
Results
Fluorescence spectroscopy
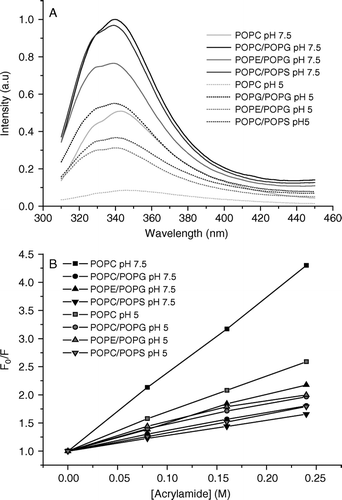
The interaction of pleurocidin with zwitterionic DOPC or mixed DOPC/DOPG membranes has been previously monitored by circular dichrosim and the intrinsic fluorescence of its Trp2 residue Citation[7]. This analysis revealed that the presence of anionic lipids is crucial for the induction of α-helical content and concomitant membrane association. We have used a similar fluorescence technique and have extended it to study the effect of varying the composition of the lipid and pH on pleurocidin binding. The intrinsic fluorescence spectra of Trp2 in the presence of lipid vesicles are shown (A) and vary dramatically depending on the composition of the lipid vesicles. A spectrum of aqueous pleurocidin revealed a maximum at 360 nm (not shown), whilst in the presence of lipid vesicles the spectra are dramatically blue shifted and have maxima between 339 and 343 nm. This indicates that the tryptophan residue experiences a hydrophobic environment and confirms that the peptide is interacting with the lipid vesicles. In the presence of anionic lipids such as PG or PS the spectra become of greater intensity when compared with that obtained in the presence of zwitterionic PC vesicles. The maximum also undergoes a blue shift to 340 nm for POPC/POPG and 339 nm for both POPC/POPS and POPE/POPG compared with 343 nm for POPC vesicles. These results confirm the previous study and indicate that the tryptophan residue penetrates further into the hydrophobic core of the membrane when anionic lipids are present. The samples prepared in pH 7.5 buffer were pelleted by centrifugation and resuspended in pH 5 buffer. The spectra of the samples at pH 5 are characterized by a slight red shift for the POPC sample and reduction of intensity for all samples. The maxima for POPC/POPG, POPE/POPG and POPC/POPS vesicles at acidic pH are 55, 41 and 39% of the maxima at neutral pH respectively. The maximum for the POPC sample is only 17% of its neutral value again suggesting that the insertion of pleurocidin into zwitterionic membranes is much weaker than when anionic lipids are present. This may indicate movement of the tryptophan residue into a more polar environment but is more likely to be a reflection of both the loss of peptide during the preparation procedure and the protonation of the three histidine residues at this pH which become more effective quenchers of tryptophan fluorescence Citation[22]. A more accurate comparison of the depth profile for Trp2 of pleurocidin can be achieved by comparing fluorescence quenching induced by an aqueous quencher such as acrylamide. The Stern-Vollmer plots of the quenching ability of acrylamide on intrinsic fluorescence resulting from Trp2 (B) confirm the analysis above that anionic lipids cause the pleurocidin N-terminus to penetrate further into the hydrophobic core since the acrylamide induced quenching is much greater in the presence of POPC liposomes than either in the presence of POPC/POPG, POPC/POPS or POPE/POPG liposomes at pH 7.5. Acidification causes very little difference in quenching profiles in the preparations containing anionic lipids indicating no significant change in the depth profile of the peptide N-terminus. A more marked change in quenching profile can be observed for the zwitterionic POPC vesicles at acidic pH but this is likely to reflect the removal of loosely or unbound pleurocidin, which could constitute as much as 69% of the total peptide, during preparation of the sample.
Figure 1. Intrinsic tryptophan fluorescence spectra of Trp2 of pleurocidin in the presence of liposomes of varying lipid composition at neutral and acidic pH are shown (A). The presence of PG causes a slight blue shift and increase in fluorescence intensity irrespective of the zwitterionic component of the liposomes whilst the spectra obtained at acidic pH are of reduced intensity. A Stern-Vollmer plot of the effect of adding an aqueous acrylamide quencher on tryptophan fluorescence intensity (B) reveals the depth of pleurocidin penetration into the model membranes.
Deuterium NMR
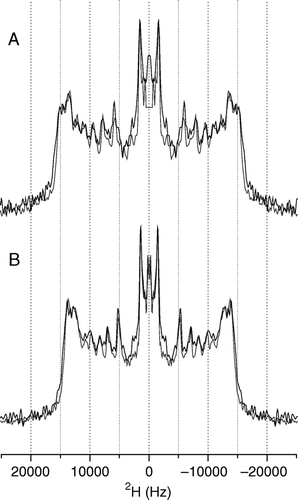
A detailed picture of the effect of pleurocidin on the lipid chains in model membranes is obtained by acquiring wideline 2H echo spectra of the lipid mixes in the absence and presence of pleurocidin at the two pHs. One method of demonstrating the effect of pleurocidin is to add peptide at increasing concentrations and to observe the effect on the lipid spectra. In this case increasing the peptide concentration much above 3 mole% is likely to induce detergent like behaviour from the peptides as has been shown for the cationic amphipathic α-helical peptides magainin Citation[23] and LAH4 Citation[16] which have been shown to induce spectral changes in 31P NMR spectra of PC bilayers consistent with the formation of small disk-like bicelles or perforated sheets. By keeping the peptide concentration constant, at a level (2.5 mole%) where an effect on fatty acyl chain order can be observed by solid-state NMR methods but without disintegration of the membrane, and varying the lipid conditions, we can obtain an accurate measure of the role that the differing lipids play in the peptide-lipid interactions. 2H echo spectra are acquired for hydrated powder samples pelleted by centrifugation from excess buffered solutions which allow relatively rapid preparation but, more importantly, this process facilitates control of the sample pH. Solid-state 2H echo spectra of either POPE-d31 or POPG-d31 in mixed POPE/POPE-d31/POPG (A) or POPE/POPG-d31 (B) vesicles reveal the strength of the interaction between the peptide and either the zwitterionic or anionic lipid component and the ability of the peptide to disrupt the lipid acyl chains Citation[16]. The disruption of the lipid chain order induced by 2.5 mole% pleurocidin can be seen in the spectra and by either 2.5 or 3.3 mole% pleurocidin in the calculated order parameter profiles which are displayed relative to the peptide free data (C). It can be seen that, particularly further down the acyl chain, the anionic PG acyl chains are strongly disrupted by the presence of pleurocidin (B, C) whilst the acyl chains of the zwitterionic PE lipids are less affected (A, C). No enhancement of the lipid acyl chain disordering effect was seen in any of the liposome systems at acidic pH as is demonstrated clearly in the spectra recorded of POPE/POPE-d31/POPG (2:1:1) POPE/POPG-d31 (3:1) at both pH 7.5 and pH 5 in the presence of 2.5 mole% pleurocidin (A, B).
Figure 2. 2H echo spectra of POPE-d31 or POPG-d31 in lipid vesicles. Vesicles contained POPE/POPE-d31/POPG (2:1:1) (A) or POPE/POPG-d31 (3:1) (B) in the absence (grey lines) or presence (black lines) of 2.5 mole% pleurocidin at pH 7.5. Spectra were recorded on a Bruker Avance 300 spectrometer at 298K. 2H order parameter profiles for the same peptide containing vesicles at pH 7.5 and also with an elevated pleurocidin concentration of 3.3 mole% are shown calculated relative to peptide free vesicles (C).
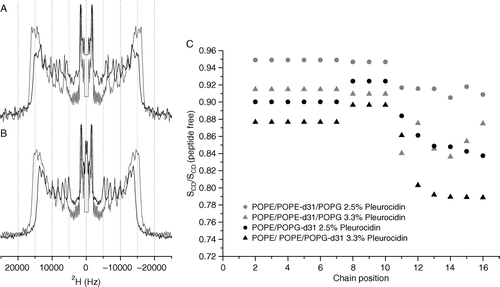
Figure 3. 2H echo spectra of POPE-d31 or POPG-d31 in lipid vesicles containing POPE/POPE-d31/POPG (2:1:1) (A) or POPE/POPG-d31 (3:1) (B) and 2.5 mole% pleurocidin at pH 7.5 (grey line) and pH 5 (black line). The change in pH has very little effect on the spectra. Spectra were recorded on a Bruker Avance 300 spectrometer at 298K.
NMR of aligned samples
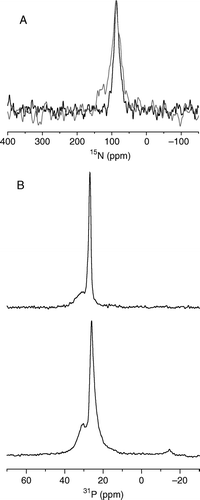
The preparation of samples containing pleurocidin simultaneously labelled with 15N at positions Ala10, Ala19 and Leu21 and aligned on glass plates and the subsequent measurement of the static proton-decoupled 15N CP spectrum allows an accurate determination of the value of the anisotropic chemical shift (A). A value of 87 ppm indicates that the long axis of the peptide helix is oriented parallel to the membrane surface. The peak is a result of the contributions from three residues but analysis by a peak fitting routine did not show any indication of shoulders or multiple components, indicating that the three labelled sites in the peptide adopt a similar orientation, although the resonance full width at half height (FWHH) of almost 17 ppm constitutes a spread of orientations and is a reflection of the disruption of the POPE/POPG membrane induced by the peptide. The quality of orientation of the POPE/POPG membranes and the effect of incorporating the peptide can be seen by obtaining static 31P spectra (B). The spectra obtained in the absence (B top) and presence (A bottom) of pleurocidin reveal that the samples remain oriented with respect to the external magnetic field but the peptide does induce some linebroadening consistent with the disruption of the lipid bilayers observed above. A spectrum was also acquired for a sample prepared similarly but with the pH controlled at pH 5. The line becomes rather broader (FWHH 28.5 ppm) but a value of 88 ppm indicates that no significant change in orientation accompanies the change in pH.
Figure 4. The static 15N NMR spectrum of 15N-alanine10, alanine19 and leucine21 labelled pleurocidin reconstituted in POPE/POPG (3:1) bilayers and deposited on glass plates oriented perpendicular to B0 (A). Spectra were acquired for samples prepared at both pH 7 (black line) and pH 5 (grey line). The observed chemical shift is characteristic of a peptide oriented parallel to the membrane surface. 31P echo spectra of POPE/POPG (3:1) bilayers (B), prepared in the same way at pH 7, in the absence (top) and presence (bottom) of 2.5 mole% pleurocidin.
Discussion
The bacterial membrane has often been considered the main site of action for antimicrobial peptides and whilst there is no doubt that such peptides interact with membranes, often rather high levels of peptide are required to induce effects such as lipid flip-flop or the release of calcein from model liposome systems. The implications of such observations, which are supported by a recent study finding no direct relationship between any particular measurement of membrane activity, including membrane permeabilisation, and peptide antibacterial activity Citation[24], are that the action of antimicrobial peptides against their bacterial targets may involve a variety of factors in combination. Indeed, the interactions of cationic antimicrobial peptides with model membranes can be extremely heterogeneous Citation[24] and traditional structural techniques will have to evolve and be used in combination if a complete understanding of such complicated processes is to be achieved. In light of this, we have conducted a study of the α-helical antimicrobial peptide pleurocidin from the winter flounder Pleuronectes americanus. This peptide is particularly interesting since it shares sequence homology with some comparatively well studied antibiotics from diverse sources in nature Citation[4–6], Citation[25–28] but, in common with a handful of other such peptides found in fish, is rich in histidine residues (). We have been interested in the role that histidine residues play in the delivery of nucleic acids across membranes Citation[16], Citation[29] and the antibiotic activity [Mason AJ, Gasnier C, Kichler A, Prévost G, Metz-Boutigue M-H, Bechinger B, unpublished work, 30] of the designed peptide LAH4 Citation[31]. Here we have applied a range of biophysical techniques to obtain a structural view of pleurocidin in model membrane environments that are designed to mimic the bacterial membrane and as a result have obtained some insight into how pleurocidin causes its various membrane disruptive effects and also the potential role of histidine residues in the antibiotic activity of this naturally occurring peptide. We consider our model membranes as a suitable system to study a number of peptide-membrane interactions that are likely to be important determinants of antibiotic activity, however our symmetric system does not include other biologically relevant effects that may arise in an asymmetric model system such as electrochemical gradients and the effects of external versus internal membrane leaflet curvature and hence lateral pressure which might also affect peptide insertion into the membrane.
Our intrinsic fluorescence measurements are in agreement with a previous study Citation[7] which demonstrated that pleurocidin inserts into lipid membranes only in the presence of anionic lipids. This study further reported that high peptide translocation activity and strong dye leakage activity are only induced by pleurocidin again in the presence of anionic lipids Citation[7]. Furthermore, macroscopic conductance measurements performed on planar lipid bilayers revealed that, again, pleurocidin was only able to demonstrate pore forming activity in mixed membranes in the presence of the anionic lipid phosphatidylserine Citation[8]. In the present study, the apparent disruption of acyl chain order in the anionic lipid phosphatidylglycerol, in mixed membranes, which is of a greater magnitude than that observed for the zwitterionic phosphatidylethanolamine lipid acyl chains, suggests a mechanism for the induction of membrane leakage by pleurocidin. Our solid-state NMR study has been performed at a peptide lipid ratio of 1:40 where calcein leakage from PC/PG vesicles has been shown to be around 30%, with 100% leakage only observed at a peptide to lipid ratio of 1:10 Citation[7]. Nonetheless, the observed changes in the spectra can be interpreted as a general increase in the mobility of the lipid chain where the strongly cationic pleurocidin inserts into the membrane and binds preferentially to the anionic PG in the head group interface region without causing an overall change in the morphology of the membrane. The resulting increase in the separation of the anionic lipids allows an increase in mobility of the lipids which is reflected in the observed reduction of order parameter throughout the acyl chain.
In a previous study Citation[16] we showed that, the cationic peptide, LAH4 associates preferentially with anionic lipids in mixed membranes similar to those used in the present study. It is likely that electrostatic interactions drive this interaction for both peptides but without quantifying the strength of this interaction it is not possible to determine the contributions of an increased association of pleurocidin and PG or a more effective disruption of PG by pleurocidin, perhaps due to the relatively small head group of PE, towards enhanced disruption of PG versus PE in this model system. This aspect of the mechanism is likely to be relevant to other similar cationic peptides and constitutes an important future research topic.
The tryptophan residue (Trp2) can be seen from the fluorescence spectra and quenching data to be dipping into the hydrophobic core whilst the static solid-state 15N NMR spectrum of pleurocidin aligned in PE/PG (3:1) membranes indicates an in plane orientation as is observed with other amphipathic helical peptides such as magainin Citation[32]. This is also in agreement with hydrophobic and amphipathic model peptides which indicate stable in-plane alignment in the presence of at least three lysine residues Citation[30]. Our determination of a surface alignment for pleurocidin is also in agreement with the proposed pore forming mechanism derived from macroscopic conductance measurements Citation[8] where a distribution of the peptide along the membrane surface is expected.
Pleurocidin, like other antimicrobial peptides from fish () are rich in histidine residues which become protonated below a pH of approximately 6.1. Since a strong interaction has been demonstrated between pleurocidin and anionic lipids and as pleurocidin has been localized in intestinal epithelial cells, we investigated the effect of acidic pH on the behaviour of pleurocidin with our model membranes. The fluorescence quenching experiments reveal that a reduction in pH in PE/PG membranes, which most closely mimic bacterial membranes, causes no dramatic change in the location of the tryptophan residue (Trp2), situated close to the N-terminus of the peptide, in the membrane. In addition, no significant changes are seen in the 2H NMR spectra of any of the model membrane systems and hence the protonation of the three histidine residues and the dramatic change in the overall charge of the peptide causes no increase in the membrane disruptive effect. It is also of note that similar studies of the interaction of histidine rich peptides derived from LAH4, which contain lysine residues only at the N and C termini, with POPE/POPG membranes indicated a much greater level of POPG acyl chain disruption at the same peptide concentration [Mason AJ, Gasnier C, Kichler A, Prévost G, Metz-Boutigue M-H, Bechinger B, unpublished work]. This further supports our conclusion that the histidine residues of pleurocidin play no part in membrane disruption.
The structure of pleurocidin has been determined by solution NMR in dodecylphosphocholine (DPC) micelles, a membrane mimicking environment, to be a rigid α-helix Citation[1]. Taking this information, together with the fluorescence data and the 15N NMR data, we can build a model of pleurocidin and consider it in relation to the membrane environment (). A surface oriented α-helix with tryptophan and tyrosine residues oriented towards the hydrophobic core of the membrane and with positively charged lysine residues interacting with the phospholipid head groups at the membrane surface, leaves all three histidine residues (His11, 15 and 23) oriented towards the extra cellular milieu. Looking down the spine of the peptide these histidine residues can be seen to adopt a very similar orientation and are flanked by lysine residues which would likely prevent a strong interaction between these histidine residues and anionic lipid head groups developing even when protonated at acidic pH. If the α-helical structure was maintained for the entire length of the peptide, His23 would be oriented closer to the lipid head groups but the NMR data Citation[1] shows the structure to become disordered towards the C terminus such that His23 is oriented similarly to His11 and His15. Our data shows that this residue does not enhance the disruption of the acyl chains and hence also supports this aspect of the previous structure determination.
Figure 5. Structure of Pleurocidin based on the coordinates 1Z64.pdb obtained from the protein databank Citation[1] with the cationic and intrinsic fluorescing residues shown. A side view (A) reveals the distribution of the lysine and histidine residues along one face of the helix and the relative orientation of the tryptophan and tyrosine side chains to the helix. A view along the helix (B) shows that all three histidine residues are oriented along the spine of the helix and would interact only weakly with the lipid head groups of anionic lipids when oriented in this manner. This Figure is reproduced in colour in Molecular Membrane Biology online.
![Figure 5. Structure of Pleurocidin based on the coordinates 1Z64.pdb obtained from the protein databank Citation[1] with the cationic and intrinsic fluorescing residues shown. A side view (A) reveals the distribution of the lysine and histidine residues along one face of the helix and the relative orientation of the tryptophan and tyrosine side chains to the helix. A view along the helix (B) shows that all three histidine residues are oriented along the spine of the helix and would interact only weakly with the lipid head groups of anionic lipids when oriented in this manner. This Figure is reproduced in colour in Molecular Membrane Biology online.](/cms/asset/d186911d-dec0-4793-90c6-af1125c16e11/imbc_a_148513_f0005_b.jpg)
Pleurocidin is one of a number of cationic antimicrobial peptides that may have multiple modes of activity. Since the histidine residues in pleurocidin appear to have no role in membrane disruption at acidic pH, and taking into account both their distribution in the modelled structure and the high translocation activity of pleurocidin, we propose that the histidine residues may contribute to an intracellular killing strategy. This strategy may be related to the DNA binding property of histidine residues, which we have previously exploited in our studies of cationic amphipathic vector peptides Citation[16], Citation[29] and the known ability of pleurocidin to inhibit nucleic acid synthesis Citation[33] or potentially the histidine residues may also have a role in fine tuning the partitioning of the peptide between membrane and solution under physiological conditions. A role for DNA binding in antimicrobial activity has been proposed previously for tachyplesin Citation[34] and buforin 2 Citation[35], and the ability of cationic peptides to inhibit nucleic acid or protein synthesis has now been identified for a variety of peptides Citation[36].
Conclusions
Our study of the membrane interaction of the cationic amphipathic antimicrobial peptide pleurocidin has provided an understanding of how the peptide destabilizes anionic lipids in mixed membranes leading to disruption of the membrane integrity. In addition, a structural characterization of the peptide, taken with data from previous studies, indicates that histidine residues in the peptide are not involved in membrane destabilization even at acidic pH, and may instead be central to the intracellular antimicrobial strategies of pleurocidin and indeed other related peptides.
This paper was first published online on prEview on 27 January 2006.
A.J.M. thanks Gérard Nullans for mass spectrometry analysis and Clemens Glaubitz for access to the fluorescence facility at the J.W. Goethe University, Frankfurt, Germany. This work was supported by Vaincre la Mucoviscidose TG0501 and a CNRS ‘Poste Rouge’ awarded to A.J.M.
References
- Syvitski RT, Burton I, Mattatall NR, Douglas SE, Jakeman DL. Structural characterization of the antimicrobial peptide pleurocidin from winter flounder. Biochemistry 2005; 44: 7282–7293
- Cole AM, Weis P, Diamond G. Isolation and characterisation of pleurocidin, an antimicrobial peptide in the skin secretions of winter flounder. J Biol Chem 1997; 272: 12008–12013
- Cole AM, Darouiche RO, Legarda D, Connell N, Diamond G. Characterization of a fish antimicrobial peptide: gene expression, subcellular localization, and spectrum of activity. Antimicrob Agents Chemother 2000; 44: 2039–2045
- Pouny Y, Rapaport D, Mor A, Nicolas P, Shai Y. Interaction of antimicrobial dermaseptin and its fluorescently labeled analogues with phospholipid membranes. Biochemistry 1992; 31: 12416
- Rozek T, Waugh RJ, Steinborner ST, Bowie JH, Tyler MJ, Wallace JC. The maculatin peptides from the skin glands of the tree frog Litoria genimaculata: a comparison of the structures and antibacterial activities of maculatin 1.1 and caerin 1.1. J Peptide Sci 1998; 4: 111–115
- Marri L, Dallai R, Marchini D. The novel antibacterial peptide ceratotoxin A alters permeability of the inner and outer membrane of Escherichia coli K-12. Curr Microbiol 1996; 33: 40–43
- Yoshida K, Mukai Y, Niidome T, Takashi C, Tokunaga Y, Hatakeyama T, Aoyagi H. Interaction of pleurocidin and its analogs with phospholipids membrane and their antibacterial activity. J Peptide Res 2001; 57: 119–126
- Saint N, Cadiou H, Bessin Y, Molle G. Antibacterial peptide pleurocidin forms ion channels in planar lipid bilayers. Biochim Biophys Acta 2002; 1564: 359–364
- Lauth X, Shike H, Burns JC, Westerman ME, Ostland VE, Carlberg JM, Van Olst JC, Nizet V, Taylor SW, Shimizu C, Bulet P. Discovery and characterization of two isoforms of moronecidin, a novel antimicrobial peptide from hybrid striped bass. J Biol Chem 2002; 277: 5030–5039
- Silphaduang U, Noga EJ. Peptide antibiotics in mast cells of fish. Nature 2001; 414: 268–269
- Chinchar VG, Bryan L, Silphadaung U, Noga E, Wade D, Rollins-Smith L. Inactivation of viruses infecting ectothermic animals by amphibian and piscine antimicrobial peptides. Virology 2004; 323: 268–275
- Iijima N, Tanimoto N, Emoto Y, Morita Y, Uematsu K, Murakami T, Nakai T. Purification and characterization of three isoforms of chrysophsin, a novel antimicrobial peptide in the gills of the red sea bream Chrysophrys major. Eur J Biochem 2003; 270: 675–686
- Bechinger B. The structure, dynamics and orientation of antimicrobial peptides in membranes by multidimensional solid-state NMR spectroscopy. Biochim Biophys Acta 1999; 1462: 157–183
- Friedrich CL, Moyles D, Beveridge TJ, Hancock REW. Antibacterial action of structurally diverse cationic peptides on Gram-positive bacteria. Antimicrob Agents Chemother 2000; 44: 2086–2092
- Wu M, Amier E, Benz R, Hancock REW. Mechanism of interaction of different classes of cationic antimicrobial peptides with planar bilayers and with the cytoplasmic membrane of Escerichia coli. Biochemistry 1999; 38: 7235–7242
- Mason, AJ, Martinez, A, Glaubitz, C, Danos, O, Kichler, A, Bechinger, B. The antibiotic and DNA transfecting peptide LAH4 selectively associates with, and disorders, anionic lipids in mixed membranes. FASEB J, Dec 13 2005 10.1096/fj.05-4293fje..
- Hallock KJ, Henzler Wildman K, Lee D-K, Ramamoorthy A. An innovative procedure using a sublimable solid to align lipid bilayers for solid-state NMR studies. Biophys J 2002; 82: 2499–2503
- Davis JH. The description of membrane lipid conformation order and dynamics H-2-NMR. Biochim Biophys Acta 1983; 737: 117–171
- Schäfer H, Mädler B, Volke F. De-PAKE-ing of NMR powder spectra by nonnegative least- squares analysis with Tikhonov regularization. J Magn Reson 1995; 116: 145–149
- Sternin E, Bloom M, MacKay AL. De-PAKE-ing of NMR spectra. J Magn Reson 1983; 55: 274–282
- Seelig A, Seelig J. Dynamic structure of fatty acyl chains in a phospholipid bilayer measured by deuterium magnetic-resonance. Biochemistry 1974; 13: 4839–4845
- Chen Y, Barkley MD. Toward understanding tryptophan fluorescence in proteins. Biochemistry 1998; 37: 9976–9982
- Bechinger B. Detergent-like properties of magainin antibiotic peptides: A 31P solid-state NMR spectroscopy study. Biochim Biophys Acta 2005; 1712: 101–108
- Zhang L, Rozek A, Hancock REW. Interaction of cationic antimicrobial peptides with model membranes. J Biol Chem 2001; 276: 35714–35722
- Marcotte I, Wegener KL, Lam Y-H, Cia BCS, de Planque MRR, Bowie JH, Auger M, Separovic F. Interaction of antimicrobial peptides from Australian amphibians with lipid membranes. Chem Phys Lipids 2003; 122: 107–120
- Balla MS, Bowie JH, Separovic F. Solid-state NMR of antimicrobial peptides from Australian frogs in phospholipid membranes. Eur Biophys J 2004; 33: 109–116
- Niidome T, Kobayashi K, Arakawa H, Hateyama T, Aoyagi H. Structure-activity relationship of an antibacterial peptide, Maculatin 1.1, from the skin glands of the tree frog, Litoria genimaculata. J Peptide Sci 2004; 10: 414–422
- Ambroggio EE, Separovic F, Bowie J, Fidelio GD. Surface behaviour and peptide-lipid interactions of the antibiotic peptides, Maculatin and Citropin. Biochim Biophys Acta 2004; 1664: 31–37
- Kichler A, Leborgne C, März J, Danos O, Bechinger B. Histidine-rich amphipathic peptide antibiotics promote efficient delivery of DNA into mammalian cells. Proc Natl Acad Sci USA 2003; 96: 1564–1568
- Vogt TCB, Bechinger B. The interactions of histidine-containing amphipathic helical peptide antibiotics with lipid bilayers. J Biol Chem 1999; 274: 29115–29121
- Bechinger B. Towards membrane protein design: pH-sensitive topology of histidine-containing polypeptides. J Mol Biol 1996; 263: 768–775
- Bechinger B, Zasloff M, Opella SJ. Structure and orientation of the antibiotic peptide magainin in membranes by solid-state nuclear magnetic resonance spectroscopy. Protein Sci 1993; 2: 2077–2084
- Patrzykat A, Friedrich CL, Zhang L, Mendoza V, Hancock REW. Sublethal concentrations of Pleurocidin-derived antimicrobial peptides inhibit macromolecular synthesis in Escherichia coli. Antimicrob Agents Chemother 2002; 46: 605–614
- Yonezawa A, Kuwahara J, Fujii N, Sugiura Y. Binding of tachyplesin I to DNA revealed by footprinting analysis: significant contribution of secondary structure to DNA binding and implication for biological action. Biochemistry 1992; 31: 2998–3004
- Park CB, Kim HS, Kim SC. Mechanism of action of the antimicrobial peptide buforin II: buforin II kills microorganisms by penetrating the cell membrane and inhibiting cellular functions. Biochem Biophys Res Commun 1998; 244: 253–257
- Brogden KA. Antimicrobial peptides: pore formers or metabolic inhibitors in bacteria?. Nature Rev Microbiol 2005; 3: 238–250