
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
Farmgate balances are used as a tool for monitoring nutrient surpluses at farm level. In Germany, preparation of farmgate balances is legally mandatory and also requires data on chemical body composition, especially concentration of nitrogen (N) and phosphorus (P), of farm animals. It is well known that increased N and P efficiency results in lowered N and P excretions with the manure and therefore mitigates negative consequences of high N and P release into the environment (e.g. eutrophication of surface waters), especially in areas with high livestock density. In this context, feeding N- and P-reduced diets can be a strategy for increasing N and P efficiency in fattening pigs. To investigate the influence of N- and P-reduced diets on chemical body composition of barrows and to update current used data basis, 8 barrows were slaughtered after a balance trial and their bodies were subjected to full body analysis. During the balance trial, pigs received the control diet (CON) meeting common nutrient requirements or the N- and P-reduced diet (NPred) in a three-phased feeding regimen (n = 4/diet, 3 weeks/phase). Pigs were slaughtered with an average live weight (LW) of 123.3 ± 7.5 kg and carcasses were manually dissected in four fractions. Fractions were analysed for nutrient concentration. Furthermore, organs were weighed individually and blood serum was sampled during exsanguination. Serum samples were analysed for clinical-chemical traits. Chemical body composition did not significantly differ between NPred- and CON-fed pigs. N concentration was 23.3 ± 0.3 and 24.5 ± 1.0 g/kg, P concentration was 5.2 ± 0.1 and 5.5 ± 0.4 g/kg in the empty body of NPred- and CON-fed pigs (p = 0.073, 0.164). N and P retention between the experiment’s start and slaughter did not differ between the feeding groups (p = 0.641, 0.240). Variables related to liver integrity, energy metabolism and electrolytes were similar between CON- and NPred-fed pigs. Traits related to protein metabolism showed significantly reduced concentrations of urea and albumin in NPred-fed pigs (p = 0.013, 0.025), but no hypoalbuminaemia. Results suggest that N- and P-reduced feeding does not significantly affect chemical body composition of contemporary barrows.
1. Introduction
Farmgate balances (= difference between nutrient input and output of a farm) are used as a monitoring tool for identifying nutrient surpluses and deficits at farm level (Van Beek et al. Citation2003). In Germany, the preparation of farmgate balances is mandatory by law in order to adequately implement the European Nitrates Directive (Council Directive 91/676/EEC Citation1991; BMEL Citation2017). Nutrient output of pig farms is calculated from the difference between nutrient intake (feed intake · dietary nutrient concentration) and nutrient retention (live weight gain · nutrient concentration in animal product) (BMEL Citation2017). Therefore, chemical body composition of pigs (animal products), in particular N and P concentration, represents the basis for farmgate balances. It is well known that chemical body composition of pigs can be affected by sex, age and genotype (Gebhardt et al. Citation1981; GfE Citation2006; Otten et al. Citation2013; Zomeño et al. Citation2023). Furthermore, nutrition has a decisive influence on chemical body composition. Undersupply with amino acids (AA), especially the first-limiting AA lysine, can lead to an increase in intramuscular fat content (Liao et al. Citation2015). For protein retention, not only the dietary supply with AA but also with energy is crucial (Pettigrew and Esnaola Citation2001). However, the risk of body adiposity increases with increasing energy intake (Pettigrew and Esnaola Citation2001; Kirchgeßner et al. Citation2014). Insufficient supply with P can lead to a decreased P content in the skeletal system of pigs (Lüdke et al. Citation1990). Since genotype determines protein and fat retention (Kirchgeßner et al. Citation2014), breeding progress consequently leads to changes in chemical body composition. As one result, an increase in the proportion of lean meat content has been achieved in the last few decades (Ruiz-Ascacibar et al. Citation2017). In addition, Weniger and Funk (Citation1953) showed that the proportion of bones in the body varies considerably between different pig breeds. Genotype therefore also has an effect on P and calcium (Ca) retention, as the majority of P and Ca in the body is stored in the bones (Pettey et al. Citation2015; Gonzalo et al. Citation2018). Hence, Kirchgeßner et al. (Citation2014) demand that data on chemical body composition have to be updated regularly, as these data are not only the basis for the preparation of farmgate balances but also for the determination of retention values. Currently used data on chemical body composition of pigs is based on experimental results from 1969 to 2003 (GfE Citation2006) and therefore cannot reflect current developments in genetics or feeding.
Meeting government environment policies, N- and P-reduced diets are increasingly applied in practice. The aim of N- and P-reduced feeding is to supply pigs close to their N and P requirements and to minimise N and P concentrations in the manure (Dourmad and Jondreville Citation2007). Manure, used as an organic fertiliser in agriculture, contributes to a major extent to high N and P emissions with negative environmental consequences (e.g. eutrophication of surface waters, contamination of groundwater, soil acidification), especially in regions with high livestock density (Oliveira and Machado Citation2013; Van den Broeke et al. Citation2022). In addition, the use of inorganic P in pig feed needs to be reduced as it represents a non-renewable source (Gonzalo et al. Citation2018). Many studies confirmed that N- and P-reduced diets are an effective strategy to increase N and P efficiency in contemporary fattening pigs without impairing growth performance (Schulin-Zeuthen et al. Citation2007; Ruiz-Ascacibar et al. Citation2017; Beckmüller et al. Citation2023). Therefore, N- and P-reduced feeding can contribute to a more environmentally sustainable pig production.
With regard to the increasing utilisation of N- and P-reduced diets in agricultural practice, the question arises whether N- and P-reduced feeding has an influence on chemical body composition and N retention of contemporary fattening pigs. In order to answer this question, 8 barrows which had previously been used for a balance trial on N- and P-reduced feeding (Beckmüller et al. Citation2023) were slaughtered and the carcasses subjected to full body analysis. Additional aim of the present experiment was to update data on chemical body composition and to compare N and P retention based on balance method and full body analysis. Furthermore, conclusions should be drawn if N- and P-reduced feeding has an effect on health traits and the physiological development of pigs during fattening period.
2. Material & methods
All procedures in the present study were conducted at the Institute of Animal Nutrition, Friedrich-Loeffler-Institut (FLI) in Braunschweig, Germany in accordance with the European Community regulations concerning the protection of experimental animals and the guidelines of the German Animal Welfare Act and was approved by the Lower Saxony State Office for Consumer Protection and Food Safety (LAVES), Oldenburg, Germany (file number 33.19 -42502-04-20/3351).
2.1. Experimental setup and diets
Animals, experimental diets and conduct were described previously (Beckmüller et al. Citation2023). Briefly, eight barrows (BHZP db. Viktoria x Piétrain) were surgically fitted with a simple T-cannula at the terminal ileum and, after a convalescence period, allotted to one of two experimental diets (live weight [LW] = 61.5 ± 2.1 kg): The control diet (CON) meeting nutrient requirements for fattening pigs according to the Society of Nutrition Physiology (GfE Citation2006) and the diet reduced in nitrogen and phosphorus concentration (NPred) adapted from recommendations of the German Agricultural Society (DLG Citation2019). Both diets were wheat-barley based and applied in a three-phased feeding regimen (3 weeks/phase). Analysed crude protein (CP) concentration amounted to 168.2/143.8, 156.8/126.6 and 137.8/114.0 g/kg diet (as-fed basis) and phosphorus concentration to 5.5/4.6, 5.5/4.9 and 4.3/3.8 g/kg diet (as-fed basis) for CON/NPred in phase I, II and III, respectively. Pigs were individually housed and had ad libitum access to feed and water. In the last week of each phase, pigs were moved to flatdeck units for the separate collection of faeces, urine (quantitatively) and ileal digesta (qualitatively). After the last collection phase (= end of phase III), pigs were slaughtered with an average LW of 123.3 ± 7.5 kg in the experimental abattoir of the Institute of Animal Nutrition, Friedrich-Loeffler-Institute (FLI), Braunschweig for full body analysis. Pigs were not fasted before slaughter.
2.2. Slaughter procedures and full body fractionation
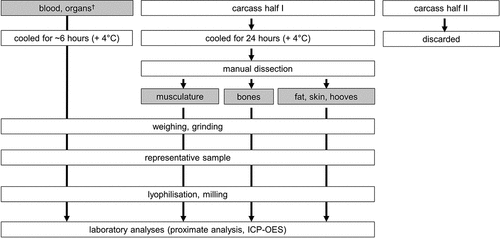
Procedures of the full body fractionation are shown in . Pigs were electrically stunned and exsanguinated. Blood was collected quantitatively for each pig during this procedure. Pigs were scalded and debristled. Hooves were collected for each pig, but the separate collection of bristles was not possible. Viscera (thoracic and abdominal) were dissected and content of gastrointestinal tract, urinary and gall bladder removed manually. Organs were weighed individually and then added to the blood fraction. The carcasses were divided into two halves and it was assumed that both halves represented comparable nutrient content. Therefore, only the right carcass half was used for further separation according to Otten et al. (Citation2013) and Ruiz-Ascacibar et al. (Citation2017). Thus, 4 distinct fractions were produced manually: 1. blood and all organs (incl. spinal cord, brain, eyes, penis), 2. skeletal muscles, 3. bones and 4. skin, fat and hooves. All fractions except fraction “blood, organs” were produced from one carcass half per pig. Since blood and organs were collected immediately after slaughter, fraction “blood, organs” was processed on the same day (immediately stored at + 4°C). Overnight, the carcass halves were stored refrigerated (+4°C). Time-consuming manual separation of fraction “skeletal muscles“, “bones“and “skin, fat, hooves“was carried out on the following day. Each fraction was weighed individually immediately after their production. All fractions except bones were ground in a commercial meat cutter. Bones were frozen at −20°C and milled in a frosted state on a bone mill. A representative sample of each fraction was taken, stored at −20°C until freeze-drying. Freeze-dried samples of the fractions “blood, organs”, “skeletal muscles” and “bones” were milled to pass a 1 mm sieve (ZM 100, RETSCH GmbH, Haan, Germany). The “skin, fat, hooves” fraction could not be ground on a mill due to its high fat concentration. Therefore, freeze-dried material was manually dispersed. Fraction samples were then analysed for their nutrient concentration. During blood collection, a blood sample was taken using serum tubes (with clotting activator, Sarstedt, Nümbrecht, Germany). The amount of blood in the serum tube was weighed and weight was re-added to the weight of fraction “blood, organs” during evaluation.
Figure 1. Procedures of the body fractionation of 8 barrows (average live weight 123.3 ± 7.5 kg). The four fractions are highlighted in grey.
2.3. Analyses
Collected samples of fractions were analysed according to the methods of the Association of German Agricultural Analytic and Research Institutes (VDLUFA Citation2012).
No defattening procedures were used prior to analyses. Nitrogen was measured using the method of Kjeldahl (method number 4.1.1) and multiplied by a factor of 6.25 to get the crude protein concentration. Dry matter, ether extract and crude ash were determined according to methods 3.1, 5.1.1 and 8.1, respectively. Phosphorus (P), calcium (Ca), magnesium (Mg), sodium (Na), zinc (Zn), copper (Cu) manganese (Mn) and iron (Fe) were analysed by optical emission spectrometry with inductively coupled plasma (ICP-OES Quantima; GBC Scientific Equipment Pty. 177 Ltd., Melbourne, Vic, Australia) according to method 10.8.2.
Serum samples were photometrically analysed (IndikoTM Plus, Thermo Scientific GmbH, Dreieich, Germany) for activity of enzymes (alkaline phosphatase (ALP), alanine aminotransferase (ALT), aspartate aminotransferase (AST), gamma-glutamyl-transferase (GGT), glutamate dehydrogenase (GLDH)) and concentration of bilirubin (total, direct). Furthermore, serum samples were analysed for traits of protein metabolism (total protein, albumin, creatinine, urea), energy metabolism (cholesterol, triglycerides, glucose) and electrolytes (calcium, chloride, phosphorus, potassium and sodium). One blood sample of a CON-fed pig was removed from the evaluation because it was haemolytic. Urine samples were photometrically analysed (IndikoTM Plus, Thermo Scientific GmbH, Dreieich, Germany) for concentration of urea and creatinine.
2.4. Calculations
As the content of gastrointestinal tract, urinary and gall bladder is affected by individual variations, empty body weight (EBW) was calculated. EBW was defined as live weight (LW) without the content of gastrointestinal tract, urinary and gall bladder according to Haude (Citation2003) and Otten et al. (Citation2013). It was calculated as the sum of the 4 fractions of full body analysis (all values in kg, fresh matter):
Weight of the fractions “skeletal muscles”, “bones” and “skin, fat, hooves” was multiplied by 2, because only one carcass half was dissected.
Carcass weight was calculated according to (Regulation (EU) No 1308/2013 Citation2013) (all values in kg, fresh matter):
Dressing percentage [kg] was calculated using carcass weight [kg] and LW [kg]:
Using the weight of the fractions [kg, fresh matter], their DM concentration [% of fresh matter] and nutrient concentration [g/kg dry matter], nutrient content per fraction was calculated (in g per fraction):
Weight of the fractions “skeletal muscles”, “bones” and “skin, fat, hooves” was previously multiplied by a factor of 2, since only one carcass half was dissected.
Total nutrient content in empty body was determined as the sum of nutrient content of all four fractions (all values in g):
For calculation of nutrient concentration in the empty body [g/kg EBW], total nutrient content in empty body was divided through EBW.
Body gross energy (GE) content was calculated using empty body content of crude protein and ether extract [both in g] according to (GfE Citation2006):
N and P retention was calculated by deducting the initial (start of the trial, LW 61.5 ± 2.1 kg) from the corresponding final (end of the trial, LW 123.3 ± 7.5) body N and P content of the barrows according to Pallauf and Walz (Citation1988), Haude (Citation2003) and Otten et al. (Citation2013). For the calculation of daily N and P retention [g/d], N and P retention was divided through the days of the experiment (3 phases · 21 days = 63 days) and furthermore related to metabolic LW0.67. Due to the surgical implantation of the simple T-cannula at the terminal ileum prior to the trial, it was not possible to analyse corresponding initial body N and P content at the start of the trial. Therefore, body crude protein (CP) content at the start of the trial in [kg/pig] was calculated using Equation 8 of (GfE Citation2006):
It was assumed that EBW is 94% of LW (GfE Citation2006). Body CP content was divided by 6.25 for body N content. Initially, regression equation (Equation 8, GfE Citation2006) was evaluated. For this purpose, data on analysed body CP content of pigs from the present experiment and from experiments published by Haude (Citation2003) was used. In contrast, no regression equation was available for calculating body P content (GfE Citation2006). Therefore, we used data published by Ruiz-Ascacibar et al. (Citation2019) on P concentration (g/kg EBW): they also conducted a full body analysis and reported an average P concentration of 5.73 g/kg EBW in barrows with a LW of 60 kg, which is comparable to the LW of the barrows at the start of the present trial. Again, it was assumed that EBW represents 94% of LW (GfE Citation2006) and this was applied to calculate initial body P content.
2.5. Statistics
Statistical analyses were performed using the PROC TTEST procedure of SAS (version 9.4; SAS Institute Inc., Cary, NC). P-values < 0.05 were deemed significant and p-values < 0.1 were regarded as a trend. The relationship between calculated (using regression Equation 8, GfE Citation2006) and analysed body CP concentration (this study) was evaluated by linear regression (PROC REG). Results are presented as means ± standard deviation. Data on urinary creatinine and urea excretion were statistically analysed using the PROC MIXED procedure of SAS with a restricted maximum likelihood model (REML). Phase, diet and their interaction were defined as fixed effects and phase was used as repeated measurement. Co-variance structure was chosen according to the corrected Akaike’s information criterion (AICC). An adjusted Tukey–Kramer test was applied as post-hoc procedure.
3. Results
3.1. Health status of finisher pigs
All pigs remained healthy during the entire experiment and there was no difference in voluntary feed intake and live weight between both dietary groups (Beckmüller et al. Citation2023). Relative organ weights did not differ significantly between CON- and NPred-fed pigs (). Relative liver weight appeared to be slightly higher in NPred-fed pigs but only numerically as a statistical trend. NPred-fed pigs showed a significant lower concentration of albumin and urea in blood (). Concentration of variables of energy metabolism, liver integrity and electrolytes did not differ between CON- and NPred-fed pigs (). Urinary concentration of urea was significantly lowered in NPred-fed pigs in all fattening phases, but no difference in urinary concentration of creatinine was observed between both dietary groups (supplementary material S1).
Table 1. Relative organ weights [g/kg live weight] of barrows at slaughter fed the control diet (CON, n = 4) or the diet with reduced nitrogen and phosphorus concentration (NPred, n = 4). Data are presented as means ± standard deviation.
Table 2. Clinical-chemical traits in blood serum of barrows at slaughter fed the control diet (CON, n = 3) and the diet with reduced nitrogen and phosphorus concentrations (NPred, n = 4). Data are presented as means ± standard deviation.
3.2. Full body analysis
Mean LW at slaughter was 123.3 ± 10.6 and 123.3 ± 4.5 kg for CON- and NPred-fed pigs, respectively (p = 0.997) (Beckmüller et al. Citation2023). Empty body weight (EBW) was similar for CON- and NPred-fed pigs, as well as the proportion of EBW in LW (). Furthermore, carcass weight and dressing percentage did not differ between CON- and NPred-fed pigs (). Weights of the fractions relative to empty body weight were similar for CON- and NPred-fed pigs (). Chemical body composition did not show any significant differences between CON- and NPred-fed pigs (). But concentration of crude protein (CP)/nitrogen (N) and calcium (Ca) was lowered in NPred-fed pigs, which was observed as a statistical trend (CP/N: p = 0.073, Ca: p = 0.077). Sum of concentrations for CP, ether extract, crude ash and water (water concentration = 1000 – dry matter concentration) was slightly below 100% of EBW caused by scope of analysis and amounted to 99.1 ± 0.6% and 98.4 ± 0.1% of EBW for CON and NPred, respectively.
Table 3. Empty body weight (EBW), carcass weight, dressing percentage and fractions of full body analysis of barrows at slaughter fed the control diet (CON, n = 4) and the diet with reduced nitrogen and phosphorus concentrations (NPred, n = 4). Data are presented as means ± standard deviation.
Table 4. Chemical body composition of barrows at slaughter fed the control diet (CON, n = 4) and the diet with reduced nitrogen and phosphorus concentrations (NPred, n = 4). Data are presented as means ± standard deviation.
Concentrations of magnesium, sodium, manganese, zinc, iron and copper in the empty body did not differ between CON- and NPred-fed pigs (supplementary material S2).
Concentration of dry matter, CP/N, ether extract, crude ash, phosphorus (P) and Ca in each fraction of full body analysis are presented in the supplementary material S3. Dry matter concentration in the fraction “blood, organs” was higher in NPred- than CON-fed pigs (p = 0.003). Concentration of CP/N was lower in the fractions “blood, organs” and “bones” of NPred-fed pigs (p = 0.030, 0.041). A significantly lower concentration of ether extract and Ca in the bones of NPred-fed pigs was observed (p = 0.040, 0.027). In addition, P concentration in the bones was lower in NPred- than CON-fed pigs, but this was only observed as a statistical trend (p = 0.060).
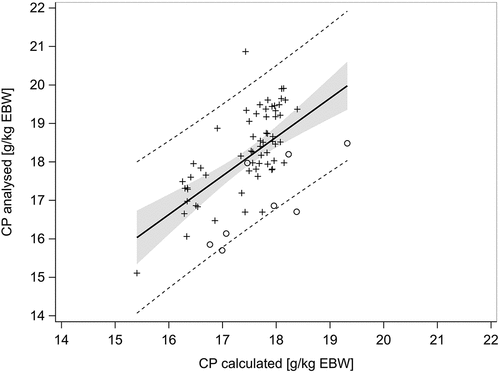
Linear regression analysis showed a significant relation between calculated (using Equation 8 of GfE (Citation2006)) and analysed body CP content (p < 0.001) (). The regression model was y = 0.53 + 1.01·x (p = 0.849, < 0.001) with RMSE = 0.92 kg and adjusted r2 = 0.36. Although the variance explained by the linear regression amounted to only 36% the slope of 1.01 suggested a good agreement between the CP content of EBW as predicted by Equation 8 (GfE Citation2006) and the analysed one retrieved from the present and other studies. Based on this agreement, it seemed to be justified to estimate the initial body CP content by using Equation 8 of GfE (Citation2006). For further use the so derived initial CP content was divided by a factor of 6.25 to obtain the body N content.
Figure 2. Regression between calculated and analysed body crude protein (CP) concentration of barrows from the present experiment (circle), a performance trial (plus) and a balance trial (cross) of Haude (Citation2003). For calculation, Equation 8 of GfE (Citation2006) was used. The black line shows the regression line, symbols show the observations (n = 71), grey shaded area shows the 95% confidence interval, the dashed lines the 95% prediction limits. The regression model is y = 0.53 + 1.01·x with RMSE = 0.92 kg and adjusted r2 = 0.36.
Calculated initial body N concentration (= at the beginning of the trial) did not differ between the feeding groups (). Values for initial body P concentration were obtained from Ruiz-Ascacibar et al. (Citation2019). Retention of N and P did not show any differences between CON- and NPred-fed pigs, neither absolutely nor in relation to time and/or metabolic LW0.67 ().
Table 5. Initial body crude protein (CP) concentration and calculated nitrogen (N) and phosphorus (P) retention of barrows fed the control diet (CON, n = 4) and the diet with reduced nitrogen and phosphorus concentrations (NPred, n = 4). Results are presented as means ± standard deviation.
4. Discussion
To examine whether N- and P-reduced feeding has an effect on health traits, chemical body composition and N and P retention of contemporary barrows, 8 barrows were slaughtered after a balance trial and their carcasses were analysed for chemical body composition.
Studies in the past confirmed that N- and P-reduced feeding can significantly reduce N and P excretion in relation to intake without impairing growth performance of fattening pigs. However, health status of the pigs should not be neglected. Evaluation of clinical-chemical traits confirmed the healthy condition of all pigs. Reduction of nitrogen (N) excretion in NPred-fed pigs of the present trial, especially via urine, was already pointed out in Beckmüller et al. (Citation2023). Excess N is metabolised to urea in the liver, which enters the bloodstream and gets excreted via urine (Cappelaere et al. Citation2021). Berghaus et al. (Citation2023) found a positive correlation between blood urea nitrogen and excretion of urea-N. In the present trial, the significantly lower urea concentration observed in blood serum of the NPred-fed pigs in combination with the significantly reduced daily urinary urea excretion (supplementary material S1) appears therefore conclusive. Concentration of total protein in blood serum was similar between the feeding groups, but albumin level was lower in NPred- than CON-fed pigs. However, compared to reference values for fattening pigs, no hypoalbuminemia could be diagnosed in the present study (Kixmöller Citation2004). Reduction in the albumin concentration can be caused by a reduced synthesis, for example, in the case of an acute-phase reaction (Dänicke Citation2019). Furthermore, increased excretion or altered distribution between the extra- and intravascular compartments can lead to a decrease of albumin concentration (Throop et al. Citation2004). Since albumin is synthesised in the liver from amino acids, Rothschild et al. (Citation1988) interpreted albumin concentration as an indicator for the “protein synthesis capacity of the liver”. Therefore, the lower albumin concentration in NPred-fed pigs could be considered as an indication that NPred-diets did not provide a sufficient amount of amino acids for albumin synthesis. The lower albumin concentration in blood serum in combination with the higher relative liver weight of NPred-fed pigs could also indicate an insufficient albumin synthesis, although the functional liver mass was increased. However, since the elevated relative liver weight was only observed as a statistical trend (p = 0.061), studies with a larger number of animals should be performed to verify this aspect. Further results on relative organ weights did not differ between CON- and NPred-fed pigs indicating a physiological development of all pigs. Findings are consistent with the results of Barea et al. (Citation2011), who were able to demonstrate differences in relative organ weight due to different pig genotypes, but not due to dietary crude protein (CP) reduction in pigs.
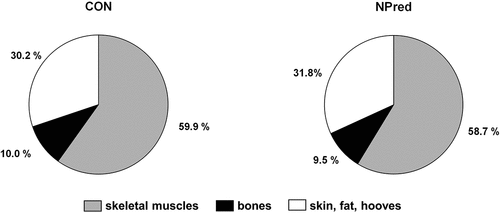
Weight of the fractions of full body analysis relative to EBW did not differ significantly between CON- and NPred-fed pigs. Additionally, relation to carcass weight () was used to compare the results on body composition of barrows of the present trial with results by Judas et al. (Citation2012), as, to our knowledge, these data represent the only means of comparison in recent literature. Pigs in the present trial showed an average 2.3% points higher skeletal muscles content (% of carcass weight). Concurrently, proportion of fraction “bones” relative to carcass weight was in average 2.4% points lower in the present experiment than in Judas et al. (Citation2012). Amount of the fraction “skin, fat, hooves” relative to carcass weight was similar compared to the data in Judas et al. (Citation2012), when summarising weights of the fractions “fat“, “rind“and “tendons“. Results of the present experiment suggest that N- and P-reduced feeding does not significantly affect chemical body composition, since nutrient concentrations did not show any significant differences between NPred- and CON-fed pigs. Lowered N concentration in the empty body of NPred-fed pigs, which was observed only as a statistical trend, was due to a significantly lowered N concentration in the fractions “blood, organs” and “bones” (supplementary material S3). Fractions “skeletal muscles“and “skin, fat, hooves“ showed similar N concentrations for both dietary groups. Reduced N concentration in fraction “blood, organs” can be explained by the reduced concentration of albumin and urea in the blood serum. Lowered N concentration in the bones of NPred-fed pigs was probably caused by a replacement with adipose tissue, as ether extract (EE) concentration in the bones was significantly higher at the same time. However, decreased N concentration in empty body of NPred-fed pigs was only observed as a statistical trend. Since only eight pigs were used for the present trial, it will be necessary to validate the data with a larger number of animals in the future. In comparison with chemical body composition of barrows after a balance trial of Haude (Citation2003), CP concentration was 7.5 and 11.9% lower and EE concentration was 15.8 and 17.1% higher in CON- and NPred-fed pigs, respectively. Lower CP concentration in combination with higher EE concentration could be an indication of an undersupply with amino acids (AA) (Martínez-Ramírez et al. Citation2008). As described in Beckmüller et al. (Citation2023), NPred-fed pigs were slightly undersupplied with precaecal digestible lysine in phase III. But NPred-fed pigs did not show an impaired growth performance compared to CON-fed pigs, which would have been expected if they had been undersupplied with the first-limiting amino acid lysine (Liao et al. Citation2015). Furthermore, decreased N retention caused by a non-ideal AA pattern in the diet could have led to the decreased body CP concentration (GfE Citation2006; Van Milgen and Dourmad Citation2015). Reduced lysine concentration in all experimental diets in fact resulted in a non-optimal AA pattern (supplementary material S4), which would explain why body CP concentration was lower in both dietary groups compared with further experiments (Haude Citation2003). Compared to other balance trials (including Haude (Citation2003)), feeding intensity in the present trial was higher, as the pigs were fed ad libitum ( = 24-hour access to feed) throughout the trial. The resulting high energy consumption could have led to an increased accretion of body fat and thus to an elevated EE concentration in both CON- and NPred-fed pigs (Kirchgeßner et al. Citation2014). However, it is important to consider LW of pigs when they are slaughtered for full body analysis. While CP concentration in the pig’s body remains relatively constant during periods of growth, EE concentration increases continuously (Gebhardt et al. Citation1981). Therefore, it seems likely that pigs of the present trial would have had a higher CP and lower EE concentration in empty body if they had been slaughtered with a similar LW ( = 110 kg) like in Haude (Citation2003). De Oliveira et al. (Citation2023) described that “reducing the CP content is an efficient strategy to reduce nitrogen excretion without affecting body protein deposition”. This is confirmed by the similar values for N retention in CON- and NPred-fed pigs in the present trial. Level of N and P retention differed markedly compared to the results using balance method for calculation of N and P retention in the same pigs published by Beckmüller et al. (Citation2023). N and P retention was about 2 to 3 times higher using the balance method compared to full body analysis. Discrepancy between mentioned methods has already been described for N by numerous authors and was reported to be 5 to 34% (Hennig et al. Citation1982; Oslage et al. Citation1987; Haude Citation2003). On the one hand, using the balance method leads to an overestimation of N retention, mainly due to inevitable volatile N losses, which cannot be avoided despite frequent faecal collection and the use of acids (GfE Citation2006). On the other hand, N retention is underestimated using the full body analysis, as N losses during growth (e.g. skin scales, bristles, saliva) are not recorded (GfE Citation2006). Mentioned reasons for the underestimation of retention can also be transferred to P, since P losses during growth are not considered. As already discussed in the first publication on this experiment (Beckmüller et al. Citation2023), it is very likely that N and P retention calculated via balance method has been significantly increased due to ad libitum access to feed. It can therefore be assumed that ad libitum feeding has a limiting effect on the conclusiveness of retention values calculated using the balance method.
Figure 3. Fractions of full body analysis relative to carcass weight of pigs fed the control diet (CON, n = 4) and the diet with reduced nitrogen and phosphorus concentrations (NPred, n = 4).
It is well known that 95–99% of calcium (Ca) and 70–75% of phosphorus (P) are stored in the skeleton system (Pettey et al. Citation2015; Gonzalo et al. Citation2018). Similar values could be observed in the present trial as 98.3 ± 0.2 and 98.2 ± 0.1% of body Ca (p = 0.230) and 76.2 ± 1.5 and 73.8 ± 1.1% of body P (p = 0.036) were stored in bones of CON- and NPred-fed pigs, respectively. Level of P stored in the bones can be regarded as a diagnostic value for assessing long-term P supply (Rieger et al. Citation2021). Therefore, the significant feeding group difference in the percentage of body P stored in bones, confirms on the one hand the lower P intake of NPred-fed pigs, while on the other hand the results support the hypothesis that CON-fed pigs were oversupplied with P. Concerns are repeatedly raised that a reduction in dietary P and Ca concentration leads to leg weakness and lameness in fattening pigs. Therefore, dietary mineral deficiency represents one differential diagnosis for non-infectious lameness (Rieger et al. Citation2021). Jongbloed (Citation1987) showed that feeding low levels of P in the LW range between 30 and 120 kg resulted in a reduction of bone calcification (diagnosis by X-ray), but was not associated with increased occurrence of leg weakness compared to a control group. In the present study, pigs also showed neither lameness nor leg weakness. In addition, blood concentrations of P and Ca gave no evidence of P and Ca deficiency in CON- or NPred-fed pigs in the present trial (Kraft and Dürr Citation2014). But according to Rieger et al. (Citation2021), serum levels of P and Ca should only be interpreted in the context of short-term mineral deficiency. Evaluation of nutrient balance indicated that Ca absorption may not have been sufficiently downregulated in NPred-fed pigs during phases I and II (Beckmüller et al. Citation2023), which could be an explanation for increased urinary Ca excretion. Therefore, it is possible that less Ca was stored in the body during these two phases, which could not be compensated in phase III. This could explain lower Ca concentration in NPred-fed pigs (observed as a statistical trend), which was likely caused by a significantly reduced Ca concentration in the bones of the NPred-fed pigs (supplementary material S3). Gonzalo et al. (Citation2018) determined body P and Ca content in pigs between 100 and 130 kg LW using dual-energy X-ray absorptiometry method. Assuming EBW to be 94% of LW according to GfE (Citation2006), they found P concentrations between 5.05 and 5.22 g/kg EBW (excluding P-undersupplied feeding group) which is similar to observed P concentrations of the present trial. Furthermore, Gonzalo et al. (Citation2018) determined Ca concentrations between 6.60 and 6.93 g/kg EBW (excluding P-undersupplied feeding group), which are markedly lower than values from the present experiment. Using the method of full body analysis, Jongbloed (Citation1987) showed similar P and Ca concentrations (5.4 g P and 8.9 g Ca per kg EBW), but in pigs with 100 kg LW, thus somewhat lighter. Recent results published by Ruiz-Ascacibar et al. (Citation2019), showed P concentrations between 5.52 and 6.06 g/kg EBW and Ca concentrations between 8.80 and 9.67 g/kg EBW in pigs with an average LW of 120 kg, respectively. Thus, concentrations are similar or slightly higher compared to the results of the present trial. Comparison with data published by GfE (Citation2006) is difficult as there are only nine studies included going back to 1969. Hence, the comparability with contemporary fattening pigs is questionable due to the breeding progress in the meanwhile.
Data on chemical body composition of pigs, especially N and P concentration, is of crucial importance in the preparation of farmgate balances. In Germany, a concentration of 25.6 g N and 5.1 g P per kilogram of pig product is assumed (BMEL Citation2017). Since data on chemical body composition of pigs should be updated regularly (Kirchgeßner et al. Citation2014), results of the present trial provide data on the chemical body composition of contemporary barrows. Results of the present experiment showed similar values for the concentration of N and P in the empty body (23.9 ± 0.9 g N/kg EBW and 5.4 ± 0.3 g P/kg EBW). Further trials with a larger number of animals and different sexes should follow in the near future to validate the results, in particular to investigate the observed trend of lower N concentration in the empty body of NPred-fed pigs.
5. Conclusion
Results indicate that N- and P-reduced feeding does not significantly affect chemical body composition and N and P retention of contemporary fattening pigs. This study provided updated data on chemical body composition of contemporary barrows. Further studies with a larger number of animals and different sexes will be necessary to confirm and extend the present results on body nutrient concentrations and health traits, especially with focus on N concentration in the empty body, serum albumin concentration and relative liver weight. In the future, it will be essential to regularly update data on chemical body composition of pigs in order to provide pig farmers a transparent and reliable data basis for the legally mandatory preparation of farmgate balances.
SV801_Paper_II_Supplements_clean final.docx
Download MS Word (33 KB)Acknowledgments
The authors thank the staff of the Institute of Animal Nutrition, Friedrich-Loeffler-Institut (FLI) in Braunschweig for taking care of the experimental animals, for supporting the time-consuming manual separation of the fractions in the slaughterhouse and the laboratory analyses of the samples.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Supplementary material
Supplementary data for this article can be accessed at https://doi.org/10.1080/1745039X.2024.2321709
Additional information
Funding
References
- Barea R, Nieto R, Vitari F, Domeneghini C, Aguilera JF. 2011. Effects of pig genotype (Iberian v. Landrace × Large White) on nutrient digestibility, relative organ weight and small intestine structure at two stages of growth. Animal. 5(4):547–557. doi: 10.1017/S1751731110002181.
- Beckmüller E, Kluess J, Hüther L, Kersten S, Kölln M, Wilke V, Visscher C, Dänicke S, Grümpel-Schlüter A. 2023. Effects of dietary-reduced nitrogen (N) and phosphorus (P) on N and P balance, retention and nutrient digestibility of contemporary fattening pigs fed ad libitum. Arch Anim Nutr. 77(6):468–486. doi: 10.1080/1745039X.2023.2288721.
- Berghaus D, Haese E, Weishaar R, Sarpong N, Kurz A, Seifert J, Camarinha-Silva A, Bennewitz J, Chillon T, Stefanski V, et al. 2023. Nitrogen and lysine utilization efficiencies, protein turnover, and blood urea concentrations in crossbred grower pigs at marginal dietary lysine concentration. J Anim Sci. 101:1–10. doi: 10.1093/jas/skad335.
- [BMEL] Bundesministerium für Ernährung und Landwirtschaft. 2017. Verordnung über den Umgang mit Nährstoffen im Betrieb und betriebliche Stoffstrombilanzen (Stoffstrombilanzverordnung - StoffBilV). BGBl. I(79):3942–3959.
- Cappelaere L, Le Cour Grandmaison J, Martin N, Lambert W. 2021. Amino acid supplementation to reduce environmental impacts of broiler and pig production: a review. Front Vet Sci. 8:1–14. doi: 10.3389/fvets.2021.689259.
- Council Directive 91/676/EEC. 1991. Council directive of 12 December 1991 concerning the protection of waters against pollution caused by nitrates from agricultural sources (91/676/EEC). Off J Eur Union. L375:1–8.
- Dänicke S. 2019. Akute-Phase-Reaktion beim Schwein: Bedeutung für die Diagnostik von Entzündungen und Wechselwirkungen mit der Ernährung. Proceedings of the 15th Symposium on Swine and Poultry Nutrition [15. Tagung Schweine- und Geflügelernährung]; Nov 19–21; Wittenberg/Halle, Germany. Halle: Zeyner & Kluth. p. 59–65.
- De Oliveira MJK, Melo ADB, Marçal DA, Valini D, Silva CA, Veira AM, Fraga AZ, Arnaut PR, Campos PHRF, Dos Santos LS, et al. 2023. Effects of lowering dietary protein content without or with increased protein-bound and feed-grade amino acids supply on growth performance, body composition, metabolism, and acute-phase protein of finishing pigs under daily cyclic heat stress. J Anim Sci. 101:1–16. doi: 10.1093/jas/skac387.
- [DLG] Deutsche Landwirtschafts-Gesellschaft. 2019. Leitfaden zur nachvollziehbaren Umsetzung stark N-/P-reduzierter Fütterungsverfahren bei Schweinen: DLG-Merkblatt 418. 4th überarbeitete Auflage. Frankfurt am Main: DLG e. V.
- Dourmad J-Y, Jondreville C. 2007. Impact of nutrition on nitrogen, phosphorus, Cu and Zn in pig manure, and on emissions of ammonia and odours. Livest Sci. 112(3):192–198. doi: 10.1016/j.livsci.2007.09.002.
- Gebhardt G, Anke M, Berger H, Bergner H, Jeroch H, Poppe S, Reinisch F, Schiemann R. 1981. Tierproduktion - Tierernährung. 1st ed. Auflage. Berlin: VEB Deutscher Landwirtschaftsverlag DDR.
- [GfE] Gesellschaft für Ernährungsphysiologie. 2006. Empfehlungen zur Energie- und Nährstoffversorgung von Schweinen. Frankfurt am Main, (Germany): DLG-Verlag.
- Gonzalo E, Létourneau-Montminy MP, Narcy A, Bernier JF, Pomar C. 2018. Consequences of dietary calcium and phosphorus depletion and repletion feeding sequences on growth performance and body composition of growing pigs. Animal. 12(6):1165–1173. doi: 10.1017/S1751731117002567.
- Haude I. 2003. Untersuchungen zu Auswirkungen einer unterschiedlichen Threoninversorgung von Mastschweinen auf N-Bilanz, Zusammensetzung des Ansatzes sowie die Leistung (Zunahmen, Energieaufwand) [ Dissertation]. Tierärztliche Hochschule Hannover, Germany. https://elib.tiho-hannover.de/servlets/MCRFileNodeServlet/etd_derivate_00002670/haudei_2003.pdf.
- Hennig U, Wünsche J, Meinl M, Borgmann E, Kreienbring F, Bock H-D. 1982. Einfluß einer abgestuften Proteinversorgung bei hohem Energieniveauf auf die Mastleistung sowie den Ansatz und die Verwertung von Futterenergie, Protein und Aminosäuren durch weibliche Mastschweine: 3. Mitteilung Mittels N-Bilanz und Schlachtkörper-N-Analyse bestimmter N-Ansatz sowie N- und Lysin-Umsatz. Arch Anim Nutr. 32:637–649.
- Jongbloed AW 1987. Phosphorus in the feeding of pigs: effect of diet on the absorption and retention of phosphorus by growing pigs [ Dissertation]. Landbouwuniversiteit te Wageningen, The Netherlands. https://edepot.wur.nl/201468.
- Judas M, Branscheid W, Höreth R. 2012. Neue Ergebnisse zur Variabilität der Gewebeanteile beim Schwein. Mitteilungsblatt Fleischforschung Kulmbach. 51(195):1–16.
- Kirchgeßner M, Stangl GI, Schwarz FJ, Roth FX. 2014. Tierernährung: Leitfaden für Studium, Beratung und Praxis. 14th ed. Frankfurt/Main, Germany: DLG-Verlag.
- Kixmöller M 2004. Labordiagnostische Referenzbereiche bei unterschiedlichen Schweinerassen sowie histopathologische und immunhistochemische Untersuchung von Gehirnen älterer Sauen und Eber auf transmissible spongiforme Enzephalopathie im Rahmen der TSE-Studie [ Dissertation]. Ludwig-Maximilians-Universität München, Germany. doi: 10.5282/edoc.2198.
- Kraft W, Dürr UM. 2014. Klinische Labordiagnostik in der Tiermedizin. 7th ed. aktualisierte und erweiterte Aufl. Stuttgart, Germany: Schattauer Verlag.
- Liao SF, Wang T, Regmi N. 2015. Lysine nutrition in swine and the related monogastric animals: muscle protein biosynthesis and beyond. Springerplus. 4:147. doi: 10.1186/s40064-015-0927-5.
- Lüdke H, Schöne F, Geinitz D, Brys J. 1990. Untersuchungen zur Phosphorversorgung von Schweinen im Lebendmasseabschnitt 11-38 kg. 2. Mitteilung. Der Phosphor-, Kalzium- und Ascheansatz im Skelett- und Weichgewebehomogenat bei P-Unterversorgung und bei Einsatz von zwei verschiedenen Futterphosphaten sowie unterschiedlicher Rationsgestaltung. Arch Tierernahr. 40(11–12):1085–1095.
- Martínez-Ramírez HR, Jeaurond EA, de Lange CFM. 2008. Dynamics of body protein deposition and changes in body composition after sudden changes in amino acid intake: I Barrows. J Anim Sci. 86:2156–2167. doi: 10.2527/jas.2007-0235.
- Oliveira M, Machado AV. 2013. The role of phosphorus on eutrophication: a historical review and future perspectives. Environ Technol Rev. 2(1):117–127. doi: 10.1080/21622515.2013.861877.
- Oslage HJ, Farries FE, Fliegel H. 1987. Über die Exaktheit von Stickstoffbilanzen bei langfristigen Stoffwechselversuchen. Arch Anim Nutr. 37(11):1021–1028. doi: 10.1080/17450398709428269.
- Otten C, Berk A, Müller S, Weber M, Dänicke S. 2013. Influence of dietary amino acid level on chemical body composition and performance of growing-finishing boars of two sire lines. Arch Anim Nutr. 67(6):477–491. doi: 10.1080/1745039X.2013.858896.
- Pallauf J, Walz OP. 1988. Untersuchung zum Einfluß von Zulagen an Fumarsäure und Citronensäure bei unterschiedlicher Fütterungsintensität auf Verdaulichkeit der Rohnährstoffe und N-Bilanz beim frühentwöhnten Ferkel. Landwirtschaftliche Forschung. 41:109–119.
- Pettey LA, Cromwell GL, Jang YD, Lindemann MD. 2015. Estimation of calcium and phosphorus content in growing and finishing pigs: whole empty body components and relative accretion rates. J Anim Sci. 93(1):158–167. doi: 10.2527/jas.2014-7602.
- Pettigrew JE, Esnaola MA. 2001. Swine nutrition and pork quality: a review. J Anim Sci. 79:E316. doi: 10.2527/jas2001.79E-SupplE316x.
- Regulation (EU) No 1308/2013. 2013. Regulation (EU) No 1308/2013 of the European parliament and of the council of 17 December 2013 establishing a common organisation of the markets in agricultural products and repealing council regulations (EEC) No 922/72, (EEC) No 234/79, (EC) No 1037/2001 and (EC) No 1234/2007. Off J Eur Union. L347:671–854.
- Rieger H, Ratert C, Wendt M, Schwennen C, Kamphues J. 2021. Comparative study on the chemical composition of different bones/parts of bones in growing pigs differently supplied with inorganic phosphorus and phytase. J Anim Physiol Anim Nutr. 105(Suppl 2):106–118. doi: 10.1111/jpn.13636.
- Rothschild MA, Oratz M, Schreiber SS. 1988. Serum albumin. J Hepatol. 8(2):385–401. doi: 10.1002/hep.1840080234.
- Ruiz-Ascacibar I, Stoll P, Bee G, Schlegel P. 2019. Dynamics of the mineral composition and deposition rates in the empty body of entire males, castrates and female pigs. Animal. 13(5):950–958. doi: 10.1017/S1751731118002495.
- Ruiz-Ascacibar I, Stoll P, Kreuzer M, Boillat V, Spring P, Bee G. 2017. Impact of amino acid and CP restriction from 20 to 140 kg BW on performance and dynamics in empty body protein and lipid deposition of entire male, castrated and female pigs. Animal. 11(3):394–404. doi: 10.1017/S1751731116001634.
- Schulin-Zeuthen M, Kebreab E, Gerrits WJJ, Lopez S, Fan MZ, Dias RS, France J. 2007. Meta-analysis of phosphorus balance data from growing pigs. J Anim Sci. 85(8):1953–1961. doi: 10.2527/jas.2006-715.
- Throop JL, Kerl ME, Cohn LA. 2004. Albumin in health and disease: causes and treatment of hypoalbuminemia. Compendium On Continuing Education For The Practising Veterinarian. 26(12):940–948.
- Van Beek CL, Brouwer L, Oenema O. 2003. The use of farmgate balances and soil surface balances as estimator for nitrogen leaching to surface water. Nutr Cycling Agroecosyst. 67(3):233–244. doi: 10.1023/B:FRES.0000003619.50198.55.
- Van den Broeke A, Cuyper de C, Kress K, Stefanski V, Škrlep M, Čandek-Potokar M, Maribo H, Millet S. 2022. The importance of pigs’ castration strategy on carbon footprint of feed intake, nitrogen and phosphorus efficiency under different management conditions. Animal. 16(12):100669. doi: 10.1016/j.animal.2022.100669.
- Van Milgen J, Dourmad J-Y. 2015. Concept and application of ideal protein for pigs. J Anim Sci Biotechnol. 6(1):15. doi: 10.1186/s40104-015-0016-1.
- [VDLUFA] Verband Deutscher Landwirtschaftlicher Untersuchungs- und Forschungsanstalten. 2012. VDLUFA-Methodenbuch: Band III Die chemische Untersuchung von Futtermitteln. 3rd ed. Auflage. Darmstadt (Germany): VDLUFA-Verlag.
- Weniger JH, Funk K. 1953. Untersuchungen über den Calcium- und Phosphorgehalt ganzer Schweinekörper. Archiv für Tierernaehrung. 3:325–341. doi: 10.1080/17450395309428371.
- Zomeño C, Gispert M, Čandek-Potokar M, Mörlein D, Font-I-Furnols M. 2023. A matter of body weight and sex type: pig carcass chemical composition and pork quality. Meat Sci. 197:109077. doi: 10.1016/j.meatsci.2022.109077.