
ABSTRACT
We document diverse and well-preserved dinoflagellate cyst assemblages from Cretaceous successions in the Kullemölla 1 drill core (640.0 m–590.0 m), Vomb Trough, southern Sweden. Palynology reveals a nearshore marine environment. Dinoflagellate index taxa indicate an Albian to Coniacian age, thus spanning the Early–Late Cretaceous boundary. The lower part of the core is Albian, based on the presence of the index dinocyst taxa Pareodinia and Callaiosphaeridium asymmetricum. The First Appearance Datum (FAD) of Oligosphaeridium prolixispinosum, together with the presence of Achomosphaera ramulifera, Heterosphaeridium difficile and Oligosphaeridium pulcherrimum, reveals a Cenomanian age for the interval 635– m–617 m. The Turonian interval is characterized by an increase in the dinocysts Chatangiella spectabilis and Florentinia spp., in combination with the FAD of Senoniasphaera rotundata, whereas the youngest samples are dated to the Coniacian as defined by the appearance of Glaphyrocysta sp.
We show that Cenomanian and Turonian strata are indeed represented by a relatively condensed section between 635 m and 612 m in the Kullemölla 1 core showing that the apparent hiatus recorded by calcareous microfossils elsewhere is likely a result of post-depositional dissolution of calcareous tests and limestone, a process that did not affect the organic-walled plankton. This is further supported by the presence of hardgrounds and dissolution features.
This updated, detailed biostratigraphical assessment based on dinoflagellates provides a framework for correlation with zonations based on other marine fossil groups, useful, for e.g., correlating aquifers in subsurface successions and, further, provides opportunities for linking marine and continental biotas.
Introduction
The Cretaceous 143.3 Ma (Wimbledon et al. Citation2020a, Citation2020b) to 66 Ma (Renne et al. Citation2018) is characterized by major tectonic events, including the breakup of Pangea, which led to the formation of new continents and ocean basins (Wimbledon et al. Citation2020a, Citation2020b). These major changes in the Earth’s plate tectonics affected climate and biodiversity globally and the period ended with a mass extinction that resulted from an asteroid impact in Yucatan, Mexico (Schulte et al. Citation2010). The Cretaceous is formally divided into the Early Cretaceous (Berriasian–Albian) and the Late Cretaceous (Cenomanian–Maastrichtian), whereas the Lower Cretaceous successions in Europe are mainly characterized by alternating non-marine and marine mudstones, limestones and sandstones (Norling Citation1981, Citation1982; Vajda & Wigforss-Lange Citation2006; Wimbledon Citation2020a, Citation2020b), the Upper Cretaceous is characterized by vast deposits of chalk (Brotzen Citation1936; Hallam Citation1992; Hart et al. Citation2012; Einarsson et al. Citation2016; Fio Firi et al. Citation2017; Khrushchev et al. Citation2022). The Albian–Turonian interval coincided with maximum surface-water temperatures, and the mid-Cretaceous transgression led to the expansion of relatively shallow and warm epicontinental seas (Hallam Citation1992; Larsson et al. Citation2000; Gale et al. Citation2008) that resulted in extensive carbonate deposition during the later part of the Cretaceous (Hallam Citation1992). These chalks were formed primarily by the remains of calcareous phytoplankton, such as coccolithophorids, zooplankton including foraminifera (Brotzen Citation1945; Norling Citation1981; Larsson et al. Citation2000; Slater et al. Citation2022), and calcareous dinoflagellates (belonging to the family Thoracosphaeraceae; Elbrachter et al. Citation2008; Montresor et al. Citation1994; Dubicka & Peryt Citation2012). The tectonic setting that resulted in the formation of sizable shallow shelves, favourable for life in the photic zones along the shorelines, and the generally warm global climate contributed to the proliferation of these groups. In addition, volcanism, a result of the enhanced tectonic activity, contributed significant quantities of greenhouse gases, such as carbon dioxide, into the atmosphere (Vajda & Bercovici Citation2014). Importantly, organic-walled dinoflagellate cysts were abundant in the Cretaceous marine environments, comprising the resting stages of both autotrophs and heterotrophs, and fossil assemblages of these organisms today provide an excellent tool for detailed stratigraphical assessments of these marine Cretaceous successions. The Late Cretaceous continental fringes also hosted the earliest mangroves and seagrasses (McLoughlin et al. Citation2018; van der Ham et al. Citation2017), which resulted in dramatic changes to marginal marine communities, nutrient cycling, and sediment stability. Due to the high productivity in the Cretaceous oceans, the rich nutrient supply from adjacent landmasses, and the extensive populations of planktonic organisms, faunal groups at higher trophic levels also proliferated and diversified during the Cretaceous (Einarsson et al. Citation2016, Einarsson Citation2018). A rich and diverse mollusc fauna existed, and importantly, both belemnites and ammonites underwent rapid evolution and inhabited extensive tracts of the oceans (Christensen Citation1985; Christensen & Schultz Citation1997). When combined with the diverse plankton, their ranges provide a robust biostratigraphical framework for detailed global correlation (Christensen Citation1985; Christensen & Schultz Citation1997). Here, we aim to provide a detailed dinoflagellate cyst biostratigraphy for the mid-Cretaceous successions of the Vomb Trough and to compare this zonation with existing biostratigraphical schemes established for northern Europe.
Geological setting
Mesozoic basin formation on the Baltic Shield was controlled by continental rifting in association with tectonic activity along the Sorgenfrei-Tornquist Zone, a tectonic feature forming a continuous fault zone from Ukraine in SE Europe to the central North Sea (Sorgenfrei & Buch Citation1964; Bergström Citation1984; Larsson et al. Citation2000; Vajda Citation2001; Vajda & Wigforss-Lange Citation2009). The Sorgenfrei-Tornquist Zone forms part of the Fennoscandian Border Zone and represents the border between the East European Craton and the Variscan terranes to the south. Southern Sweden hosts thick Mesozoic successions typified by mainly non-marine Upper Triassic and Jurassic successions, but with marine conditions becoming predominant from the early Late Cretaceous with deposition of glauconitic sandstones and chalks. Cretaceous successions occur throughout the southern province of Skåne () and are represented within several basins; the Båstad Basin, the Höllviken Halfgraben, the Kristianstad Basin, and the Vomb Trough (Larsson et al. Citation2000)
Figure 1. A map outlining the present distribution of Mesozoic strata in southern Sweden. The Kullemölla 1 core is marked with a red dot. Modified after Kruger et al. (Citation2021).
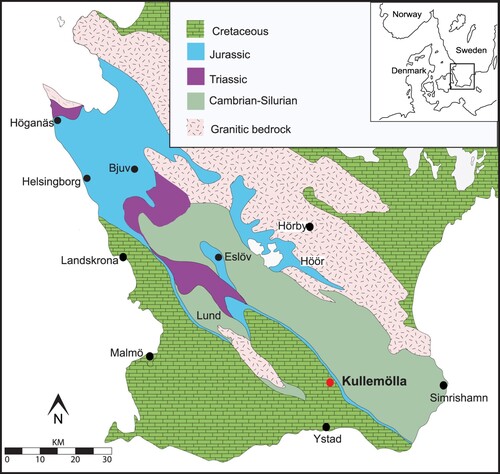
The Vomb Trough initiated during the Triassic as a rift basin (Norling Citation1982; Chatziemmanouil Citation1982; Vajda Citation1998; Lindström & Erlström Citation2011). Maximum sedimentary accumulations occurred in the southern part of the trough, near Köpingsberg, where the succession reaches about 1500 m in thickness (Norling Citation1982; Vajda, Citation1988, Citation1999; Lindström & Erlström Citation2007, Citation2011). The Lower Cretaceous succession is ∼200 m thick and generally comprises sandstones (some glauconitic) and shales; the Upper Cretaceous mostly comprises weakly consolidated carbonate rocks, with some representation of claystones and sandstones. Tectonic activity late in the period led to higher deposition rates, and the Upper Cretaceous successions reached a thickness of ∼1100 m (Bergström et al. Citation1982).
The Kullemölla 1 core was drilled in the southeastern part of the Vomb Trough () and, in 1919, as part of a coal prospecting initiative by the Swedish government targeting the Jurassic coal-bearing strata underlying the Cretaceous sedimentary successions in the area. Lower Cretaceous successions rest unconformably on Lower Jurassic “Liassic” strata, and the boundary between the Jurassic and Cretaceous has been placed previously at 644 m in the Kullemölla 1 core, where it is characterized by a major hiatus (Gaveling Citation1919; Lundegren Citation1935; Guy Ohlson Citation1982). The present study is the first detailed dinocyst biostratigraphic analysis of Early and mid-Cretaceous deposits from Sweden.
Material and Methods
Twenty-four samples were collected for palynological analyses from a 50-metre interval (640 m–590 m) within the Kullemölla 1 drill core. Around 20 g of sedimentary rock was processed according to standard palynological procedures at Global Geolab Limited in Medicine Hat, Canada. The organic residue was sieved using a 5-μm mesh and mounted in epoxy resin on two microscope slides per sample. All dinoflagellate cysts in the two slides per sample were identified taxonomically. Additionally, one kerogen slide was prepared from each sample, i.e., the residue was not exposed to oxidation or sieving processes. Only dinoflagellate cysts have been treated taxonomically in this study, as these are the most valuable for stratigraphic purposes, with several Cretaceous index taxa present. We follow the dinocyst taxonomy of Lentin and Williams (Citation1993) with some more recent amendments (Fensome & Williams Citation2004). Palynomorphs were studied in permanent preparations using an Olympus BX51 biological modular light microscope. The illustrated palynomorphs are identified by NRM numbers and marked with England Finder coordinates. The slides and residues are stored in the Palaeobiology collections of the Department of Palaeobiology at the Swedish Museum of Natural History, Stockholm, Sweden.
Dinoflagellate cyst biostratigraphy
All the studied samples from the Kullemölla 1 drill core yielded well-preserved and abundant dinoflagellate cyst assemblages, and over 100 taxa were identified. The range of selected taxa is presented in , and micrographs of selected dinoflagellate cysts and algae in .
Figure 2. Stratigraphic ranges of selected important dinocysts. Legend 1–3% ![]()
 7–10%
7–10%  .
.
Figure 3. Micrographs of selected dinoflagellate cysts identified in the samples from the Kullemölla 1 drill core. A. Muderongia sp., 614_S202689_S45. B. Muderongia simplex, 637.5_W38. C. Vesperopsis nebulosa, 610.95_M41-3. D. Muderongia sp., 610.95_M41-3. E. Cribroperidinium muderongense, 598_S202703_U36-1. F. Cribroperidinium tenuiceras, 637.5_T29-4. G. Cribroperidinium confossum, 640_O28-4. H. Palaeoperidinium cretaceum, 616_S202711_J27-4. I. Apteodinium sp., 616_S202711_K29. J. Apteodinium granulatum_610.95_L9-4. K. Pareodinia sp., 640_Q15-4. L. Gonyaulacysta sp., 614_S202689_M23-1. M. Leptodinium sp., 637.5_W21-4. N. Chatangiella granulifera, 614_S202689_F17-3. O. Chatangiella sp., 637.5_J32. P. Chatangiella spectabilis, 614_S202689_G25-2. Q. Spinidinium sp., 610.95_D20. R. Endoceratium sp., 592_S202484_U17-4. S, Alterbidinium sp., 590_S202719_H12. T, Trichodinium castanea, 590_S202719_E29. U. Batiacasphaera sp., 614_S202689_D21-1. V. Stephodinium coronatum, 614_S202689_M19-3. W. Dinogymnium sp., 610.95_J14-2. Scale bars = 10 μm.
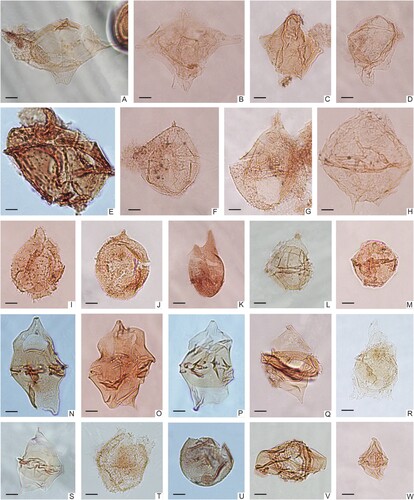
Figure 4. Micrographs of selected dinoflagellate cysts identified in the samples from the Kullemölla 1 drill core. A. Odontochitina operculata, 610.95_H39. B. Odontochitina operculata, 640_E15-3. C. Cyclonephelium filoreticulatum, 598_S202703_G17. D. Leberidocysta sp., 614_S202689_O23-4. E. Canningia sp., 616_S202711_M27-3. F. Senoniasphaera rotundata, 614_S202689_L21. G. Pteridodinium cingulatum, 616_S202711_D28. H. Callaiosphaeridium asymmetricum, 637.5_P20. I. Hystrichosphaeridium tubiferum, 614_S202689_P45-2. J. Florentina laciniata, 590_S202719_E48-2. K. Occisucysta sp., 592_ S202717_P16-2. L. Conosphaeridium striatoconum, 610.95_D37-2. M. Florentinia deanei, 610.95_F36-2. N. Florentinia sp., 590_S202719_K29-4. O. Florentinia ferox, 616_S202711_M27-3. P. Callaiosphaeridium asymmetricum, 592_S202717_H46. Q. Florentinia laciniata, 598_S202703_O25-1. Scale bars = 10 μm.
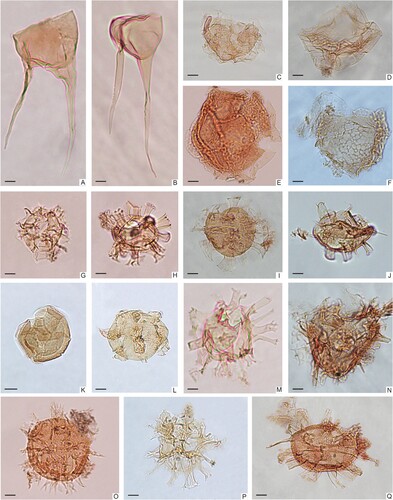
Figure 5. Micrographs of selected dinoflagellate cysts identified in the samples from the Kullemölla 1 drill core. A. Hystrichodinium solare, 640_X15-2. B. Pervosphaeridium pseudhystrichodinium, 610.95_J14-3. C. Hystrichodinium sp., 640_N14-1. D. Exochosphaeridium phragmites, 590_S202719_R13-2. E, Oligosphaeridium pulcherrimum, 640_G19-4. F. Oligosphaeridium complex, 640_T13-2. G. Hystrichoshaeridium bowerbankii, 610.95_T32. H. Spiniferites ramosus, 592_S202717_B28. I. Systematophora sp., 590_S202719_K24-4. J, Cyclonephelium sp., 640_F35-1. K. Circulodinium distinctum, 590_S202719_P28-2. L. Circulodinium distinctum, 598_S202703_V32. M. Glaphyrocysta sp., 610.95_U47-2. N. Achomosphaera ramulifera, 610.95_H13. O. Achomosphaera sagena, 610.95_V36-2. P. Exochosphaeridium sp., 610.95_C27-1. Q. Circulodinium distinctum, 610.95_S25. R. Cleistosphaeridium sp., 640_K45-2. S. Operculodinium sp, 637.5_P20-4. T. Coronifera striolata, 614_S202689_O-22-3. U. Tanyosphaeridium sp., 640_T19-2. Scale bars = 10 μm.
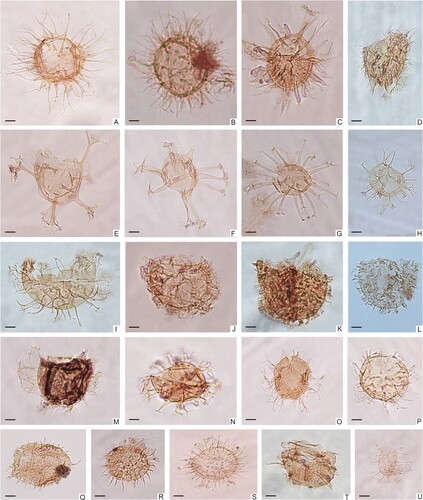
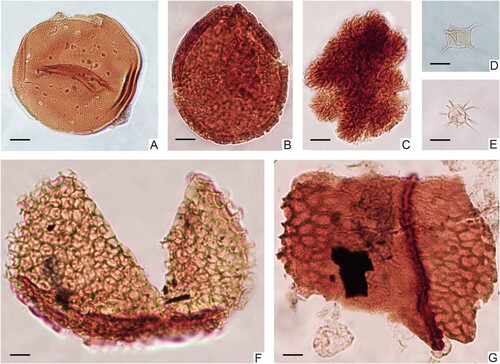
Figure 6. Micrographs of selected acritarchs, prasinophtes, cuticles, and other microfossils from the Kullemölla 1 core. A. Tasmanites sp. 1, 590_S202719_U21-2; B. Tasmanites sp. 2, 640_K39; C. Botryococcus braunii, 640_S22-4; D. Leiofusa filifera, 590_S202719_O25-4; E. Micrhystridium fragile, 610.95_R24; F. Schizosporis reticulatus, 640_T31-2; G. Plant cuticle, 637.5_G37.
Biostratigraphy
Based on dinoflagellate key-taxa, we interpret the studied interval to span the Albian to Coniacian. Reworking is rather extensive at the base of the core.
Interval 640 m–637.5 m, age Albian (samples 640 m and 637.5 m)
The interval 640 m–637.5 m comprises dark grey calcareous siltstones. The dinoflagellate assemblages are diverse and an Albian age is inferred for this interval based on the presence of the age-diagnostic dinocyst taxa (–5) Pareodinia spp., species belonging to the Chatangiella group, and Odontochitina singhii, together with the first occurrence of Callaiosphaeridium asymmetricum (. H, P). Other characteristic dinocysts of this interval include Spiniferites ramosus, Hystrichosphaeridium spp., Cleistosphaeridium spp., Downiesphaeridium sp., Odontochitina operculata, Oligosphaeridium complex and single Cleistosphaeridium polypes, Chytroeisphaeridia sp., Ascodinium sp., Endoscrinium sp., Valensiella sp., Tanyosphaeridium sp., and Sentusidinium sp.
Reworked palynomorphs occur in low numbers <1% represented by 1–2 specimens per taxon throughout the succession. These include typical Early Cretaceous (Berriasian–Barremian) dinocysts, such as Cribroperidinium confossum (G), Cribroperidinium tenuiceras (F), Oligosphaeridium albertense, Muderongia simplex (B), and Scriniodinium sp.
Fresh to brackish-water algae present in this interval include Botryococcus braunii (C), Tasmanites (B), Schizosporis reticulatus (F) and the acritarchs – Micrhystridium fragile and Baltisphaeridium.
Results are compared and contrasted with those from coeval sections elsewhere in the western and eastern Europe (Heilmann-Clausen Citation1987; Tocher & Jarvis Citation1996; Sánchez-Pellicer et al. Citation2018; Shevchuk and Pustovoitova Citation2021).
Interval 635 m–617 m, age Cenomanian (samples 635, 632.6, 629, 626, 624, 622, 620 and 617 m)
The interval 635 m–617 m comprises mainly dark-grey calcareous mudstones. The Cenomanian age of this interval is based on the first occurrences of Achomosphaera ramulifera, Heterosphaeridium difficile, Hystrichosphaeridium bowerbankii, Circulodinium distinctum and Oligosphaeridium prolixispinosum. Taxa dominating in terms of relative abundance include Pervosphaeridium pseudhystrichodinium, Achomosphaera spp., Batiacasphaera spp., Chlamydophorella sp., Chlamydophorella nyei, Oligosphaeridium spp. Pterodinium sp., Spiniferites scabrosus, Spiniferites spp., Hystrichosphaeridium spp., Heterosphaeridium sp., Cleistosphaeridium spp., Downiesphaeridium sp. and Odontochitina spp. Single occurrences of dinocyst taxa in this interval include Systematophora sp., Litosphaeridium sp., Cyclonephelium sp., Hystrichodinium sp., Hystrichodinium solare, Oligosphaeridium pulcherrimum, Trithyrodinium sp., among others ().
Fresh to brackish-water algae present in this interval include Botryococcus, Palambages, Tasmanites and acritarchs – Micrhystridium fragile, M. singulare, Veryhachium reductum, and Baltisphaeridium.
Interval 616 m–612 m, age Turonian (samples 616, 614, 612 m)
The interval 616 m–612 m comprises weakly consolidated, light-grey marlstones. The Turonian assemblages are characterized by an increase in diversity and relative abundance of dinocysts. Florentinia ferox is a zonal taxon, appearing at 616 m, marking the lower Turonian. Senoniasphaera rotundata (appearing at 614 m) is also a zonal species with a First Appearance Datum in the middle Turonian (Ogg et al. Citation2008; Ivanik et al. Citation2012; Shevchuk Citation2016, Citation2018, Citation2020). Heterosphaeridium difficile and Florentinia buspina are also stratigraphically important middle Turonian species within this interval (Radmacher et al. Citation2014). Chatangiella spectabilis, Chatangiella granulifera and single occurrences of the dinocysts Alterbidinium sp., Alterbidinium acutulum are also represented in this interval. The assemblages are dominated by the following taxa: Coronifera oceanica, Chlamidophorella nyei, Spiniferites spp., Spiniferites ramosus, Achomosphaera ramulifera, Hystrichosphaeridium spp., Cleistosphaeridium spp., Oligosphaeridium spp., Membranosphaera sp., Circulodinium distinctum, and Odontochitina spp. Typical taxa that are age-diagnostic for the Turonian include Pterodinium cingulatum, Coronifera striolata, Callaiosphaeridium asymmetricum, Hystrichosphaeridium tubiferum, Xenikoon australis, Stephodinium coronatum, Canningia sp. and Leberidocysta sp. Single occurrences of the dinocysts Trichodinium castanea, Palaeoperidinium cretaceum, Microdinium sp., Microdinium ornatum, Senoniasphaera sp., Prolixosphaeridium sp., and Pyxidiella sp. occur in this interval along with the last occurrence of Gonyaulacysta sp. Examples of reworked dinocysts include Apteodinium spp., Apteodinium granulatum, Apteodinium maculatum, and Sirmiodinium sp. ().
Fresh-brackish water algae present in this interval include Botryococcus, Palambages, Tasmanites, and the acritarch taxa Micrhystridium fragile.
Interval 610.95 m–590 m, age Coniacian (samples 610.95, 608, 606, 604.5, 602, 600, 598, 596, 594, 592, and 590 m)
The interval 610.95 m–590 m comprises mainly light grey mudstones. This Coniacian interval is characterized by the first appearance of the dinocysts Glaphyrocysta sp. (M) and Dinogymnium sp. (W). Other typical Coniacian index taxa include the dinocysts Achomosphaera sagena, Hystrichodinium pulchrum, Pterodinium spp., Pterodinium pterotum, Pterodinium cingulatum, Cauveridinium membraniphorum, Amphidiadema sp., Conosphaeridium striatoconum, Odontochitina costata, and Odontochitina operculata. Cleistosphaeridium spp. and Downiesphaeridium sp. show little variation in relative abundance throughout the section. An acme of Spiniferites spp. and Spiniferites ramosus is noted in this interval. Single occurrences of stratigraphically important dinocysts include Exochosphaeridium phragmites, Exochosphaeridium sp., Cyclonephelium filoreticulatum, Trichodinium castanea, Spinidinium sp., Occisucysta sp., Cribroperidinium sp., Cribroperidinium edwardsii, Cribroperidinium muderongense, Hystrichodinium sp., Oligosphaeridium pulcherrimum, Operculodinium sp., Ginginodinium spinulosum, Florentinia laciniata, Florentinia deanei, and Tehamadinium coummia (). There are also numerous Kallosphaeridium sp. and Pervosphaeridium pseudhystrichodinium.
Fresh to brackish-water algae present in this interval include Shizosporis reticulatus, Tasmanites, Zygnemaceae and acritarchs – Leiofusa filifera (D) and Micrhystridium fragile (E), Veryhachium reductum, Baltisphaeridium.
Remains of plant cuticles and tracheids are found uniformly throughout all intervals (G).
Discussion and conclusions
We have provided a detailed palynostratigraphy for the successions from the Vomb Trough, accessed through the Kullemölla 1 core, southern Sweden. This study significantly expands on pioneer palynological investigations (Guy-Ohlson Citation1982) concerning both taxonomic richness and refined stratigraphy across the Lower–Upper Cretaceous boundary in Skåne. We show that the 50 m sedimentary succession within the Kullemölla 1 core reflects a relatively condensed interval spanning the Albian to Coniacian, thus representing a time interval of about 25 million years, indicating a sedimentation rate of 2 m/million years or 2 millimetres per 1000 years, which signifies a slow depositional rate. The studied assemblages can further be placed in a broader biostratigraphical context and linked to dinoflagellate, ammonite and foraminifera zonations from elsewhere in the Boreal and Tethys regions ().
Figure 7. Overview of Cretaceous zonation scheme, modified after Ogg et al., Citation2008.

Based on the dinoflagellate assemblages, the environment was marine during the entire time of deposition in this part of the Vomb Trough. In fact, sea levels were rising throughout this time interval, as previously demonstrated by others (Hart Citation1980; Solakius and Larsson Citation1985; Larsson et al. Citation2000). Important findings herein include the identification of species belonging to the Chatangiella dinocyst complex very low in the studied interval (within the Albian successions). Importantly, Glaphyrocysta was also recorded within the Coniacian part of the analyzed succession. Elsewhere these genera generally characterize the Coniacian–Santonian (Westin Citation1992; Riding and Crame Citation2002; Shevchuk and Vajda Citation2014). Another notable observation is the presence of Dinogymnium dinocysts in the Coniacian interval of the Kullemölla 1 core. The Dinogymnium complex is an important component of Upper Cretaceous dinoflagellate cyst assemblages from marine palaeoenvironments worldwide (Evitt et al. Citation1967).
Previous studies from Cretaceous basins in southern Sweden have revealed a complex depositional history, with a hiatus across the Cenomanian–Turonian boundary (Solakius and Larsson Citation1985; Larsson et al. Citation2000; Hart et al. Citation2012; Ezampanah et al. Citation2021). According to Hart et al. (Citation2012) upper Cenomanian to lower Coniacian strata are lacking in the basin altogether, with no evidence of Turonian microfossils such as calcareous plankton. In other parts of the European Platform, such as in Ukraine, there is evidence of blooms of calcareous dinoflagellate cysts with an acme during the Albian to the Turonian (Dias Brito Citation2000; Ciurej et al. Citation2023), also coinciding with the highest recorded Cretaceous seawater surface temperatures (Ciurej et al. Citation2023). Around the Cenomanian–Turonian boundary, a sea level rise reached a maximum for the Cretaceous, estimated to have reached ∼ 250 m above the present-day maximum (Haq Citation2014). After reaching its Cretaceous maximum, sea level fell by the end of the Turonian, remaining relatively stable for almost 20 Myr during the Coniacian and the Campanian (Haq Citation2014).
Based on organic-walled dinoflagellate cysts we show that Cenomanian and Turonian strata are indeed represented by a relatively condensed section between 635 and 612 m in the Kullemölla 1 core. The diversity of cysts is characteristic of the Cenomanian / Turonian boundary interval, which is typified by the dominance of tolerant cosmopolitan forms, such as Circulodinium distinctum, Hystrichosphaeridium bowerbankii, Oligosphaeridium complex, Odontochitina costata, and O. operculata (Bruce and Jarvis Citation1995; Shevchuk Citation2020). Turonian assemblages are characterized by diverse Florentinia spp. dinocysts and important Senoniasphaera rotundata and Heterosphaeridium difficile. The Coniacian is characterized by the first appearance of the dinocysts genera Glaphyrocysta and Dinogymnium. This indicates that the apparent hiatus recorded from calcareous microfossils and strata was likely the result of post-depositional dissolution of carbonate tests, which did not affect organic-walled remains.
Post-depositional dissolution and consequent mixing of fossil assemblages complicate the stratigraphical assessment. The complex lithologies and slow depositional rates are further evidenced by the many hardgrounds, intensely bioturbated intervals, formation of glauconitic sands, and phosphatization (Vajda et al. Citation1999; Larsson et al. Citation2000; Hart et al. Citation2012). These factors have resulted in reworking or alteration of faunal remains hampering the identification of discrete events (Hart et al. Citation2012).
An example of reworking within the studied dinoflagellate assemblages is the record of Muderongia simplex and other single occurrences within the transgressive bed at the base of the core at 637.5 m. These species are characteristic of successions not younger than Valanginian (Riding et al. Citation2000); however, in the base of the Kullemölla 1 core they co-occur with large numbers of Odontochitina operculata typical of Albian assemblages (Ogg et al. Citation2008).
Dinocyst biozonation provides a high-resolution means of dating marine strata in southern Sweden that can be useful for correlating aquifers in subsurface successions, along with evaluating the rates of subsidence and sea-level fluctuations in the Cretaceous. Spore-pollen zonations and carbon isotope data from the studied marine successions of southern Sweden are at a preliminary stage but future work on these terrestrial microfossil assemblages will provide an opportunity to link the neritic and continental biozonations and the global carbon isotope stratigraphy.
Acknowledgements
Funding is acknowledged from the Swedish Foundation for Strategic Research, grant UKR22-0020 (O. Shevchuk). We are further thankful to the Knut & Alice Wallenberg Foundation KAW 2020.0145 (V. Vajda), and the Swedish Research Council VR grants 2019–4061 (V. Vajda). Olena Shevchuk wishes to thank the Swedish Government, the Swedish Museum of Natural History, and Stockholm University for hosting her under the Temporary Protection Directive, following the Russian invasion of Ukraine. We thank the reviewers for their constructive comments.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Bergström, J., 1984: Lateral movements in the Tornquist Zone. Geologiska Föreningens i Stockholm Förhandlingar 106, 379–380. doi: 10.1080/11035898509454667
- Bergström, J., Holland, B., Larsson, K., Norling, K. & Sivhed, U., 1982: Guide to excursions in Scania. Sveriges Geologiska Undersökning Ca 54, 1–95.
- Brotzen, F., 1936: Foraminiferen aus dem schwedischen untersten Senon von Eriksdal in Schonen. Sver. Geol. Unders. C 396, 206.
- Brotzen, F., 1945: De geologiska resultaten från borrningarna vid Höllviken (in Swedish with English summary). Sveriges Geologiska Undersökning ser. C 465, 1–64.
- Bruce, A.T. & Jarvis, I., 1995: Dinocyst distributions and stratigraphy of two Cenomanian–Turonian boundary (Upper Cretaceous) sections from the western Anglo-Paris Basin. Journal of Micropalaeontology 14(2), 97–105. DOI: 10.1144/jm.14.2.97.
- Chatziemmanouil, J., 1982: The Upper Cretaceous of the Vomb Trough. Part I: Structure, geology and sedimentology. Stockholm Contribution in Geology 38(5), 57–112, Part II: Foraminiferal palaeoecology of the Coniacian and Santonian sequences and its applications. Stockholm Contribution in Geology 38(6), 113–161.
- Christensen, W.K., 1985: The Albian to Maastrichtian of southern Sweden and Bornholm, Denmark: a review. Cretaceous Research 5, 313–327. doi: 10.1016/S0195-6671(84)80027-0
- Christensen, W.K. & Schultz, M.-G., 1997: Coniacian and Santonian belemnite faunas from Bornholm, Denmark. Fossils and Strata 44, 1–73. doi: 10.18261/8200376958-1997-01
- Ciurej, A., Dubicka, Z. & Poberezhskyy, A., 2023: Calcareous dinoflagellate blooms during the Late Cretaceous ‘green-house’ world – a case study from western Ukraine. PeerJ 11, e16201. doi: 10.7717/peerj.16201
- Dias-Brito, D., 2000: Global stratigraphy, palaeobiogeography and palaeoecology of Albian–Maastrichtian pithonellid calcispheres: impact on Tethys configuration. Cretaceous Research 21, 315–349. DOI: 10.1006/cres.2000.0196.
- Dubicka, Z. & Peryt, D., 2012: Foraminifers and stable isotope record of the Dubivtsi chalk (upper Turonian, Western Ukraine): palaeoenvironmental im plication. Geological Quarterly 56(1), 199–214.
- Einarsson, E., 2018: Palaeoenvironments, palaeoecology and palaeobiogeography of Late Cretaceous (Campanian) faunas from the Kristianstad Basin, southern Sweden, with applications for science education. Litholund Thesis 32. Lund University, Lund. 248.
- Einarsson, E., Praszkier, A. & Vajda, V., 2016: First evidence of the Cretaceous decapod crustacean Protocallianassa from Sweden. Geological Society, London, Special Publications 434(1), 241–250. doi.org/10.1144/SP434.6.
- Elbrachter, M., Gottschling, M., Hildebrand-Habel, T., Keupp, H., Kohring, R., Lewis, J., Meier, K.J.S., Montresor, M., Streng, M., Versteegh, G.J.M., Willems, H. & Zonneveld, K., 2008: Establishing an Agenda Calcareous Dinoflagellate Research (Thoracosphaeraceae, Dinophyceae) including a nomenclatural synopsis of generic names. Taxon. 57(4), 1289–1303. doi: 10.1002/tax.574019
- Evitt, W.R., Clarke, R.F.A. & Verdier, J.-P., 1967: Dinoflagellate studies III. Dinogymnium acuminatum n. gen., n.sp. (Maastrichtian) and other fossils formerly referable to Gymnodinium Stein. Stanford University Publications, Geological Sciences 10(4), 1–27, pl.1–3.
- Ezampanah, Y., Scopelliti, G., Sadeghi, A., Adabi, M.H., Jamali, A.M., Caruso, A., Mohseni, H. & Razmjooei, M.J., 2021: Turonian–Maastrichtian biostratigraphy and isotope stratigraphy of the Kopet-Dagh Basin deposits, northeastern Neo-Tethys, Iran. Palaeogeography, Palaeoclimatology, Palaeoecology 581, 110605. doi: 10.1016/j.palaeo.2021.110605
- Fensome, R.A. & Williams, G.L., 2004: The Lentin and Williams index of fossil dinoflagellates: Edition. AASP Contributions 42, 1–824.
- Fio Firi, K., Bercovici, A., Shevchuk, O. & Sremac, J., 2017: Late Cretaceous palynoflora from the central part of the Adriatic Carbonate Platform (Islands of Hvar and Šćedro), southern Croatia. Cretaceous Research 74, 142–150. DOI: 10.1016/j.cretres.2017.02.003.
- Gale, A.S., Voigt, S., Sageman, B.B. & Kennedy, W.J., 2008: Eustatic sea-level record for the Cenomanian (Late Cretaceous) – extension to the Western Interior Basin, USA. Geology 36(11), 859–862. DOI: 10.1130/G24838a.1.
- Gavelin, A., 1919: Ett preliminärt meddelande om resultaten af djupborrningarna efter stenkol i Skåne. (Referat av ett hållet föredrag). Geologiska Föreningens i Stockholm Förhandlingar 41, 223–224.
- Guy-Ohlson, D., 1982: Biostratigraphy of the Lower Jurassic–Cretaceous unconformity at Kullemölla Southern Sweden. Sver. Geol. Unders 52, 1–45.
- Hallam, A., 1992: Phanerozoic Sea-Level Changes. Columbia University Press, New York. 266.
- Haq, B.U., 2014: Cretaceous Eustasy Revisited. Global Planetary Change 113, 44–58. http://doi.org/10.1016/j.gloplacha.2013.12.007.
- Hart, M.B., 1980: The recognition of mid-Cretaceous sea level changes by means of Foraminifera. Cretaceous Research 1, 289–297. doi: 10.1016/0195-6671(80)90040-3
- Hart, M.B., Bromley, R.G. & Packer, S.R., 2012: Anatomy of the stratigraphical boundary between the Arnager Greensand and Arnager Limestone (Upper Cretaceous) on Bornholm, Denmark. Proceedings of the Geologists Association 123(3), 471–478. doi: 10.1016/j.pgeola.2011.11.006
- Heilmann-Clausen, C., 1987: Lower Cretaceous dinoflagellate biostratigraphy in the Danish Central Trough. Danmarks Geologiske Undersogelse, A 17, 1–89. doi: 10.34194/seriea.v17.7036
- Ivanik, M.M., Shevchuk, O.A. & Klymenko, Y.V., 2012: Biostratigraphic characteristics of the Albian–Turonian deposits of the southwestern slope of the Ukrainian shield and the parastratotype of the Rusavska suite. Tectonics and Stratigraphy 39, 87–95. (in Ukranian). DOI:10.30836/igs.0375-7773.2012.93101
- Khrushchev, D.P., Remezova, O.O., Vasylenko, S.P., Shevchuk, O.A. & Yaremenko, O.V., 2022: Digital structural-lithological model and geological-genetic characteristics of the Kopytkiv phosphorite deposit. Bulletin of Kharkiv National University Named After V.N. Karazina, Series “Geology. Geography. Ecology” 56, 88–104, in Ukrainian. doi: 10.26565/2410-7360-2022-56-06
- Krüger, A., Slater, S. & Vajda, V., 2021: 3D imaging of shark egg cases (Palaeoxyris) from Sweden with new insights into Early Jurassic shark ecology 143(2–3), 229–247. DOI:10.1080/11035897.2021.1907442.
- Larsson, K., Solakius, N. & Vajda-Santivanez, V., 2000: Foraminifera and palynomorphs from the green sand-limestone sequences (Aptian–Coniacian) in south western Sweden. Neues Jahrbuch fur Geologie und Palaontologie 216, 277–312. doi: 10.1127/njgpa/216/2000/277
- Lentin, J.K. & Williams, G.L., 1993: Fossil dinoflagellates: index to genera and species 1993 edition. American Association of Stratigraphic Palynologists Contribution 28, 1–856.
- Lindström, S. & Erlström, M., 2007: Dating and correlating potential aquifers for geothermal energy, CO2-, and energy storage, within the late Triassic–early Cretaceous succession in the Swedish part of the Danish Basin. 1–47.
- Lindström, S. & Erlström, M., 2011: The Jurassic–Cretaceous transition of the Fårarp-1 core, southern Sweden: sedimentological and phytological indications of climate change. Palaeogeography, Palaeoclimatology, Palaeoecology 308(3–4), 445–475. doi: 10.1016/j.palaeo.2011.05.052
- Lundegren, A., 1935: Die Stratigraphi schem Ergebnisse der Tiefbohrung bei Kullemölla im Sudöstlichen Schonen. Sver. Geol. Unders 386, 1–19.
- McLoughlin, S., Haig, D.W., Siversson, M. & Einarsson, E., 2018: Did mangrove communities exist in the Late Cretaceous of the Kristianstad Basin, Sweden? Palaeogeography, Palaeoclimatology, Palaeoecology 498, 99–114. doi: 10.1016/j.palaeo.2018.03.007
- Montresor, M., Montesarchio, E., Marino, D. & Zingone, A., 1994: Calcareous dinoflagellate cysts in marine sediments of the Gulf of Naples (Mediterranean Sea). Review of Palaeobotany and Palynology 84(1–2), 45–56. doi: 10.1016/0034-6667(94)90040-X
- Norling, E., 1981: Upper Jurassic and Lower Cretaceous geology of Sweden. Geol. Fören. Stockh. Förh. 103, 253–269. doi: 10.1080/11035898109454522
- Norling, E., 1982: Eriksdal (and adjacent Kurremölla valley). In J. Bergström, B. Holland, K. Larsson, E. Norling & U. Sivhed (eds.): Guide to excursions in Scania. Sver. geol. unders. Ca 54, 75–81. Uppsala: Swedish Geological Survey.
- Ogg, J.G., Ogg, G. & Gradstein, F. M. 2008. The Concise Geologic Time Scale. 177. Cambridge, New York, Melbourne: Cambridge University Press.
- Radmacher, W., Tyszkaa, J., Mangerud, G. & Pearce, M.A., 2014: Dinoflagellate cyst biostratigraphy of the Upper Albian to Lower Maastrichtian in the southwestern Barents Sea. Marine and Petroleum Geology 57, 109–121. doi: 10.1016/j.marpetgeo.2014.04.008
- Renne, P.R., Arenillas, I., Arz, J.A., Vajda, V., Gilabert, V. & Bermúdez, H.D., 2018: Multi-proxy record of the Chicxulub impact at the Cretaceous-Paleogene boundary from Gorgonilla Island, Colombia. Geology 46(6), 547–550. doi: 10.1130/G40224.1
- Riding, J.B. & Crame, J.A., 2002: Aptian to Coniacian (Early–Late Cretaceous) palynostratigraphy of the Gustav Group, James Ross Basin, Antarctica. Cretaceous Research 23, 739–760. doi: 10.1006/cres.2002.1024
- Riding, J.B., Poulsen, N.E. & Bailey, D.A., 2000: A taxonomic study of the dinoflagellate cyst Muderongia simplex Alberti 1961 and related species. Palynology 24, 21–35. doi: 10.2113/0240021
- Sánchez-Pellicer, R., Masure, E. & Villier, L., 2018: Distribution of Albian dinoflagellate cyst associations along a proximal–distal transect across the Iberian margin. Cretaceous Research 92, 240–256. doi: 10.1016/j.cretres.2018.08.004
- Schulte, P., Alegret, L., Arenillas, I., Arz, J.A., Barton, P.J., Bown, P.R., Bralower, T.J., Christeson, G.L., Claeys, P., Cockell, C.S., Collins, G.S., Deutsch, A., Goldin, T.J., Goto, K., Grajales-Nishimura, J.M., Grieve, R.A.F., Gulick, S.P., Johnson, K.R., Kiessling, W., Koeberl, C., Kring, D.A., MacLeod, K.G., Matsui, T., Melosh, J., Montanari, A., Morgan, J.V., Neal, C.R., Nichols, D.J., Norris, R.D., Pierazzo, E., Ravizza, G., Rebolledo-Vieyra, M., Uwe Reimold, W., Robin, E., Salge, T., Speijer, R.P., Sweet, A.R., Urrutia-Fucugauchi, J., Vajda, V., Whalen, M.T. & Willumsen, P.S., 2010: The chicxulub asteroid impact and mass extinction at the cretaceous–paleogene boundary. Science 327(5970), 1214–1219. DOI: 10.1126/science.1177265.
- Shevchuk, O., 2016: New data on the Zonal scale of the Albian–Campanian biostratigraphic subdivisions of the western regions of platform Ukraine. Materials of the Session of the Paleontological Society of the National Academy of Sciences of Ukraine. Kyiv. XXXVII, 55–57, in Ukranian.
- Shevchuk, O., 2018: Zonal scales of biostratigraphic subdivisions of the middle, upper Jurassic and cretaceous of Ukraine by dinocysts. Materials of the IX All-Ukrainian scientific conference “Problems of Phanerozoic geology of Ukraine”. Lviv. 13–21. (in Ukranian)
- Shevchuk, O., 2020: Stratigraphy of the Middle Jurassic – Cretaceous of Ukraine according to microfossils. Dissertation abstract. 44. (in Ukranian).
- Shevchuk, O. & Pustovoitova, D., 2021: Stratification of Cretaceous sediments of the section of well No. 29 – Zahidno-Oktyabrska (Karkinitsko-North Crimean depression) according to palynological data. Bulletin of Taras Shevchenko Kyiv National University Series Geology 3(94), 27–36, (in Ukranian).
- Shevchuk, O. & Vajda, V., 2014: Taxonomic diversity of dinoflagellates in the Cretaceous seas traced in the Crimean Mountains. Materials of the III International Scientific and Practical Conference “Biodiversity and Sustainable Development”. 391–392.
- Slater, S.M., Bown, P., Twitchett, R.J., Danise, S. & Vajda, V., 2022: Global record of “ghost” nannofossils reveals plankton resilience to high CO2 and warming. Science 376, 853–856. DOI: 10.1126/science.abm7330.
- Solakius, N. & Larsson, K., 1985: Foraminifera and Biostratigraphy of the Arnager Limestone. Bornholm, Denmark: Danmarks Geologiske Undersøgelse Serie C, 5, 1–42. DOI:10.34194/seriec.v5.7099
- Sorgenfrei, T. & Buch, A., 1964: Deep tests in Denmark 1935–1959. Danmarks Geologiske Undersøgelse III. Række 36, 1–146. doi: 10.34194/raekke3.v36.6941
- Tocher, B.A. & Jarvis, I., 1996: Dinoflagellate cyst distributions and the Albian–Cenomanian boundary (mid-Cretaceous) at Cordebugle, NW France and Lewes, southern England. Journal of Micropalaeontology 15, 55–67. doi: 10.1144/jm.15.1.55
- Vajda, V., 1988: Biostratigrafisk indelning av den mesozoiska lagerföljden i Köpingsbergsborrningen 2, Skåne. BSc Thesis, Lund University 25, 1–13. (in Swedish).
- Vajda, V., 1998: Cretaceous palynofloras of southwestern Scandinavia. PhD thesis. Lund Publication in Geology. No 135, 1–63.
- Vajda, V., 1999: Miospores from Upper Cretaceous–Paleocene strata in northwestern Bolivia. Palynology 23, 183–198.
- Vajda, V., 2001: Aalenian to Cenomanian terrestrial palynofloras of SW Scania, Sweden. Acta Palaeontologica Polonica 46(3), 403426.
- Vajda, V. & Bercovici, A., 2014: The global vegetation pattern across the Cretaceous–Paleogene mass-extinction interval – an integrated global perspective. Global and Planetary Change 12, 29–49. doi: 10.1016/j.gloplacha.2014.07.014
- Vajda, V. & Solakius, N., 1999: Palynomorphs, foraminifera and calcisphaeres from the greensand-limestone transition at Arnager, Bornholm: evidence for the late Cenomanian to early Coniacian transgression. GFF 12, 281–286. doi: 10.1080/11035899901214281
- Vajda, V. & Wigforss-Lange, J., 2006: The Jurassic–Cretaceous transition of Southern Sweden – palynological and sedimentological interpretation. Progress in Natural Science 16, 1–38. doi: 10.1080/10020070612331343185
- Vajda, V. & Wigforss-Lange, J., 2009: Onshore Jurassic of Scandinavia and related areas. GFF 131, 5–23. doi: 10.1080/11035890902975309
- van der Ham, R.W.J.M., Konijnenburg-van Cittert, J.H.A., Jagt, J.W.M., Indeherberge, L., Meuris, R., Deckers, M.J.M., Renkens, S. & Laffineur, J., 2017: Seagrass stems with attached roots from the type area of the Maastrichtian stage (NE Belgium, SE Netherlands): morphology, anatomy, and ecological aspects. Review of Palaeobotany and Palynology 241, 49–69.
- Westin, H. 1992: Cretaceous dinoflagellate cyst stratigraphy of the Hollvikenl well, Scania, Southern Sweden. Doctoral Dissertation 1992. Department of Geology, University of Stockholm. 175.
- Wimbledon, W.A.P., Rehakova, D., Svobodova, A., Elbra, T., Schnabl, P., Pruner, P., Šifnerová, K., Kdýr, Š, Dzyuba, O., Schnyder, J., Galbrun, B., Košťák, M., Vaňková, L., Copestake, P., Hunt, C.O., Riccardi, A., Poulton, T.P., Bulot, L.G., Frau, C. & de Lena, L.F., 2020a: The proposal of a GSSP for the Berriasian Stage (Cretaceous System): part 1. Volumina Jurassica XVIII(1), 53–106. doi: 10.7306/vj.18.5
- Wimbledon, W.A.P., Rehakova, D., Svobodova, A., Elbra, T., Schnabl, P., Pruner, P., Šifnerová, K., Kdýr, Š, Frau, C., Schnyder, J., Galbrun, B., Vaňková, L., Dzyuba, O., Copestake, P., HUNT, C.O., Riccardi, A., Poulton, T.P., BULOT, L.G. & de Lena, L.F., 2020b: The proposal of a GSSP for the Berriasian Stage (Cretaceous System): Part 2. Volumina Jurassica XVIII, 119–157. doi: 10.7306/vj.18.5
Appendix
List of dinocyst species and genera recorded from the Kullemölla 1 drill core. The taxa are arranged alphabetically by genus and species:
Achomosphaera ramulifera (Deflandre 1937b) Evitt, 1963
Achomosphaera sagena Davey & Williams, 1966a
Achomosphaera sp.
Alterbidinium sp.
Alterbidinium acutulum (Wilson, 1967b) Lentin and Williams, 1985
Amphidiadema sp.
Apteodinium sp.
Apteodinium granulatum Eisenack 1958a
Apteodinium maculatum Eisenack & Cookson, 1960
Ascodinium sp.
Batiacasphaera sp.
Callaiosphaeridium asymmetricum (Deflandre and Courteville, 1939) Davey and Williams, 1966b
Canningia sp.
Cauveridinium membraniphorum (Cookson and Eisenack, 1962b)
Conosphaeridium striatoconum (Deflandre and Cookson 1955) Cookson and Eisenack 1969
Chatangiella sp.
Chatangiella granulifera (Manum, 1963) Lentin and Williams, 1976
Chatangiella spectabilis (Alberti, 1959b) Lentin and Williams, 1976
Chlamydophorella sp.
Chlamydophorella nyei Cookson and Eisenack, 1958
Chytroeisphaeridia sp.
Circulodinium distinctum (Deflandre and Cookson, 1955) Jansonius, 1986
Cleistosphaeridium polypes (Cookson and Eisenack, 1962b) Davey, 1969a
Cleistosphaeridium sp.
Coronifera oceanica Cookson and Eisenack, 1958
Coronifera striolata (Deflandre, 1937b) Stover and Evitt, 1978
Cribroperidinium sp.
Cribroperidinium confossum (Duxbury, 1977) Helenes, 1984
Cribroperidinium edwardsii (Cookson and Eisenack, 1958) Davey, 1969a
Cribroperidinium tenuiceras (Eisenack, 1958a) Poulsen, 1996
Cribroperidinium muderongense (Cookson and Eisenack, 1958) Davey, 1969a
Cyclonephelium sp.
Cyclonephelium filoreticulatum (Slimani, 1994) Prince et al., 1999
Dinogymnium sp.
Downiesphaeridium sp.
Endoscrinium sp.
Exochosphaeridium sp.
Exochosphaeridium phragmites Davey et al., 1966
Florentinia sp.
Florentinia deanei (Davey and Williams, 1966b) Davey and Verdier, 1973
Florentinia ferox (Deflandre, 1937b) Duxbury, 1980
Florentinia laciniata Davey and Verdier, 1973
Florentinia buspina (Davey and Verdier, 1976)
Ginginodinium spinulosum Cookson and Eisenack, 1960a
Glaphyrocysta sp.
Gonyaulacysta sp.
Heterosphaeridium sp.
Heterosphaeridium difficile (Manum and Cookson, 1964) Ioannides, 1986
Hystrichodinium sp.
Hystrichodinium pulchrum Deflandre, 1935
Hystrichodinium solare Pestchevitskaya, 2009
Hystrichosphaeridium bowerbankii Davey and Williams, 1966b
Hystrichosphaeridium tubiferum (Ehrenberg, 1837b) Deflandre, 1937b
Hystrichosphaeridium sp.
Kallosphaeridium sp.
Leberidocysta sp.
Leptodinium sp.
Litosphaeridium sp.
Membranosphaera Samoilovitch and Mtchedlishvili, 1961
Microdinium sp.
Microdinium ornatum Cookson and Eisenack, 1960a
Muderongia sp.
Muderongia simplex Alberti, 1961
Occisucysta sp.
Odontochitina sp.
Odontochitina costata Alberti, 1961
Odontochitina operculata (Wetzel, 1933a) Deflandre and Cookson, 1955
Odontochitina singhii Morgan, 1980
Oligosphaeridium albertense (Pocock, 1962) Davey and Williams, 1969
Oligosphaeridium complex (White, 1842) Davey and Williams, 1966b
Oligosphaeridium prolixispinosum Davey and Williams, 1966b
Oligosphaeridium pulcherrimum Deflandre and Cookson, 1955
Oligosphaeridium sp.
Operculodinium sp.
Palaeoperidinium cretaceum (Pocock, 1962 ex Davey, 1970) Lentin and Williams, 1976
Pareodinia sp.
Pervosphaeridium pseudhystrichodinium (Deflandre, 1937b) Yun Hyesu, 1981
Prolixosphaeridium sp.
Pterodinium sp.
Pterodinium cingulatum (Wetzel, 1933b) Below, 1981a
Pterodinium pterotum (Cookson and Eisenack) Pavlishina, 1990
Pyxidiella sp.
Scriniodinium sp.
Senoniasphaera sp.
Senoniasphaera rotundata Clarke and Verdier, 1967
Sentusidinium sp.
Sirmiodinium sp.
Spinidinium sp.
Spiniferites ramosus (Ehrenberg, 1837b) Mantell, 1854
Spiniferites scabrosus (Clarke and Verdier, 1967) Lentin and Williams, 1975
Spiniferites sp.
Stephodinium coronatum Deflandre, 1936a
Systematophora sp.
Tanyosphaeridium sp.
Tehamadinium coummia (Below, 1981a) Jan du Chêne et al., 1986b
Trichodinium castanea Deflandre, 1935 ex Clarke and Verdier, 1967
Trithyrodinium sp.
Valensiella sp.
Vesperopsis nebulosa Bint, 1986
Xenikoon australis Cookson and Eisenack, 1960a
Green algae
Botryococcus braunii Kutzing, 1849
Palambages sp.
Shizosporis reticulatus Cookson & Dettmann 1959 emend. Pierce 1976
Tasmanites sp.
Zygnemaceae sp.
Аcritarchs
Baltisphaeridium sp.
Cymatiosphaera sp.
Micrhystridium fragile Deflandre, 1947
Micrhystridium singulare Firtion, 1952
Leiofusa filifera Downie, 1959
Veryhachium reductum Deunff, 1958
Other HF-resistent organic matter
Tracheids
Cuticles
Structured wood fragments
Opaque phytoclast
Microforaminifera