Abstract
The red marine alga (Polysiphonia virgata. C. Agardh; Rhodomelaceae) was investigated for antimycobacterial activity. Bioassay-guided fractionation of the dichloromethane algal extract using direct bioautography resulted in isolation of a mixture of long-chain fatty acids, namely oleic acid, linoleic acid, lauric acid, and myristic acid as the major antimycobacterial compounds. Oleic acid showed the greatest inhibition of the growth of Mycobacterium smegmatis. with a minimum inhibitory quantity (MIQ) of 0.8 μ g; linoleic acid and lauric acid had MIQ values of 1.56 and 3.125 μ g, respectively. Stearic acid, palmitic acid, and myristic acid did not inhibit the growth of M. smegmatis.. Using the Bactec-460 radiometric method, oleic acid showed 100% inhibition of the growth of Mycobacterium tuberculosis. at a minimum inhibitory concentrations (MIC) of 25 μ g/mL; lauric acid, myristic acid, and linoleic acid all showed 100% inhibition at MIC values of 50 μ g/mL. Myristic acid and lauric acid showed 90% and 76% inhibition at 50 μ g/mL. Linoleic acid showed moderate inhibition of the growth of a clinical strain of multidrug-resistant M. tuberculosis. 50 μ g/mL.
Introduction
South Africa is among the 22 countries that bear 80% of the global tuberculosis (TB) case burden (Raviglione, Citation2002). The 2004 WHO estimates showed that the population of 44,759,187 had a TB case incidence of 558/100,000 population, with 60% of adult cases being co-infected with HIV (WHO Report, 2004). In the treatment of HIV infection, protease inhibitors (PIs) and non-nucleoside reverse transcriptase inhibitors (NNRTIs) form an integral part of the multi-drug regimens in AIDS patients (Fujiwara et al., Citation2000). However, these medications have serious consequences for the treatment of tuberculosis. They interact with rifamycins, the most important chemotherapeutic class of drugs in the treatment of tuberculosis, and particularly with rifampicin. Current tuberculosis drugs must be taken during at least 6 months and in combination to minimize the chances of multidrug-resistant tuberculosis (MDR-TB) developing (Kaufmann, Citation2000). The adverse reactions experienced by many patients lead to poor adherence and treatment failure (O'Brien, Citation2001). The drugs used to treat MDR-TB are expensive and more toxic than the first-line drugs (O'Brien, Citation2001). New TB-drug treatments are needed that would shorten the treatment period, be effective against MDR-TB, eliminate latent TB infection (O'Brien, Citation2001), and be affordable to all.
Marine natural products are increasingly receiving attention in the search for new and effective medicinal compounds. This is reflected in the many investigations into their bioactivities such as antiangiogenic (Koyanagi et al., Citation2003), antitumor (Hiroishi et al., Citation2001; Zhou et al., Citation2004), anticoagulant (Chevolot et al., Citation1999), immunomodulating (Shan et al., Citation1999), antioxidant (Ahn et al., Citation2004; Zhang et al., Citation2004), antiviral (Boyd et al., Citation1997; Carlucci et al., Citation1999; Huheihel et al., Citation2002; Ahn et al., Citation2002), anti-inflammatory (Jiang et al., Citation1999; Okai & Higashi-Okai, Citation1997), antimicrobial (Donia & Hamann, Citation2003; Haefner, Citation2003), hemagglutination (Kakita et al., Citation1999), and more.
Antimycobacterial compounds have been isolated from marine organisms such as sponges, gorgonia, and soft corals (Donia & Hamann, Citation2003). König et al. (2000) screened 39 marine-derived natural products for antimycobacterial activity against Mycobacterium tuberculosis. and Mycobacterium avium.. The compounds represented various structural classes including terpenes, aliphatics, aromatics, alkaloids, and sterols. Fifteen compounds were derived from red algae, one from a brown alga, three from cyanobacteria, 10 from sponges, one from a soft coral, three from a gorgonian, three from lichens, and three were semisynthetic. Seven of the sponge-derived compounds were the most active against M. tuberculosis., with minimum inhibitory concentrations (MIC) values less than 10 μ g/mL. The compounds isolated from the red algae Laurencia obtusa. Hudson (Rhodomelaceae), Laurencia flexilis. Setchell (Rhodomelaceae), L. rigida. J. Agardh (Rhodophyceae), and Plocamium cartilagineum. Linnaeus (Plocamiaceae) had antimycobacterial activity ranging from MIC values of 16 to 64 μ g/mL. South Africa is home to a diverse seaweed population (Silva et al., 1996), and it is estimated that there are about 800 species (Stegenga et al., Citation1997). Red seaweeds constitute most of the 800 species, and more red than brown and green seaweeds are potentially harvestable and/or economically important (Critchley et al., Citation1998). Considering the amount of seaweeds available along the South African coast, there has been little investigation into their pharmaceutical potential. Vlachos et al. (Citation1997) screened 56 seaweeds for antimicrobial activity against 16 microorganisms. The brown alga, Zonaria subarticulata. Lamouroux (Dictyotaceae), showed the greatest inhibition against the microorganisms tested. Stirk et al. (Citation1996) tested crude algal extracts of 35 seaweed species for inhibitory activity to prostaglandin synthesis and found species from the red, brown, and green seaweeds with potential novel anti-inflammatory activity. Stirk and Van Staden (Citation1997) screened 20 algal species for cytokinin-like activity and found it present in most of the extracts.
In this investigation, Polysiphonia virgata. C. Agardh (Rhodomelaceae) was screened for antimycobacterial activity. This dark-red to blackish plant grows to 30 cm or more. It often exists within the tissues of other plants, but it is not necessarily parasitic; for example, it may be an epiphyte on the kelp (Ecklonia maxima. Osbeck; Alariaceae) (Stegenga et al., Citation1997). Polysiphonia virgata. C. Agardh is endemic to southern Africa and is found from the coast of Namibia to Brandfontein near Cape Agulhas. Other species of the genus Polysiphonia such as Polysiphonia furcellata. C. Agardh (Rhodomelaceae) (Caccamese et al., Citation1985), P. lanosa. Linnaeus (Rhodomelaceae), P. elongata. Hudson (Rhodomelaceae), P. nigra. Hudson (Rhodomelaceae), P. nigrescens. Hudson (Rhodomelaceae), P. urceolata. Lightfoot ex Dillwyn (Rhodomelaceae) (Hornsey & Hide, Citation1976), and P. virgata. C Agardh (Vlachos et al., Citation1997) contain antimicrobial compounds, and P. hendryi. Gardner (Rhodomelaceae) has antiviral compounds (Kim et al., Citation1997).
Materials and Methods
Sample collection
A sample of P. virgata. was collected at Sweet Water, Western Cape coast, on 4 July 2005. The alga was collected and verified by Mark Rothman and Chris Boothroyd from the Seaweed Research Unit of the Department of Environmental Affairs and Tourism: Marine and Coastal Management, which is based in the Botany Department at the University of Cape Town. A voucher specimen of the seaweed (BH 126936) was deposited at the Bolus Herbarium at the Botany Department of the University of Cape Town (UCT).
Extraction
The seaweed sample was placed in plastic bags and kept cool on ice. In the laboratory, the alga was washed with distilled water to remove surface salts, sand, and epiphytes. The seaweed was dabbed with paper towels to remove excess water, cut into small pieces, and placed in an oven at 50°C until completely dry (782.50 g). The dried material was milled using a blender and soaked in cold methanol for 2 h at 4°C. The methanol was filtered and the algal material steeped in 2 L dichloromethane (DCM) overnight at 4°C. The cold DCM was filtered and replaced with more DCM (5 × 2000 mL) at room temperature. The extracts were concentrated under vacuum using a Büchi Rotavapor R-205, (Switzerland) at 50°C.
Preparative fractionation using flash chromatography
Flash silica gel 60 chromatography of the resultant sample material (7.074 g) was done with hexane and increasing proportions of ethyl acetate (EtOAc) as eluant, resulting in 44 main fractions. Fraction 45 was obtained by washing the silica column with absolute ethanol until the eluant became colorless. The fractions were concentrated under vacuum using a Büchi Rotavapor R-205 at 50°C and dried completely under a stream of nitrogen.
Centrifugal preparative layer chromatography
Centrifugal preparative layer chromatography was performed using a Chromatotron model 8924 from Harrison Research Inc. (Palo Alto, CA, USA). The sample was fractionated on the Chromatotron using silica gel 60 PF-254 coated plates (2 or 1 mm thickness) and hexane-DCM-MeOH (7:2:1) as solvent. The polarity of the solvent system was gradually increased to remove the more polar fractions until only MeOH was effective. The less polar fractions were further fractionated using hexane-DCM-MeOH (5:3:2) or DCM-MeOH (5:5), and the polar fractions resolved well with either toluene or hexane:DCM:MeOH (7:2:1). In some cases, starting with hexane and introducing more polar solvents enabled the bands to elute from the silica plate with better separation. The fractions were observed under ultraviolet light at 254 nm and 366 nm.
Gas-liquid chromatography analysis of methyl esters
The sample was treated with methanol HCl, 3 M, to obtain the methyl esters of the lipids present, by refluxing 6 to 10 mg in 5 mL methanol HCl for 2 h. The reaction mixture was evaporated to dryness under vacuum. The fatty acid methyl esters were purified by silica gel column chromatography using a Pasteur pipette and eluting with hexane/ether (8:2 v/v). The solvent was evaporated to dryness and the methyl esters dissolved in hexane and analyzed by gas chromatography–mass spectrometry (GC-MS). The GC-MS analyses were recorded at 70 eV using a Carlo Erba GC 6000 Vega Series (Milan, Italy) equipped with a 40 m × 0.3 mm special performance capillary column (Lexus, PS089-OH) silanol-terminated (95%)-methyl-(5%)-phenylpolisiloxane copolymer stationary phase and He as the carrier gas. Analyses were performed under the following conditions: initial temperature, 130°C; rate of increase, 3°C /min; final temperature, 270o.C, held for 10 min.
Direct bioautography
Each fraction (500 μ g) was applied to an analytical thin-layer chromatography (TLC) 20 × 20 cm glass-backed plate with a fluorescent indicator and allowed to develop to the middle of the plate (about 9 cm) using hexane:chloroform:methanol (7:2:1) as the mobile phase. This permitted 18 fractions to be evaluated per plate. After development, the solvents were allowed to evaporate completely. Antimycobacterial activity was assessed using a Pasteur Institute strain of Mycobacterium aurum. and a clinical strain of Mycobacterium smegmatis.. These mycobacteria were cultured in Middlebrook-7H9 broth. The developed TLC plates were dabbed with the bacterial suspension using sterile cotton balls and incubated in a moist atmosphere at 37°C. Plates with M. aurum. were incubated for 3 days and M. smegmatis. for 2 days. At the end of the incubation period, the plates were sprayed with a p.-iodonitrotetrazolium violet aqueous solution (0.4 mg/mL), which was reduced to a pink color by the mitochondrial enzymes of live organisms. The sprayed plates were incubated for 1 h at 37°C, after which the clear zones were recorded as inhibition zones. The Rf.value was calculated as the inhibition zone distance from origin (midpoint) (mm)/solvent front distance from origin (mm). Purified fractions (500 μ g) were applied to the plates, but not allowed to develop in a solvent system. Inhibition zones were observed around the bioactive samples, and the zone sizes were measured. The results of the pure compounds were presented in minimum inhibitory quantity (MIQ), which is the minimal quantity of compound applied to the TLC plate that showed inhibition of the microorganisms as done by Moulari et al. (Citation2006). The MIQ values were determined by applying sample sizes ranging from 500 μ g to 0.4 μ g to the TLC plates. Ciprofloxacin (12 μ L of 1 mg/mL solution, i.e., 12 μ g) was used as a drug control.
Radiometric method
The radiometric method used was described by S.H. Siddiqi in the Product and Procedure Manual of the Bactec 460 TB System (1995). The main component of the method is the 7H12 Middlebrook TB medium that contains 14C-labeled substrate (palmitic acid) as a source carbon. As the bacterium grows, it consumes the palmitic acid, with release of 14CO2 into the atmosphere of the sealed vial. The Bactec-460 instrument (Johnston Laboratories, Towson, MD, USA) records the amount of 14CO2 and processes it as a growth index (GI) on the scale 0–999 (Lall & Meyer, Citation1999).
A rifampicin, isoniazid, and ethambutol sensitive M. tuberculosis. H37Rv strain was used in this investigation (supplied by the Microbiology Laboratory of the National Health Laboratory Services, based at Groote Schuur Hospital, Cape Town). The bacterium was grown on Löwenstein-Jensen slants or in Bacto Middlebrook 7H9 liquid medium. The same laboratory supplied a multidrug-resistant clinical isolate of M. tuberculosis. that was resistant to rifampicin, isoniazid, and ethambutol. The sensitive M. tuberculosis. H37Rv strain was inoculated into a Bactec vial containing 4 mL 7H12 Middlebrook TB medium. When the growth index of the bacterium vial reached values between 300 and 500, the broth culture was considered ready to be introduced into the test and control vials, each of which contained 4 mL 7H12 Middlebrook TB medium. The test vials contained 0.1 mL of the broth culture and fatty acids sample solution while the two positive control vials contained 0.1 mL of the broth culture and 0.1 μ g/mL isoniazid and 2.0 μ g/mL rifampicin, respectively. Two negative controls were used: one with 0.1 mL broth culture and the other was prepared by adding 0.1 mL of 1:100 dilution of the broth culture. The dilution was prepared in special diluting fluid. Test stock solutions were prepared in 12.5% ethanol and Bacto Middlebrook 7H9 broth. The test vials received 100 μ L of each fatty acid stock solution resulting in a 40-fold dilution and final concentrations (μ g/mL) of 50, 25, 12.5, 6.25, and 3.125. All the vials were incubated at 37°C and the growth index (GI) determined daily at the same time over a period of 4 to 12 days until the GI of the 1:100 control vial reached 30 or more. The sample vials with GI values below or equal to that of the 1:100 control vials on the final day of the assay were considered active against M. tuberculosis.. The percent inhibition was calculated as (König et al., Citation2000):
The MIC of each fatty acid solution was determined as the lowest concentration that yielded a growth index of less than that of the 1:100 control vial.
Results
The fractionation of the crude extract using flash chromatography produced 45 fractions of which 38 were active against M. aurum. and 27 against M. smegmatis. in the bioautography method, with Rf. values between 0.1 and 0.9. Samples with similar 1H NMR spectra, thin-layer chromatography profiles, and inhibition zones were combined and further purified using the Chromatotron, such as fractions 2 and 3 and fractions 17 to 30. Some of the purified fractions inhibited the growth of M. smegmatis. although the initial cruder fraction had shown no inhibition. The purified samples were screened for activity against M. smegmatis. only using direct bioautography. Further bioassay-guided fractionation using centrifugal preparative layer chromatography and analytical thin-layer chromatography resulted in the isolation of a mixture of fatty acids and hydrocarbons as had been anticipated from the 1H NMR spectra obtained (see ).
Figure 1 1H NMR spectrum of sample 6/9.
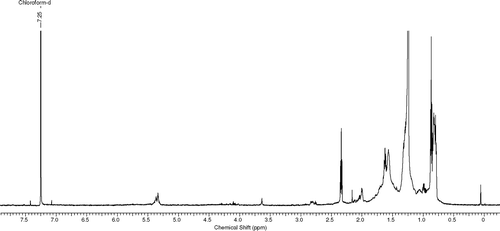
The resultant sample (69.9 mg) was tested against M. smegmatis. and the MIQ found to be 7.8 μ g (zone size = 4 mm), which compared well with ciprofloxacin, which showed a 12-mm-diameter zone at an amount of 12.5 μ g on the thin-layer chromatography plate. The sample was methylated and submitted for GC-MS analysis (16.4 mg). Hydrocarbons and methyl esters were identified, confirming the presence of fatty acids. All the methyl esters in the mixture were identified by comparing the retention times of GC peaks with those of the standard compounds under the same chromatographic conditions using commercially available standards (Sigma Aldrich Co., Germany).
Of the fatty acids identified in the GC-MS analysis, only dodecanoic acid, stearic acid, palmitic acid, oleic acid, linoleic acid, and myristic acid could be obtained commercially for further analysis. Pentadecanoic acid had to be omitted. Antimycobacterial activity of the fatty acids was assessed against M. smegmatis. using direct bioautography and M. tuberculosis. H37Rv and a clinical multidrug-resistant (MDR) strain of M. tuberculosis. in the Bactec-460 radiometric method. Dodecanoic acid, linoleic acid, and oleic acid inhibited the growth of M. smegmatis. with MIQ values of 3.125, 1.56, and 0.8 μ g, respectively (). Myristic acid, palmitic acid, and stearic acid showed no inhibition of M. smegmatis..
Table 1 Antimycobacterial activity of the bioactive fatty acids against M. smegmatis., M. tuberculosis,. and MDR M. tuberculosis..
In the Bactec-460 method, the fatty acid and hydrocarbon mixture showed moderate inhibition at 125 μ g/mL against M. tuberculosis. H37Rv. Lauric acid and linoleic acid showed 100% inhibition at 50 μ g/mL. Myristic acid showed 98% inhibition at 50 μ g/mL, and oleic acid showed 100% inhibition at 25 μ g/mL. Palmitic acid and stearic acid showed no significant inhibition. When tested against MDR M. tuberculosis., linoleic acid showed moderate inhibition at 50 μ g/mL, whereas lauric acid and myristic acid showed 75% and 88% inhibition, respectively, at the same concentration (). None of the other fatty acids inhibited the growth of MDR M. tuberculosis..
Discussion
Fatty acids are important components of terrestrial plant oils (Evans, Citation1996), and the same is true for marine plants. Many investigations into marine animal and plant lipids have been published (Carballeira, Citation2002; Dembitsky & Srebnik, Citation2002; Huang & Wang, Citation2004; Sanina et al., Citation2004). The fatty acids identified in this investigation of P. virgata. are common to red algae and specifically to the genus Polysiphonia.. Johns et al. (Citation1979) investigated the fatty acid composition of 10 seaweeds from Australian waters. Polysiphonia pungens. Hollenberg (Rhodomelaceae) was one of the species investigated and showed high levels of polyunsaturated fatty acids, such as arachidonic acid. Palmitic acid was the main saturated fatty acid. cis.-Vaccenic acid, cis.-9-hexadecenoic or palmitoleic acid, and myristic acid were minor components in the fatty acid mixture. Pettitt et al. (Citation1989) investigated the lipids of the algae Chondrus crispus. Stackhouse (Gigartinaceae) and Polysiphonia lanosa. Linnaeus (Rhodomelaceae). The major fatty acids detected were palmitic, oleic, arachidonic, and eicosapentaenoic acids. A red alga with a lipophilic portion high in antioxidant activity, Rhodomela confervoides. Hudson (Rhodomelaceae), had only 37.7% mg/g dry weight in lipid content but the most unsaturated fatty acids (46% of lipophylic content), such as 11-hexadecenoic acid and 5,8,11,14,17-eicosapentaenoic acid. Halogenated fatty acids are very common in the marine environment, and many have been isolated from algae and other marine organisms as shown in a review done by Dembitsky and Srebnik (Citation2002).
Many publications are available about fatty acids with antibiotic activities. Findlay and Patil (Citation1984) isolated the fatty acids (6Z.,9Z.,12Z.,15Z.)-hexadecatetraenoic acid, (6Z.,9Z.,12Z.,15Z.)-octadecatetraenoic acid, and (6Z.,9Z.,12Z.)-hexdecatrienoic acid as the antibacterial compounds from the diatom (Navicula delognei f. elliptica. Lobban). It inhibited the growth of Staphylococcus aureus., Staphylococcus epidermidis,. and Proteus vulgaris.. Benyagoub et al. (Citation1996) isolated 9-heptadecenoic acid and 6-methyl-9-heptadecenoic acid as the compounds responsible for the antibiotic activity of the fungus Sporothrix flocculosa.. Ohta et al. (Citation1993) investigated the antibiotic activities of 10 fatty acids and their methyl esters and found γ-linolenic acid (C18:3) to have the highest antibiotic activity. α-Linolenic acid, along with eicosapentaenoic acid, and docosahexaenoic acid also had strong antibiotic activity.
The antimycobacterial activities of long-chain fatty acids have been demonstrated by a few investigations. A potent antimycobacterial compound, 3-nitropropionic acid, was isolated from endophytic fungi found on Thai medicinal plants with a minimum inhibition concentration of 3.3 μ M (Chomcheon et al., Citation2005). Antimycobacterial (MIC value of 25 μ g/mL) and antiplasmodial activity (IC50 value of 7.2 μ g/mL) was reported for scleropyric acid isolated from the twigs of Scleropyrum wallichianum. Wight & Arn. (Santalaceae) (Suksamrarn et al., Citation2005). Stavri et al. (Citation2004) investigated the hexane extract of hops, namely Humulus lupulus. L. (Cannabaceae), for antimycobacterial compounds, and found a fatty acid mixture to be the bioactive principle. Among the fatty acids were palmitic, stearic, and oleic acid with small quantities of lignoceric, arachidic, behenic, and linoleic acids. All saturated fatty acids were inactive against M. fortuitum., whereas the unsaturated fatty acids, namely oleic and linoleic acids, showed MICs between 4 and 16 μ g/mL. Kondo and Kanai (Citation1972) looked at the lethal effect of long-chain fatty acids on mycobacteria and found that of 11 long-chain fatty acids, oleic, linoleic, and myristic acids showed the greatest activity against M. tuberculosis. and M. bovis.. Seidel and Taylor (Citation2004) investigated the dried roots of Pelargonium reniforme. Curtis (Geraniaceae) and P. sidoides. D.C. (Geraniaceae) for antibacterial activity against M. aurum. and M. smegmatis,. and showed the active components to contain palmitic, oleic, and linoleic acids as the major bioactive components. Although the initial mixture of fatty acids and hydrocarbons was highly active against M. smegmatis. in direct bioautograph but weakly active against M. tuberculosis. in the radiometric method, the antimycobacterial activity of the individual fatty acids such as oleic acid, and linoleic acid are consistent with the above investigations.
Acknowledgments
The authors thank the National Research Foundation for financial assistance, Dr. Néstor M. Carballeira from the Department of Chemistry at the University of Puerto Rico for his invaluable guidance, Ms. Jean Mckenzie for the NMR spectra, and Dr. Stephan Louw, from the Central Analytical Facility, University of Stellenbosch, for the LC-MS and GC-MS analysis.
References
- Ahn C, Jeon Y, Kang D, Shin T, Jung B. Free radical scavenging activity of enzymatic extracts from a brown seaweed Scytosiphon lomentaria. by electron spin resonance spectrometry. Food Res Int 2004; 37: 253–258
- Ahn M, Yoon K, Kim C Y, Min S, Kim Y, Kim H J, Kim J H, Shin C, Lee C, Kim T G, Kim S H, Huh H, Kim J. Inhibition of HIV-1 reverse transcriptase and HIV-1 integrase and antiviral activity of Korean seaweed extracts. J Appl Phycol 2002; 14: 325–329
- Benyagoub M, Rhlid R B, Bélanger R R. Purification and characterization of new fatty acids with antibiotic activity produced by Sporothrix flocculosa.. J Chem Ecol 1996; 22: 405–413
- Boyd M R, Gustafson K R, McMahon J B, Shoemaker R H, O'Keefe B R, Mori T, Gulakowski R J, Wu L, Rivera M I, Laurencot C M, Currens M J, Cardellina J H, II, Buckheit R W, Nara P L, Pannell L K, Sowder R C, Henderson L E. Discovery of cyanovirin-N, a novel human immunodeficiency virus-inactivating protein that binds viral surface envelope glycoprotein gp 120: Potential application to microbicide development. Antimicrob Agents Chemother 1997; 41: 1521–1530
- Caccamese S, Toscano R M, Furnari G, Cormaci M. Antimicrobial activities of red and brown algae from southern Italy coast. Bot Mar 1985; 28: 505–507
- Carballeira N M. New advances in the chemistry of methoxylated lipids. Prog Lipid Res 2002; 41: 437–456
- Carlucci M J, Ciancia M, Matulewicz M C, Cerezo A S, Damonte E B. Antiherpetic activity and mode of action of natural carrageenans of diverse structural types. Antiviral Res 1999; 43: 93–102
- Chevolot L, Foucault A, Chaubet F, Kervarec N, Sinquin C, Fisher A, Boisson-Vidal C. Further data on the structure of brown seaweed fucans: Relationships with anticoagulant activity. Carbohydr Res 1999; 319: 154–165
- Chomcheon P, Wiyakrutta S, Sriubolmas N, Ngamrojanavanich N, Isarangkul D, Kittakoop P. 3-Nitropropionic acid (3-NPA), a potent antimycobacterial agent from endophytic fungi: Is 3-NPA in some plants produced by endophytes?. J Nat Prod 2005; 68: 1103–1105
- Critchley A T, Gillespie R D, Rotmann K WG. Seaweed resources of South Africa. Seaweed Resources of the World, A T Critchley, M Ohno. Japan International Cooperation Agency, YokosukaJapan 1998; 413–425
- Dembitsky V M, Srebnik M. Natural halogenated fatty acids: Their analogues and derivatives. Prog Lipid Res 2002; 41: 315–367
- Donia M, Hamann M T. Marine natural products and their potential applications as anti-infective agents. Lancet Infect Dis 2003; 3: 338–348
- Evans W C. Hydrocarbons and derivatives. Trease and Evans' Pharmacognosy, 14th edn. WB Saunders, London 1996; 172
- Findlay J A, Patil A D. Antibacterial constituents of the diatom Navicula delognei.. J Nat Prod 1984; 47: 815–818
- Fujiwara P I, Simone P M, Munsif S S. Treatment of tuberculosis. Tuberculosis, a Comprehensive International Approach, 2nd ed. revised and expanded, L B Reichman, E S Hershfield. Marcel Dekker Inc., New York 2000; 401
- Haefner B. Drugs from the deep: Marine natural products as drug candidates. Drug Discovery Today 2003; 8: 536–544
- Hiroishi S, Sugie K, Yoshida T, Morimoto J, Taniguchi Y, Imai S, Kurebayashi J. Antitumour effects of Marginisporum crassissimum. (Rhodophyceae), a marine red alga. Cancer Lett 2001; 167: 145–150
- Hornsey I S, Hide D. The production of antimicrobial compounds by British marine algae. II. Seasonal variation in production of antibiotics. British Phycol J 1976; 11: 63–67
- Huang K, Wang B. Antioxidant capacity and lipophilic content of seaweeds collected from the Qingdao coastline. J Agric Food Chem 2004; 52: 4993–4997
- Huheihel M, Ishanu V, Tal J, Arad S. Activity of Porphyridium. sp. polysaccharide against Herpes simplex viruses in vitro in vivo.. J Biochem Biophys Methods 2002; 50: 189–200
- Jiang Z, Jensen P R, Fenical W. Lobophorins A and B, new antiinflammatory macrolides produced by a tropical marine bacterium. Bioorg Med Chem Lett 1999; 9: 2003–2006
- Johns R B, Nichols P D, Perry G J. Fatty acid composition of ten marine algae from Australian waters. Phytochemistry 1979; 18: 799–802
- Kakita H, Fukuoka S, Obika H, Kamishima H. Isolation and characterisation of a fourth hemagglutinin from the red alga, Gracilaria verrucosa., from Japan. J Appl Phycol 1999; 11: 49–56
- Kaufmann S HE. Is the development of a new tuberculosis vaccine possible?. Nat Med 2000; 6: 955–960
- Kim J H, Hudson J B, Huang A M, Bannister K, Jin H, Choi T J, Towers G HN, Hong Y K, De Wreede R E. Biological activities of seaweed extracts from British Columbia, Canada, and Korea. I. Antiviral activity. Can J Bot 1997; 75: 1656–1660
- Kondo E, Kanai K. Lethal effect of long-chain fatty acids on mycobacteria. Jpn J Med Sci Biol 1972; 25: 1–13
- König G M, Wright A D, Franzblau S G. Assessment of antimycobacterial activity of a series of mainly marine derived natural products. Planta Med 2000; 66: 337–342
- Koyanagi S, Tanigawa N, Nakagawa H, Soeda S, Shimeno H. Oversulfation of fucoidan enhances its anti-angiogenic and antitumor activities. Biochem Pharmacol 2003; 65: 173–179
- Lall N, Meyer J JM. In vitro. inhibition of drug-resistant and drug-sensitive strains of Mycobacterium tuberculosis. by ethnobotanically selected South African plants. J Ethnopharmacol 1999; 66: 347–354
- Moulari B, Pellequer Y, Lboutounne H, Girard C, Chaumont J P, Millet J, Muyard F. Isolation and in vitro. antibacterial activity of astilbin, the bioactive flavanone from the leaves of Harungana madagascariensis. Lam. ex Poir. (Hypericaceae). J Ethnopharmacol 2006; 106: 272–278
- O'Brien R J. Tuberculosis: Scientific Blueprint for Tuberculosis Drug Development. Global Alliance for TB Drug Development, New York, Churchill Livingstone 2001; 3
- Ohta S, Chang T, Kawashima A, Aozasa O, Mase Y, Nagate T, Kitamura K, Kondo M, Miyata H. Antibiotic activity of unsaturated fatty acids on methicillin-resistant Staphylococcus aureus.. Biosci Biotechnol Biochem 1993; 57: 2194–2195
- Okai Y, Higashi-Okai K. Potent anti-inflammatory activity of pheophytin A derived from edible green alga, Enteromorpha prolifera. (Sujiao-nori). Int J Immunopharmacol 1997; 19: 355–358
- Pettitt T R, Jones A L, Harwood J L. Lipids of the marine red algae, Chondrus crispus Polysiphonia lanosa.. Phytochemistry 1989; 28: 399–405
- Raviglione M C. The TB epidemic from 1992 to 2002. Tuberculosis 2002; 83: 4–14
- Sanina N M, Goncharova S N, Kostetsky E Y. Fatty acid composition of individual polar lipid classes from marine macrophytes. Phytochemistry 2004; 65: 721–730
- Seidel V, Taylor P W. In vitro. activity of extracts and constituents of Pelagonium. against rapidly growing mycobacteria. Int J Antimicrob Agents 2004; 23: 613–619
- Shan B E, Yoshida Y, Kuroda U, Yamashita U. Immunomodulating actvity of seaweed extract on human lymphocytes in vitro.. Int J Immunopharmacol 1999; 21: 59–70
- Siddiqi S H. Bactec 460 TB System. Product and Procedure Manual, revision D. Becton Dickinson Diagnostic System, Sparks, MD 1995
- Stavri M, Schneider R, O'Donnell G, Lechner D, Bucar F, Gibbons S. The antimycobacterial components of hops (Humulus lupulus.) and their dereplication. Phytother Res 2004; 18: 774–776
- Stegenga H, Bolton J J, Anderson R J. Seaweeds of the South African West Coast. 1997, Cape Town, Contributions from the Bolus Herbarium, Number 18, pp. 8, 554
- Stirk W A, Jäger A K, Van Staden J. Screening of some South African seaweeds for prostaglandin-synthesis inhibitors. South African J Bot 1996; 62: 108–110
- Stirk W A, Van Staden J. Screening of some South African seaweeds for cytokinin-like activity. South African J Bot 1997; 63: 161–164
- Suksamrarn A, Buaprom M, Udtip S, Nuntawong N, Haritakun R, Kanokmedhakul S. Antimycobacterial and antiplasmodial unsaturated carboxylic acid from the twigs of Scleropyrum wallichianum.. Chem Pharm Bull 2005; 53: 1327–1329
- Vlachos V, Critchley A T, Von Holy A. Antimicrobial activity of extracts from selected southern African marine macroalgae. South African J Sci 1997; 93: 328–332
- World Health Report. Global tuberculosis control: surveillance planning, financing. 2004, http://www.who.int/tb/publications/global_report/en/ Available at URL
- Zhang Q, Li N, Liu X, Zhao Z, Li Z, Xu Z. The structure of a sulfated galactan from Porphyra haitanensis. and its in vivo. antioxidant activity. Carbohydr Res 2004; 339: 105–111
- Zhou G, Sun Y P, Xin H, Zhang Y, Li Z, Xu Z. In vivo. antitumor and immunomodulating activities of different molecular weight lambda-carrageenans from Chondrus ocellatus.. Pharmacol Res 2004; 50: 47–53