
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
The aim was to determine ileal endogenous nitrogen losses (ENL) and true ileal N-digestibility (TD-N) under non-steady-state conditions of the 15N-isotope dilution technique (15N-IDT), using diets generating low and high ENL and compare results to those obtained under steady-state conditions. Twelve growing pigs (mean LW 22.4 kg) fitted with a post-valve T-caecum cannula were fed an enzyme-hydrolysed casein (EHC)-based diet or an EHC diet + 4% quebracho tannins (QT) and were labelled via continuous 15N-leucine i.v. infusion or twice daily oral 15N-leucine administration. Digesta were collected daily over three consecutive hours with blood plasma sampled on the four consecutive days after cessation of 15N-labelling. There was a significant effect of sampling day on the dilution factor. Endogenous N losses were significantly lower for the EHC than the EHC+QT diet (2.41 vs. 8.69 g/kg DMI), while no significant effect of sampling day was observed. The TD-N of the EHC+QT diet did not differ from the TD-N of the EHC diet (95.1 vs. 92.0%). A significant effect of sampling day was observed for TD-N with day 1 and 2, being higher than day 4. Non-steady-state conditions overestimated ENL by 25–28% as compared to 3 h collections in steady-state conditions, but the relative overestimation was similar for the EHC diet as for the EHC+QT diet. TD-N did not differ significantly compared to 12 h steady-state measurements, but comparison to 3 h steady-state measurements showed that non-steady-state conditions overestimated TD-N for the EHC+QT diet by 9%. However, on day 4 this overestimation disappeared. Using the 15N-IDT during non-steady-state conditions can provide valuable additional data on endogenous N losses and TD-N.
1. Introduction
A common method to measure total ileal endogenous nitrogen losses (ENL) and true ileal nitrogen digestibility (TD-N) is the 15N-isotope dilution technique (15N-IDT). In this method, either the animal or the diet is labelled with a stable isotope, often 15N-leucine, and ileal ENL are determined from the ratio of labelled N in ileal digesta and in a precursor pool (Souffrant et al. Citation1981; de Lange et al. Citation1990). It is assumed that 15N in TCA-soluble plasma, which is derived from 15N-leucine and 15N-amino acids enriched by transamination, is representative of the endogenous amino acid pool (Leterme et al. Citation1998).
An often stated prerequisite for the 15N-IDT is a steady-state of labelling of the precursor pool (Nyachoti et al. Citation1997), although plateau values are not always reached (Lien et al. Citation1997; Steendam et al. Citation2004) and plasma enrichments fluctuate between meals (Steendam et al. Citation2004). Tamminga et al. (Citation1995) suggest that when the dilution factor (DF), i.e. the ratio between enrichment of the precursor pool and enrichment of ileal digesta, is constant, a steady-state may not be essential. A study in rats showed that the ratio of specific activity of ileal digesta and the precursor pool consisting of either plasma fractions or small intestinal tissue was relatively constant during a 2–10 d 3H-leucine infusion period (Moughan and Buttery, unpublished in (Moughan et al. Citation1992)). The same was found in sheep, where the ratio of 15N-enrichment of digesta in several parts of the small intestine to that of the precursor pool, after cessation of 15N infusion, appeared to remain relatively constant over days (Van Bruchem et al. Citation1997; Van Bruchem, unpublished in (Tamminga et al. Citation1995)).
In the present study, the ratio of 15N-enrichment of TCA-soluble plasma to the 15N-enrichment of ileal digesta was investigated in growing pigs under non-steady-state conditions. From these data, ENL and TD-N were calculated and compared to values determined during steady-state conditions. Animals were labelled with 15N-leucine using either continuous i.v. infusion or twice daily oral administration of label with the meal. Measurements of 15N-enrichments of TCA-soluble plasma and ileal digesta were conducted for four consecutive days after cessation of label administration of the study of Steendam et al. (Citation2004).
Endogenous gut losses depend on several factors including dietary factors such as tannins. Condensed tannins interact with dietary and endogenous N such as salivary proteins, and the gastrointestinal mucosa and epithelium, lowering N-digestibility and increasing ENL (Jansman Citation1993; Nyachoti et al. Citation1997; Shimada Citation2006). To test if the level of ENL influences the effect of the non-steady-state conditions on ENL estimates, a diet based on EHC (low ENL) and an EHC-diet with added condensed tannins in the form of quebracho extract (quebracho tannin, QT; high ENL) were used.
We hypothesise that determination of ENL and TD-N in pigs directly after cessation of continuous labelling yields estimates comparable to steady-state conditions of the 15N-IDT. Therefore, the study aimed to determine ENL and TD-N under non-steady-state conditions, using diets generating low and high ENL and compare results to those obtained under steady-state conditions.
2. Materials and methods
2.1. Ethical approval
Ethical approval was provided by the TNO Committee for Animal Welfare (No. 71-8A; 8 December 1994).
2.2. Experimental design
The study was conducted directly after the experiment described by Steendam et al. (Citation2004) with animal care, procedures, sampling methods and diet composition as described in the latter publication. Briefly, 12 growing pigs (LW 22.4 ± 1.1 kg) fitted with a post-valve T-caecum cannula, were fed an EHC based diet or an EHC diet + 4% QT in a cross-over design. Animals were labelled via continuous 15N-leucine i.v. infusion or twice daily oral 15N-leucine administration. Plasma was sampled at 9:00, 14:00 and 19:00 h, analysed for 15N-enrichtment of TCA-soluble plasma, and a daily average of 15N-plasma enrichment was calculated. Endogenous N losses and TD-N were determined, based on a pooled sample from 12 h ileal digesta collections from day 8–10 of each period of the cross-over. Digesta in the second period (P2) was pooled so that a separate analysis of digesta collected from 10:00 to 13:00 h on day 10 was possible. One pig (in the oral tannin group) was euthanised on day 9 of the second period due to problems with the cannula, thereby continuing the trial with 11 pigs.
The start of the current study was at the end of day 10 of the second period in the study of Steendam et al. (Citation2004), when the pigs continued to receive their allocated diets but without i.v. or oral 15N-leucine administration. Pigs were kept on these diets for four consecutive days with sample collection continuing during this time. In the current study, pigs were fed at 8:00 and 15:00 h with feed mixed with water (1:1.25, w:w), provided at 2.6 times energy required for maintenance (Agricultural Research Council Citation1981). Diet composition was as reported previously ( and Steendam et al. Citation2004). An overview of the experiment is provided in Table 1 of the Supplemental Material.
Table 1. Composition of the control and quebracho tannin dietsǂ.
2.3. Isotope administration and sampling
The oral dosed pigs of Steendam et al. (Citation2004) received the final dose of 15N-leucine on day 10 of P2 with their 08:00 h feed, while 15N-leucine i.v. infusion to the other pigs was ceased at 20:00 h. The period of sample collection of digesta was decreased from 12 h in the preceding study, to 3 h (10:00–13:00 h) on all four days after termination of 15N-labelling. Blood plasma was obtained between 13:00 and 13:30 h.
2.4. Chemical analyses
Chemical analysis for dry matter, N, chromium oxide and 15N-enrichments in ileal digesta and TCA-soluble plasma samples was conducted as previously described (Steendam et al. Citation2004), except for N in ileal digesta for which total N values were obtained from the isotope ratio mass spectrometer analyses.
2.5. Calculations and statistics
Calculations and statistical analyses were performed using SAS® software v 9.4 (SAS Institute, Cary NC). Calculations of apparent and true ileal N-digestibility (AD-N, TD-N) and ENL were conducted as described by Steendam et al. (Citation2004) with the DF calculated per day, instead of an average over three days.
Data were tested for outliers, with outliers defined as values outside three times the interquartile range below Q1 or above Q3, using the UNIVARIATE procedure of SAS®. For 15N-enrichments of plasma and digesta, values were compared within animals as well as within sampling day, yielding no outliers. For AD-N, TD-N and ENL, no outliers were detected within Day or within Day × Diet.
Change in 15N-enrichment per diet per sample type (plasma or ileal digesta) over time was modelled using a single-phase exponential decay curve, both with and without day zero included (SAS®, NLIN procedure, Marquardt method):
where a is the asymptotic value of the 15N-enrichment (atom%excess, set to be > 0), k the rate constant (d−1) representing a constant relative rate of decay and t the time of sampling expressed as day (0–4) after ceasing 15N-administration. Pseudo R2 was calculated as 1-(SSres/SScortot).
Statistical analyses for AD-N, DF, ENL and TD-N were performed using the MIXED procedure of SAS® with Animal, Diet, Method of 15N-administration and Day of sampling as class variables. Fixed effects were Diet, Method of 15N-application and Day of sampling and their two-way interactions. Day of sampling was used as the repeated factor with animal as subject.
The average values per animal over day 1–4 were calculated and comparisons were made with data of Steendam et al. (Citation2004) of P2 (steady-state). For this comparison, the d 1–4 average of the current study (Trial 1) and parts of Steendam et al. (Citation2004), were all viewed as separate trials. Comparison 1 involved the values obtained during the 12 h collections during steady-state conditions over day 8–10 (P2 only, Trial 2) and comparison 2 with values obtained from 3 h ileal collections on day 10 (10:00 to 13:00 h; plasma at 14:00 h) of P2 (Trial 3). A third comparison involved the 12 h ileal digesta collections during steady-state conditions on day 8–10 (P2) and the 3 h ileal digesta collections on day 10 of that same period. Animal, Diet, Method of 15N-administration and Trial were defined as class variables (MIXED procedure, SAS®). Fixed effects were Diet, Method of 15N-administration, and their two-way interactions. Trial was used as the repeated factor with Animal as subject.
Covariance structure was chosen for the lowest values for the AICC criterium combined with simplicity of the structure. Differences were considered significant at p < 0.05 and a trend defined as 0.05 ≤ p < 0.10. Multiple comparisons between treatments were determined using the Tukey-Kramer method when significant effects were detected. Variables and residuals of the statistical analyses were tested for normality using the UNIVARIATE procedure and the Kolmogorov-Smirnov criterion.
3. Results
15N-enrichments of TCA-soluble plasma and ileal digesta declined sharply in the first day after ceasing the label administration, with enrichment of plasma showing a sharper decline than that of ileal digesta. The difference in the rate of enrichment decline between blood plasma and digesta resulted in change of the ratio, which was most noticeable between day 0 and 1. From day 1 onwards, the decline for both plasma and digesta, but especially plasma, was less pronounced.
Exponential decay curves fitted to the EHC and the EHC+QT groups as a whole are provided in the Supplemental Material.
No significant effect of method of 15N-administration preceding the experiment was detected for any of the N-digestibility or ENL variables (). There were also no significant interactions between Method and Diet or between Method and Day of sampling.
Table 2. Apparent (AD-N) and true (TD-N) ileal nitrogen digestibility, dilution factor and ileal endogenous nitrogen losses (ENL) in pigs fed an enzyme-hydrolysed casein (EHC) and EHC+quebracho tannin (QT) diet determined over a 4-d period after 15N labelling.
The AD-N was significantly higher for the EHC than EHC+QT diet ( panel a and ). There was no significant effect of Day of sampling and no interaction between Diet and Day of sampling.
Figure 1. Daily apparent nitrogen (N) digestibility (AD-N, panel a), dilution factor (DF, panel b), endogenous N losses (ENL, panel c) and true N-digestibility (TD-N, panel d) at the terminal ileum of pigs fed an enzyme-hydrolysed casein (EHC, □) and EHC+quebracho tannin (QT, ■) diet, determined over the four subsequent days directly after ceasing 15N labelling. Values are least square means ± standard error. Different subscripts within panels represent significant differences (p < 0.05) between diets (panels a and c) or days (panels b and d).
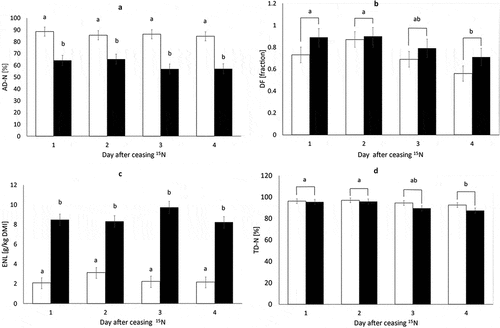
The DF, calculated from the daily measurements of plasma and ileal digesta enrichment and indicating the fraction of endogenous N in the total ileal N, did not differ significantly between the EHC and EHC+QT group (). Day of sampling had a significant effect on the DFs. The average DF was the highest on day 2 (0.88) and the lowest on day 4 (0.63) ( panel b). There was no interaction between Diet and Day of sampling.
Endogenous N losses were significantly lower for the EHC than EHC+QT diet (). For the ENL, no significant effect of sampling day was observed as illustrated in (panel c) for ENL in g/kg DMI, but similar fluctuations were found for ENL expressed as g/d. There was, however, a trend for interactions between Diet and Day of sampling.
The TD-N was not significantly different between the EHC and EHC+QT diet (). A significant effect of sampling day was observed for TD-N with day 1 and 2, being higher than day 4 ( panel d).
4. Discussion
The 15N-IDT allows estimation of ENL and thus calculation of TD-N, for feedstuffs other than N-free diets or diets with a high digestible protein source such as synthetic amino acids, EHC or casein. It is based on the fact that there is a flux between various protein pools within the body. The 15N-IDT with continuous infusion of 15N-leucine does, however, have certain limitations, including uncertainties concerning the precursor pool (Moughan et al. Citation1992), rapid label recycling (Leterme et al. Citation1996; Ravindran Citation2016), uneven distribution of label within precursor pool (Lien et al. Citation1997; Nyachoti et al. Citation1997; Leterme et al. Citation1998) and relative contributions of different endogenous sources possibly varying with diet (Nyachoti et al. Citation1997). For accurate ENL determination, steady-state of labelling of the precursor pool is often seen as a prerequisite (Nyachoti et al. Citation1997). Here we provide information regarding the requirement of a steady-state in the precursor pool, within the given limitations of the 15N-IDT.
4.1. Method of tracer administration
We detected no effect of Method of 15N-administration preceding the experiment, on any of the N-digestibility or ENL variables (). There were also no significant interactions between Method and Diet or between Method and Day of sampling. This corresponds with previous findings on oral 15N-administration by Steendam et al. (Citation2004).
4.2. Non-steady-state conditions
4.2.1. Day to day differences
The ENL estimates did not differ between days of measurement (, panel c) indicating that a single 3 h measurement is sufficient to determine ENL. Apparent ileal N-digestibility did not differ between days either, but for the QT treatment on days 3 and 4, AD-N was numerically lower. Furthermore, the DF and the TD-N showed a significant day effect, meaning that for TD-N determination it may be important to measure over multiple days. The day effect can either be attributed to random differences between days but, in the case of DF and TD-N, it could also have been caused by fluctuations in the difference in the decrease in plasma and ileal digesta 15N-enrichments, caused by the absence of 15N-label administration.
4.3. Comparison to steady-state conditions
To evaluate how accurate the estimates for ENL and TD-N obtained under non-steady- state conditions are, we compared these data to data from Steendam et al. (Citation2004). In the present study, the diets and animals were identical to those in Steendam et al. (Citation2004) and only data of the immediately preceding period was included for the comparison. Steendam et al. (Citation2004) showed that the labelling of TCA-soluble plasma is not constant during the day as a consequence of feeding distinct meals and a daily average was calculated and compared to enrichment values of pooled ileal digesta. From day 9 onwards, there were no differences between average daily plasma 15N-enrichments and it can be concluded that a steady-state between days was reached (Steendam et al. Citation2004).
Values for the 12 h collections by Steendam et al. (Citation2004) were determined from pooled ileal digesta samples collected between 8:00 and 20:00 h on d 8–10 and from overall mean values of plasma samples which were taken three times a day during the 3 d collection period. In the present study, ileal digesta were collected during 3 h to minimise the burden on the animals and the DF was calculated from one timepoint for plasma enrichment instead of an average over three timepoints. For the comparison, average values of day 1–4 of the present study were calculated.
The EHC+QT diet resulted in a lower AD-N () compared to the value obtained from pooled 12 h collections during continued 15N-leucine administration, while no difference was found for the EHC diet. Comparison between either steady-state or non-steady-state 3 h collections vs. 12 h steady-state collections, showed that AD-N for the EHC+QT diet was 9 to 10 ppt higher during the 12 h collection period (; p = 0.017 and 0.007). Comparison of 3 h digesta collection in non-steady-state to 3 h in steady-state, showed no significant difference for AD-N (). The differences in AD-N are not dependent on a 15N-steady-state condition but might be explained from the difference in the length of the collection period between both studies. Collecting for a shorter period was possibly more affected by fluctuations in chromium passage rate or diurnal variation in the flow of total N and ENL.
Table 3. Apparent (AD-N) and true (TD-N) ileal nitrogen digestibility, dilution factor and ileal endogenous nitrogen losses (ENL) in pigs fed an enzyme-hydrolysed casein (EHC) and EHC+quebracho tannin (QT) diet determined over the four subsequent days (non-steady-state) directly after ceasing 15N labelling and determined from the combined 3 d, 12 h collections and the 3 h collection on day 10 of Steendam et al. (Steendam et al. Citation2004, steady-state)ǂ.
Table 4. Significanceǂ,§ of the difference in apparent (AD-N) and true (TD-N) ileal nitrogen digestibility, dilution factor and ileal endogenous nitrogen losses (ENL) in pigs fed an enzyme-hydrolysed casein (EHC) and EHC+quebracho tannin (QT) diet determined over the four subsequent days (non-steady-state) directly after ceasing 15N labelling non-steady-state and determined from the combined 3 d, 12 h collections and the 3 h collection on day 10 of Steendam et al (Steendam et al. Citation2004, steady-state).
The rate of enrichment decline differed between blood plasma and digesta resulting in a change of the ratio of enrichments, i.e. the DF. This difference was most noticeable going from day 0 to 1. After day 1, the ratio stabilised as can be seen by comparing the fitted curves with and without day 0 (Supplemental Material Figure a and b). This is possibly caused by a lag time between plasma and digesta enrichment.
The DFs in the present study were higher compared to 12 h collections during the steady-state, for both the EHC and EHC+QT group (; +34 and + 19% resp.), but the difference was only significant for the first. Compared to 3 h collections during the steady-state, the non-steady-state led to a 51 and 32% higher DF for the EHC and EHC+QT diet, respectively (). For DF, and variables calculated from the DF (ENL, TD-N), differences can be related to the length of the collection period (3 vs. 12 h) but also to non-steady-state conditions and/or time of blood sampling.
Large differences were seen in the ENL for the EHC+QT diet, which was 53% higher in the present study compared to values obtained from pooled 12 h collections of Steendam et al. (Citation2004) (). For the EHC diet, ENL were 17% higher in the former, but this was not significant. Comparison of the current study (3 h, non-steady state) to 3 h digesta collection in steady-state (Steendam et al. Citation2004), showed 28% higher ENL for the EHC+QT diet in the present study (). This difference was, however, not as large as compared to the 12 h steady-state collections. For the EHC diet, the current study yielded 25% higher values than the 3 h collection during the steady-state. These differences were not significant, but show that the relative overestimation was similar for the EHC as for the EHC+QT diet, indicating that the ranking for ENL of different diets may be unaffected by non-steady-state conditions.
Overall, the resulting TD-Ns for the current study for both diets were not different to 12 h values obtained by Steendam et al. (Citation2004) (). Comparison of the current study to day 10 of the latter study showed that TD-N for the EHC diet did also not differ between 3 h collections during the non-steady vs. steady-state (92.3 vs. 90.7%, p = 0.214). However, for the EHC+QT diet, TD-N was 9% higher for 3 h collections under non-steady-state conditions compared to values obtained by 3 h collections in the steady-state (92.3 ± 2.11 vs. 85.1 ± 2.00% (p = 0.029), ). panel d, on the other hand, shows that on day 4 (TD-N = 87.4 ± 1.48%) this overestimation disappeared, indicating that for estimation of TD-N it may be possible to measure some days after ceasing 15N-administration.
Despite the overestimation of ENL on both diets and of TD-N on the EHC+QT diet under non-steady-state conditions, the marked effect of QT on ENL remained significant on subsequent days unlike the effect of QT on TD-N. This indicates that non-steady state conditions can be used to show a (lack of) diet effect on ENL and TD-N.
4.3.1. Length and timing of collection period
Souffrant et al. (Citation1981) found that endogenous N flows came in two peaks with the first peak at 4–8 h after feeding. A second peak, occurring after 16–20 h, may have originated from a bout of drinking at 12 h after feeding. Hodgkinson et al. (Citation2002) found a relative stable ratio between ileal ENL and chromium flow during the day, with the exception of larger variations in the 4 h after feeding. In their study, ENL started to rise at 4 h after the morning meal with the highest contributions after 8 h after the morning meal and 9 h after the afternoon meal. Both studies (Souffrant et al. Citation1981; Hodgkinson et al. Citation2002) seem to show that it may be better for short collection periods to start a number of hours after feeding, e.g. 4 h, which is in line with Kim et al. (Citation2016). Using chromic oxide as a marker, they found that 4–6 h of collection at 4–6 h after the start of feeding, may provide representative samples for measurement of basal endogenous losses and CP-digestibility. However, we collected digesta from 2 h after feeding and did not find significant differences for DF and for ENL per kg DMI or d when comparing the 3 d 12 h collection period to the 3 h collection period on day 10, so the effect of a later collection period may be limited when estimating ENL.
4.3.2. Time of blood sampling
Steendam et al. (Citation2004) showed that for animals receiving a continuous i.v. 15N-leucine infusion, sampling at 14:00 h gave values for plasma enrichment that were similar to the daily average. In the current study, sampling of blood occurred between 13:00 and 13:30 h, which immediately followed the sampling period of digesta. Simon et al. (Citation1986) suggest a lag time between plasma and digesta of 1 h, meaning sampling of plasma should have been done earlier to correspond with the digesta collection period. Since enrichment of plasma without 15N-administration is in constant decline, the actual plasma enrichment during the digesta collection period may have been somewhat underestimated and, therefore, DF and ENL may have been slightly overestimated.
4.4. Level of ENL and effect of quebracho
The level of ENL of 2.41 g N/kg DMI for our pigs (22.4 ± 1.1 kg LW) on the EHC diet (180 g/kg) is similar to values in the literature (1.01 to 3.7 g N/kg DMI) for pigs between 20–50 kg receiving diets containing 100–114 g/kg EHC (Butts et al. Citation1993, Citation1993; Leterme et al. Citation1996; Hodgkinson et al. Citation2000, Citation2003). However, for pigs between 10–33 kg LW fed diets containing 180–200 g/kg EHC, higher values (3.12 to 5.74 g N/kg DMI) have been reported (Schulze et al. Citation1995; Hodgkinson et al. Citation2000). Differences in level of ENL could be related to differences in feed intake (Butts et al. Citation1993), dietary fibre content (Nyachoti et al. Citation1997), or molecular weight distribution of the EHC. Comparing diets with either 211 g/kg 15N-casein or 15N-hydrolysed casein in 34 kg LW pigs, Deglaire et al. (Citation2019) found similar ENL of 1.91 and 1.83 g N/kg DMI, respectively. In another study (Miner-Williams et al. Citation2009), pigs fed a casein diet had the same total ileal N flow (3.9 g N/kg DMI) as found in the present study, although they did not determine ENL.
Addition of 4% QT to the EHC diet resulted in a significant decrease in AD-N (86.3 vs. 60.8%) and a significant increase in ENL (2.31 vs. 8.31 g N/kg DMI). True ileal N digestibilities did not differ between diets (95.1 vs. 92.0%, ). Literature data on effects of tannin on ENL or N-digestibility in pigs are scarce. In 10 kg LW pigs fed a 145 g/kg casein-based diet, addition of 1.5% grape tannins resulted in an AD-N of 87.2% (Myrie et al. Citation2008). This value is considerably higher than in our study (60.7%), which is likely related to the lower inclusion level or source of tannins. However, the standardised ileal N-digestibility of the diet containing grape tannins (SID, calculated assuming 99% digestibility of casein) was 93.8%, which is comparable to the high end of the TD-N range of 85–95% in our study.
4.5. Practical application
Using the 15N-IDT during the non-steady-state condition can provide valuable additional data on ENL and may, for example, be used as a pilot study. This decreases the number of animals compared to doing two separate studies, thus improving overall welfare as well as reducing costs. Because the decay curve is expected to be a mirror image of the curve immediately after starting 15N-labelling, the latter period may possibly be used in the same way. The enrichment, however, has to be sufficiently high as small measurement errors would otherwise have a large impact on the results.
5. Conclusions and recommendation
The non-steady-state condition after cessation of labelling in the 15N-isotope dilution technique overestimated ENL. The relative overestimation was similar for the enzyme-hydrolysed casein diet as for the same diet with added quebracho tannins, with the difference in ENL between the two diets remaining significant. True nitrogen digestibility measured during the 3 h non-steady-state, yielded values not significantly different to those obtained during 12 h steady-state measurement, but comparison to the same 3 h under steady-state measurements, resulted in a 9% overestimation of TD-N for the diet with added tannins. On day 4, however, this overestimation disappeared, indicating that for estimation of TD-N it may be possible to measure some days after ceasing 15N-administration. As such, non-steady-state conditions of the 15N-isotope dilution method in pigs may be used to test for diet effects with respect to ENL and TD-N. More research, however, is recommended to determine the impact of a shorter collection period and the timing of the collection period and blood sampling.
Steendam_Supplemental_Non_revised_final_2_clean.docx
Download MS Word (374.5 KB)Acknowledgments
We thank Marianne van Westerneng-van ‘t End for chemical analyses and the late Huug Boer and the late Henk Visser for coordination of the (isotope) analyses.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Data availability statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Supplementary material
Supplementary data for this article can be accessed at https://doi.org/10.1080/1745039X.2024.2322201
Additional information
Funding
References
- Agricultural Research Council. 1981. The nutrient requirements of pigs. Farnham Royal, UK: Commonwealth Agricultural Bureaux.
- Butts CA, Moughan PJ, Smith WC, Carr DH. 1993. Endogenous lysine and other amino acid flows at the terminal ileum of the growing pig (20 kg bodyweight): the effect of protein-free, synthetic amino acid, peptide and protein alimentation. J Sci Food Agric. 61:31–40.
- Butts CA, Moughan PJ, Smith WC, Reynolds GW, Garrick DJ. 1993. The effect of food dry matter intake on endogenous ileal amino acid excretion determined under peptide alimentation in the 50 kg liveweight pig. J Sci Food Agric. 62:235–243. doi: 10.1002/jsfa.2740620306.
- de Lange CF, Souffrant WB, Sauer WC. 1990. Real ileal protein and amino acid digestibilities in feedstuffs for growing pigs as determined with the 15N-isotope dilution technique. J Anim Sci. 68:409–418.
- Deglaire A, Moughan PJ, Tomé D. 2019. A casein hydrolysate does not enhance ileal endogenous protein flows compared with the parent intact casein when fed to growing pigs. Curr Dev Nutr. 3:nzy083.
- Hodgkinson SM, Moughan PJ, Morel PCH, Reynolds GW. 2002. The diurnal pattern of ileal dry matter and endogenous ileal nitrogen flows in the growing pig. J Sci Food Agric. 82:1860–1866.
- Hodgkinson SM, Moughan PJ, Reynolds GW, James KA. 2000. The effect of dietary peptide concentration on endogenous ileal amino acid loss in the growing pig. Br J Nutr. 83:421–430.
- Hodgkinson SM, Souffrant WB, Moughan PJ. 2003. Comparison of the enzyme-hydrolyzed casein, guanidination, and isotope dilution methods for determining ileal endogenous protein flow in the growing rat and pig. J Anim Sci. 81:2525–2534. doi: 10.2527/2003.81102525x.
- Jansman AJ. 1993. Tannins in feedstuffs for simple-stomached animals. Nutr Res Rev. 6:209–236.
- Kim BG, Liu Y, Stein HH. 2016. Effects of collection time on flow of chromium and dry matter and on basal ileal endogenous losses of amino acids in growing pigs. J Anim Sci. 94:4196–4204.
- Leterme P, Monmart T, Théwis A, Morandi P. 1996. Effect of oral and parenteral N nutrition vs N-free nutrition on the endogenous amino acid flow at the ileum of the pig*. J Sci Food Agric. 71:265–271.
- Leterme P, Sève B, Théwis A. 1998. The current 15N-leucine infusion technique is not suitable for quantitative measurements of ileal endogenous amino acid flows in pigs. J Nutr. 128:1961–1968.
- Leterme P, Théwis A, François E, Van Leeuwen P, Wathelet B, Huisman J. 1996. The use of 15N-labeled dietary proteins for determining true ileal amino acid digestibilities is limited by their rapid recycling in the endogenous secretions of pigs. J Nutr. 126:2188–2198.
- Lien KA, Sauer WC, Mosenthin R, Souffrant WB, Dugan ME. 1997. Evaluation of the 15N-isotope dilution technique for determining the recovery of endogenous protein in ileal digestion of pigs: effect of dilution in the precursor pool for endogenous nitrogen secretion. J Anim Sci. 75:148–158.
- Miner-Williams W, Moughan PJ, Fuller MF. 2009. Endogenous components of digesta protein from the terminal ileum of pigs fed a casein-based diet. J Agric Food Chem. 57:2072–2078.
- Moughan PJ, Buttery PJ, Essex CP, Soar JB. 1992. Evaluation of the isotope dilution technique for determining ileal endogenous nitrogen excretion in the rat. J Sci Food Agric. 58:165–172.
- Myrie SB, Bertolo RF, Sauer WC, Ball RO. 2008. Effect of common antinutritive factors and fibrous feedstuffs in pig diets on amino acid digestibilities with special emphasis on threonine. J Anim Sci. 86:609–619.
- Nyachoti CM, CFMd L, McBride BW, Schulze H. 1997. Significance of endogenous gut nitrogen losses in the nutrition of growing pigs: a review. Can J Anim Sci. 77:149–163.
- Ravindran V. 2016. Feed-induced specific ileal endogenous amino acid losses: measurement and significance in the protein nutrition of monogastric animals. Anim Feed Sci Technol. 221:304–313.
- Schulze H, Butts CA, Moughan PJ, Verstegen MWA. 1995. The 15N-isotope dilution method for determining ileal endogenous nitrogen excretion in the young (10 kg liveweight) pig. J Sci Food Agric. 69:41–50.
- Shimada T. 2006. Salivary proteins as a defense against dietary tannins. J Chem Ecol. 32:1149–1163.
- Simon O, Zebrowska T, Münchmeyer R, Bergner H, Rohrman H. 1986. Studies on the secretion of amino acids and of urea into the gastro intestinal tract of pigs. 1. Secretion of leucine into the stomach and upper part of the duodenum during continous (sic) intravenous infusion of 14C-leucine. Arch Anim Nutr. 36:5–16.
- Souffrant WB, Köhler R, Gebhardt G. 1981. Untersuchungen zur Bestimmung der endogenen N-Sekretion im Dünndarm bei Schweinen. [Endogenous N-secretion in the small intestine of pigs]. Arch Tierernahr. 31:35–43.
- Souffrant WB, Köhler R, Matkowitz R, Gebhardt G, Schmandke H. 1981. Ernährungsphysiologische Untersuchungen an Schweinen zur Beurteilung von modifizierten Proteinen. 2. Mitteilung. Bestimmung des endogenen N im Dünndarminhalt mit Hilfe der 15N-Tracertechnik. [nutritional and physiological studies of swine for the evaluation of modified proteins. 2. Determination of endogenous n in the contents of the small intestines using the 15N-tracer method]. Arch Tierernahr. 31:675–683.
- Steendam CA, Verstegen MW, Tamminga S, Boer H, van ‘t End M, Verstappen B, WR C, Visser GH. 2004. Route of tracer administration does not affect ileal endogenous nitrogen recovery measured with the 15N-isotope dilution technique in pigs fed rapidly digestible diets. J Nutr. 134:3068–3075.
- Tamminga S, Schulze H, Van Bruchem J, Huisman J. 1995. The nutritional significance of endogenous N‐losses along the gastrointestinal tract of farm animals. Archiv für Tierernaehrung. 48:9–22.
- Van Bruchem J, Voigt J, Lammers-Wienhoven TS, Schönhusen U, Ketelaars JJ, Tamminga S. 1997. Secretion and reabsorption of endogenous protein along the small intestine of sheep: estimates derived from 15N dilution of plasma non-protein-N. Br J Nutr. 77:273–286.