
Abstract
In insects, localized tissue injury often leads to global (organism-wide) delays in development and retarded metamorphosis. In Drosophila, for example, injuries to the larval imaginal discs can retard pupariation and prolong metamorphosis. Injuries induced by treatments such as radiation, mechanical damage and induction of localized cell death can trigger similar delays. In most cases, the duration of the developmental delay appears to be correlated with the extent of damage, but the effect is also sensitive to the developmental stage of the treated animal. The proximate cause of the delays is likely a disruption of the ecdysone signaling pathway, but the intermediate steps leading from tissue injury and/or regeneration to that disruption remain unknown. Here, we review the evidence for injury-induced developmental delays, and for a checkpoint or checkpoints associated with the temporal progression of development and the on-going efforts to define the mechanisms involved.
Abbreviations
| PTTH | = | prothoracicotropic hormone |
| E | = | ecdysone |
| 20E | = | 20-hydroxyecdysone |
| DILP8 | = | Drosophila Insulin-Like Peptide 8 |
Developmental Delays are Triggered by a Variety of Injuries
In insects, various types of tissue damage can trigger a systemic injury response which results in prolonged larval and/or pupal stages. For example, in Galleria, small incisions to the integument prolong larval instars by approximately one day.Citation1 In Blattella and Periplaneta, amputation of appendages results in developmental retardation.Citation2-5 In Drosophila, exposure to microwave frequency electromagnetic fields (EMF) delays the onset of pupariation.Citation6 Finally, in Drosophila, Ephestia and Galleria, damage to imaginal discs via irradiation or induction of cell death clones results in prolonged larval instars.Citation4,7-Citation17 These delays appear to be systemic as when one wing imaginal disc is damaged, growth slows or stops in the second wing disc.Citation4,9-Citation11 In most cases, development appears to arrest until the damaged tissue is healed, whereupon development commences and the emerging adult appears morphologically normal.Citation9,13,15,17
The extent of the injury-induced developmental delay is correlated with two factors: (1) the amount of damaged tissue present and (2) the developmental stage of the animal at the time injury is sustained.Citation2,3,6,8,10,11,14,16,21 There is a positive correlation between the amount of amputated or damaged tissue and the length of the molting delay.Citation2,3,6,8,10,16,18,19 For example, in Periplaneta and Blattella, molting is significantly delayed when a single limb is autotomized (self-amputated as an escape mechanism) and autotomy of a second limb causes a measurable extra delay when done simultaneously with the first autotomy.Citation2–5,19 In Drosophila, small lesions induced by temperature-sensitive cell death clones in the wing discs do not produce a measurable delay.Citation10 However, increasing the amount of lethal tissue delays both pupariation and eclosion.Citation10 There also appear to be dose-dependent effects of X-irradiation, ethyl methanesulfonate (EMS) treatment, and electromagnetic field (EMF) exposure on pupariation times, supporting the notion that the amount of damaged tissue dictates, at least in part, the length of the molting delay.Citation6,8,14,22
The duration of injury-induced developmental delays is affected not only by the amount of tissue damage present but also by the developmental stage during which the damage occurs. Damage induced by X-irradiation has different effects on developmental timing depending on the age of the animal at the time of treatment.Citation8,14 Larvae irradiated early (48–84 hours after hatching) pupariate late while those irradiated closer to the normal pupariation time often pupariate precociously.Citation8,14 Delays induced by other means also show a stage-dependent response; lesions induced by targeted cell death in imaginal discs only appear to retard pupariation when induced at or before a certain critical stage in development – around the beginning of the third larval instar.Citation10,14,17 Interestingly, damage induced via irradiation to second instar Drosophila larvae does not delay the subsequent molt but does prolong the third larval stage suggesting the presence of an injury response developmental checkpoint in the last larval instar.Citation14 The injury response checkpoint senses damaged tissue and responds by activating a series of events that inhibit developmental transitions. This checkpoint appears to be analogous to cell cycle checkpoints which sense incomplete or inaccurate cell cycle events and respond by inhibiting cell cycle transitions.Citation23
In Drosophila, we know of several critical “moments of passage” that occur during the third larval stage. These include critical weight (CW), minimum viable weight (MVW), and the mid-third instar transition (MIT). Critical weight is defined as the minimum size at which starvation no longer delays the onset of metamorphosis while minimum viable weight refers to the minimum weight required to survive the next developmental transition without additional feeding.Citation24,25 In Drosophila, CW and MVW occur early in the third larval instar.Citation26 The mid-third instar transition (MIT) is a developmental landmark that occurs during the middle of the third larval instar and is characterized by widespread changes in gene expression.Citation27 Tissue damage is associated with delayed developmental progression when induced before or after both CW and MVW has been achieved suggesting that the injury response checkpoint does not coincide with either CW or MVW.Citation14,15 Tissue damage induced via temperature-sensitive cell death induction in wing discs no longer triggers delayed development after animals progress past the MIT.Citation15 Together these studies suggest the presence of an injury response checkpoint that delays developmental transitions in response to tissue damage sustained prior to attainment of the MIT.
Regenerating Tissue May Activate the Injury Response Checkpoint
Damaged imaginal tissues and amputated appendages in many insects have the ability to regenerate through a process involving localized cell proliferation.Citation28 Regeneration of certain tissues is associated with molting delays in insects and such an influence on the molt cycle has been associated with regeneration of imaginal discs and amputated appendages.Citation11,13,29-37 Experiments in Ephestia indicate that the presence of regenerating imaginal tissue is sufficient to induce systemic developmental delays. For example, RahnCitation35 implanted fragments of wing imaginal discs into the body cavities of Ephestia larvae. The implanted fragment regenerated a complete disc and during this process, further growth of host larval structures and pupation were delayed.Citation35 Similarly, DewesCitation36 transplanted male genital imaginal disc fragments into host Ephestia larvae of various ages. Regeneration of the disc fragments was associated with delayed onset of pupation, and, in general, the greater the amount of disc regeneration that occurred, the greater the pupation delay.Citation36 No delay in development was observed when the disc fragment failed to regenerate.Citation36
In Drosophila, the larval stage during which damage is inflicted influences the amount of regeneration that occurs. Imaginal wing disc damage induced in young larvae, before the MIT, results in almost complete regeneration of wings, delayed development, and adults that are morphologically normal.Citation13,15 Damage induced in older larvae, after the MIT, results in almost no tissue regeneration, no developmental retardation, and wingless adults.Citation13,15 These studies suggest that the injury response checkpoint may signify a change in the ability of imaginal tissues to regenerate. A similar ‘regeneration critical’ period has been observed in other organisms including cockroaches and assassin bugs (Rhodnius prolixus).Citation2,21
SimpsonCitation10 proposed a direct relationship between growth of imaginal discs and the timing of pupariation. She suggested that discs that have not completed a certain amount of growth produce a signal that inhibits pupariation. In fact, there is abundant evidence that links proliferating imaginal cells with a delay in pupariation.Citation11,12,29,30,32,34,38 For example, the mutations lethal(2)giant discs (l(2)gd) and lethal(1)discs large-1 (l(1)d.lg-1) each result in extensive overgrowth of all imaginal discs and display a third larval instar which is extended by up to 9 days and often a failure to pupariate.Citation32,34 In addition to imaginal discs, regeneration of amputated appendages has also been shown to be associated with a developmental delay in some insects. For example, O’Farrell and StockCitation3,19 investigated the regeneration of appendages and its effect on molting in the larvae of Blattella germanica and Periplaneta americana. They demonstrated that removal of an appendage postponed the subsequent molt until the wound had healed and regeneration of the appendage was complete. They inferred that regenerating tissue triggers a systemic inhibition of development. These studies suggest that it may be localized cell proliferation in regenerating tissue, rather than signals produced by damaged tissue, that leads to inhibition of developmental progression. However, the developmental delay must depend on more than just dividing cells since even extensive hypertrophy of the haematopoietic system in l(1)TumCitation1 mutant larvae does not result in delayed pupariation.Citation39
The Injury Response Checkpoint Modulates Ecdysone Signaling
Ecdysteroids control the timing of developmental transitions including molts for growth (larval-larval molts), and for metamorphosis (larval-pupal and pupal-adult molts).Citation40-42 Pulses of ecdysteroids trigger a variety of developmental and reproductive processes including, but not limited to, embryonic organ development, imaginal disc morphogenesis, glue protein secretion and polytene chromosome puffing, cuticle tanning, degeneration of larval tissues during metamorphosis, proliferation of histoblasts during metamorphosis and control of oogenesis in adult females.Citation40,43-Citation56 The production of ecdysteroids by the prothoracic gland cells of the ring gland is stimulated by prothoracicotropic hormone (PTTH), a cerebral neuropeptide ().Citation57-62
Figure 1. A Proposed Model for the Injury Response Checkpoint. (A) Synthesis of Ecdysone (E) by the prothoracic cells of the larval Ring Gland is stimulated by PTTH. Citation57–62 PTTH release stimulates production of E which is released into the hemolymph. Citation60 Ecdysone is converted to its active form 20-hydroxyecdysone (20E) by enzymes expressed in target tissues where 20E triggers a series of downstream events leading to developmental progression.87 (B) Tissue damage and regenerating tissues lead to activation of JNK signaling and increased expression of dilp8. Citation22,69 DILP8 disrupts ecdysteroid production by decreasing expression of ecdysteroidogenic enzymes including dib and phm and possibly delaying the PTTH peak. Citation22,69 Tissue damage also influences ptth expression via a retinoid-dependent pathway which is separate from the DILP8 pathway. Citation14,22 Wing imaginal disc damage is associated with decreased expression of the ecdysone receptor and several ecdysone response genes, however it is unclear whether or not this is a direct effect of the injury response signals or a by-product of reduced ecdysteroid production. Citation15 Increased expression of ecdysone oxidase, which encodes an enzyme
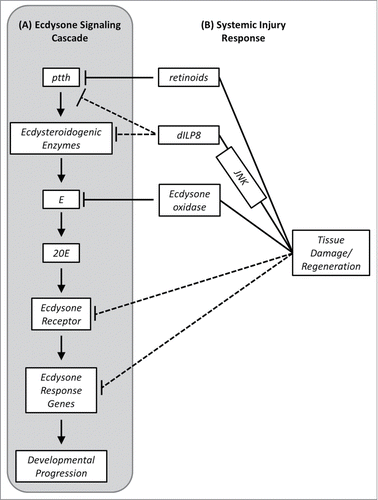
Since developmental progression is under ecdysteroid control it is not surprising that developmental perturbations elicited by injury and regeneration also have an endocrine basis and appear to be due to a disruption in the production of ecdysteroids ().Citation1,2,4,7,8,14,15,21,38 In the kissing bug, Rhodnius prolixus, leg amputation results in a temporary decrease in the hemolymph ecdysteroid titer. Abolishing nervous system activity with the neurotoxin, tetrodotoxin, delays the recovery of the ecdysteroid titer.Citation21 Recovery of the ecdysteroid titer closely parallels the recovery of electrical activity in the corpora cardiaca (CC) region of the ring gland which receives PTTH from axons of neurosecretory cells.Citation21,63 This suggests that the decrease in hemolymph ecdysteroids is due to decreased stimulation of ecdysteroidogenesis by PTTH. In Drosophila, developmental delays induced by X-irradiation are associated with a decrease in ptth expression and the delays can be suppressed via ectopic feeding of 20-hydroxyecdysone (20E) or by disrupting components of the retinoid biosynthesis pathway suggesting a role for retinoids in inhibiting PTTH production or release following tissue damage ().Citation14
Additional studies in Drosophila have demonstrated that the decrease in the hemolymph ecdysteroid titer is due to additional factors, not simply a decrease in ptth expression. Damage to wing imaginal discs also results in decreased expression of many genes encoding ecdysteroidogenic enzymes including neverland (nvd), spookier (spok), disembodied (dib) and phantom (phm) ().Citation14,15,22 Genes encoding transcription factors, including molting defecting (mld) and without children (woc), which are also required for ecdysone synthesis also show reduced expression following wing disc damage.Citation15,64,65 Another factor which contributes to the decline in circulating ecdysone is ecdysone oxidase, an enzyme which inactivates ecdysone by converting it to 3-dehydroecdysone.Citation66 Expression of ecdysone oxidase (Eo) is increased following localized cell death in the wing disc ().Citation15 These studies suggest that not only is there reduced synthesis of ecdysone in the ring gland, but circulating ecdysone may also be degraded to lower the hemolymph ecdysteroid titer. It is possible that the decreased expression of ecdysteroidogenic enzymes and ecdysone oxidase is a by-product of decreased ptth expression; however this possibility has not been fully explored.
In Drosophila larvae carrying lethal mutations including dlg [lethal (1)discs large], lgd [lethal (2)giant discs], dco [lethal (3)discs overgrown], c43 [lethal (3)c43] and fat [lethal (2)fat], all of which are characterized by imaginal disc overgrowth and developmental delays, the ecdysteroid surge that normally occurs at pupariation is absent.Citation38 No accumulation of unusual metabolites or conjugates has been detected in imaginal disc overgrowth mutants indicating that the reduced titer of ecdysteroids is primarily due to reduced ecdysone production by the ring gland rather than increased metabolic inactivation.Citation38 Implantation of wild type ring glands can suppress the pupariation delay observed in larvae homozygous for the imaginal disc overgrowth mutation lgl (lethal (2)giant larvae), indicating that the pupariation delay is caused by failure of ecdysone production or release.Citation39,67 Together these studies indicate that injured or regenerating tissues produce a signal that inhibits ecdysteroid production or secretion by the larval ring gland, and that this injury-response pathway involves regulation of the neuropeptide PTTH.
An Insulin-like Signal Mediates the Systemic Injury Response
The signal which triggers delayed development following injury appears to originate from either damaged tissues or regenerating/proliferating tissues. Studies have suggested that x-rays, which induce chromosome aberrations in dividing cells, cause the death of cells which in turn release a diffusible morphogenetic substance which results in developmental delays.Citation12,68 It was recently shown in Drosophila that the signal produced by damaged and regenerating imaginal tissues is an insulin-like peptide (DILP8), which is stimulated by c-jun-terminal kinase (JNK) signaling and is capable of disrupting the timing of developmental progression through inhibition of ecdysteroid production ().Citation22,69
Increased expression of dilp8 leads to pupariation delays and retards the expression of disembodied (dib) and phantom (phm), two genes required for ecdysone synthesis in the ring gland ().Citation22,69 These delays can be rescued by feeding larvae 20E which is consistent with the idea that injury and regeneration induced developmental disruptions have an endocrine basis.Citation22,69 However, there are conflicting reports regarding the mechanism by which DILP8 influences ecdysteroid production. ColombaniCitation69 demonstrated that ectopic expression of dilp8 in wing imaginal discs retards the onset of ptth expression in the ring gland by as much as 36 hours. In contrast, GarelliCitation22 demonstrated that dilp8 overexpression had only a minimal effect on ptth expression and suggested that a second signal may act on PTTH-secreting neurons to contribute to the decrease in ecdysteroid synthesis following tissue damage. The idea that multiple pathways inhibit development following injury is supported by the fact that cell death-induced wing disc damage inhibits PTTH synthesis and the resulting developmental delay can be enhanced by the consumption of β-carotene in the diet.Citation14,22 The effects of DILP8 on development appear to be independent of retinoids ().Citation22
Of all the insulin-like peptides tested, dilp8 is the only one that has been shown to be differentially expressed in imaginal disc tumors as well as in imaginal tissues that have been damaged through overexpression of the proapoptotic gene reaper (rpr) or EMS treatment.Citation11,22 Mutations in other insulin/insulin-like growth factor 1 (IGF1) signaling pathway components, are often associated with delays in pupariation and a decrease in body and organ size.Citation70–72 However, Garelli and colleaguesCitation22 demonstrated that IGF1 signaling was largely unaffected in larvae overexpressing dilp8 suggesting that DILP8 may function through a novel insulin-like signaling pathway to regulate ecdysone synthesis.
Remaining Questions and Future Perspectives
The mechanism by which damaged or regenerating tissues trigger developmental delays associated with a reduction in the hemolymph ecdysteroid titer remain poorly understood. As shown in the proposed injury-response checkpoint model (), the injury-response checkpoint clearly involves humoral signals including insulin-like peptides and retinoids which appear to act directly on the ring gland or on PTTH-secreting neurons.Citation14,22,69 However, the checkpoint also appears to involve signaling through the central nervous system (CNS), at least in some organisms. For example, developmental delays induced by amputation of appendages in Periplaneta and Blattella require intact nerve connections from the amputated limb to the CNS.Citation2 In contrast, delays observed following injury to wing imaginal discs in Galleria are not suppressed by either transaction of the nerve cord or brain implantation suggesting that the delays are triggered by a humoral signal.Citation1
It is currently unclear whether both larval and imaginal tissues are capable of triggering developmental delays upon injury. Several studies have found that pupariation can occur in the absence of imaginal discs and that larval tissues contain all of the necessary components to initiate metamorphosis.Citation10,73,74 It has been clearly shown that proliferating or damaged imaginal tissues can produce signals which retard development.Citation12,29,31,32,34 The effects of larval tissue injuries on developmental timing are less well characterized, however Mala and colleaguesCitation1 have demonstrated that incisions to larval integument can trigger a slight developmental delay in Galleria.
Many genetic mutations have been identified which, like physical damage, are capable of triggering a developmental delay or arrest.Citation75 For example, flies bearing mutations in known cell cycle regulators display prolonged larval stages and delays in adult eclosion.Citation76–78 Mutations that appear to specifically disrupt the development of imaginal discs also display a systemic retardation of development.Citation79,80 In fact there are numerous mutations – too many to discuss at length in this review – that are associated with developmental delays and/or arrest at various stages. It is possible that a number of these mutations activate the injury response checkpoint; however it is unknown how common this may be.
It is known that injury in insects triggers developmental delays associated with a reduction in the hemolymph ecdysteroid titer. Similarly, in vertebrates, severe injury, chronic disease and infection can each lead to developmental delays – most notably disruptions in gamete development and delayed onset of puberty.Citation81–84 As in insects, injury-induced developmental delays are often associated with changes in steroid hormone titers.Citation81,85 The exact mechanism underlying injury- and infection-induced developmental delays in vertebrates remains poorly understood but may involve signaling via cytokines released from either damaged/infected tissues or cells of the immune system.Citation83,84,86 Understanding how injured or infected tissues trigger changes in endocrine function is critical to understanding and treating various disease states and analysis of the injury-induced molting checkpoint in insects promises to provide valuable insights into the interactions between injured tissues and endocrine centers.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
References
- Malá J, Sehnal F, Kumaran AK, Granger NA. Effects of starvation, chilling, and injury on endocrine gland function in Galleria mellonella. Arch. Insect Biochem. Physiol. 1987;4(2):113-128; http://dx.doi.org/10.1002/arch.940040205
- Kunkel JG. Cockroach molting. II. The nature of regeneration-induced delay of molting hormone secretion. Biol. Bull. 1977;153(1):145-62; PMID:889943; http://dx.doi.org/10.2307/1540698
- O’farrell A, Stock A. Regeneration and the Moulting Cycle in. Blattella Germanica L. III. Successive Regeneration of Both Metathoracic Legs. Aust. J. Biol. Sci. 1954;7(4):525-536; PMID:13229848
- Pohley J. Experimentelle Beitrage zur Lenkung der Organentwicklung, des Hautungsrhytmus und der Metamorphose bei der Schabe Periplaneta americana L. Wilhelm Roux; Arch Entwicklungsmech Org. 1959;151:323-80.; http://dx.doi.org/10.1007/BF00577774
- Stock A, O’Farrell AF. Regeneration and the moulting cycle in Blattella germanica L. II. Simultaneous regeneration of both metathoracic legs. Aust. J. Biol. Sci. 1954;7(3):302-7 PMID:13219037
- Atli E, Unlü H. The effects of microwave frequency electromagnetic fields on the development of Drosophila melanogaster. Int. J. Radiat. Biol. 2006;82(6):435-41. PMID:16846978; http://dx.doi.org/10.1080/09553000600798849
- Villee CA. Some effects of x-rays on development in Drosophila. J. Exp. Zool. 1946;101(2):261-280. ; http://dx.doi.org/10.1002/jez.1401010206
- Bourgin RC, Krumins R, Quastler H. Radiation-Induced Delay of Pupation in Drosophila. Radiat. Res. 1956;5(6):657-673; PMID:13379619; http://dx.doi.org/10.2307/3570585
- Madhavan K, Schneiderman H. Hormonal control of imaginal disc regeneration in galleria mellonella (lepidoptera). Biol. Bull. 1969;137(2):321-31; http://dx.doi.org/10.2307/1540104
- Simpson P, Berreur P, Berreur-Bonnenfant J. The initiation of pupariation in Drosophila: dependence on growth of the imaginal discs. J. Embryol. Exp. Morphol. 1980;57:155-65; PMID:7430927
- Stieper BC, Kupershtok M, Driscoll M V, Shingleton AW. Imaginal discs regulate developmental timing in Drosophila melanogaster. Dev. Biol. 2008;321(1):18-26. PMID:18632097.; http://dx.doi.org/10.1016/j.ydbio.2008.05.556
- Poodry CA, Woods DF. Control of the developmental timer for Drosophila pupariation. Roux's Arch. Dev. Biol. 1990;199(4):219-227
- Smith-Bolton RK, Worley MI, Kanda H, Hariharan IK. Regenerative growth in Drosophila imaginal discs is regulated by Wingless and Myc. Dev. Cell 2009;16(6):797-809; PMID:19531351
- Halme A, Cheng M, Hariharan IK. Retinoids regulate a developmental checkpoint for tissue regeneration in Drosophila. Curr. Biol. 2010;20(5):458-63; PMID:20189388; http://dx.doi.org/10.1016/j.cub.2010.01.038
- Hackney JF, Zolali-Meybodi O, Cherbas P. Tissue damage disrupts developmental progression and ecdysteroid biosynthesis in Drosophila. PLoS One 2012;7(11):e49105; PMID:23166607; http://dx.doi.org/10.1371/journal.pone.0049105
- Pohley H. Regeneration and the moulting cycle in Ephestia kuhniella. In: Kiortis V, Trampusch H, eds. Regeneration in Animals. Amsterdam: North Holland Pub. Co.; 1965:324-30
- Simpson P, Schneiderman HA. Isolation of temperature sensitive mutations blocking clone development in Drosophila melanogaster, and the effects of a temperature sensitive cell lethal mutation on pattern formation in imaginal discs. Wilhelm Roux's Arch. Dev. Biol. 1975;178(3):247-275
- Brindley H. On the regeneration of the legs in the Blattidae. Zool Soc Proc 1897:907-16
- O’Farrell AF, Stock A, Rae C, J.A. M. Regeneration and development in the cockroach B. germanica. Acta Soc Entomol Cech 1960;57:317-24
- Villee CA. A Quantitative Study of Phenocopy Production with Monochromatic Ultraviolet Irradiation. Biol. Bull. 1947;92(1):1-9.; http://dx.doi.org/10.2307/1537964
- Knobloch CA, Steel CGH. Interactions between limb regeneration and ecdysteroid titres in last larval instar Rhodnius prolixus (Hemiptera). J. Insect Physiol. 1988;34(6):507-514.; http://dx.doi.org/10.1016/0022-1910(88)90192-8
- Garelli A, Gontijo AM, Miguela V, Caparros E, Dominguez M. Imaginal discs secrete insulin-like peptide 8 to mediate plasticity of growth and maturation. Science 2012;336(6081):579-82; PMID:22556250; http://dx.doi.org/10.1126/science.1216735
- Johnson DG, Walker CL. Cyclins and cell cycle checkpoints. Annu. Rev. Pharmacol. Toxicol. 1999;39:295-312; PMID:10331086; http://dx.doi.org/10.1146/annurev.pharmtox.39.1.295
- Davidowitz G, D’Amico LJ, Nijhout HF. Critical weight in the development of insect body size. Evol. Dev. 2003;5(2):188-97; PMID:12622736; http://dx.doi.org/10.1046/j.1525-142X.2003.03026.x
- Nijhout HF, Williams CM. Control of Moulting and Metamorphosis in the Tobacco Hornworm, Manduca Sexta (L.): Cessation of Juvenile Hormone Secretion as a Trigger for Pupation. J. Exp. Biol. 1974;61(2):493-501
- Mirth C, Truman JW, Riddiford LM. The role of the prothoracic gland in determining critical weight for metamorphosis in Drosophila melanogaster. Curr. Biol. 2005;15(20):1796-807; PMID:16182527; http://dx.doi.org/10.1016/j.cub.2005.09.017
- Andres AJ, Cherbas P. Tissue-specific ecdysone responses: regulation of the Drosophila genes Eip28/29 and Eip40 during larval development. Development 1992;116(4):865-76; PMID:1295740
- Worley MI, Setiawan L, Hariharan IK. Regeneration and transdetermination in Drosophila imaginal discs. Annu. Rev. Genet. 2012;46:289-310; PMID:22934642; http://dx.doi.org/10.1146/annurev-genet-110711-155637
- Gateff E, Schneiderman HA. Developmental capacities of benign and malignant neoplasms of Drosophila. Wilhelm Roux Arch. für Entwicklungsmechanik der Org. 1974;176(1):23-65; http://dx.doi.org/10.1007/BF00577830
- Stewart M, Murphy C, Fristrom JW. The recovery and preliminary characterization of X chromosome mutants affecting imaginal discs of Drosophila melanogaster. Dev. Biol. 1972;27(1):71-83; PMID:4621757; http://dx.doi.org/10.1016/0012-1606(72)90113-3
- Bryant PJ. Regeneration and duplication following operations in situ on the imaginal discs of Drosophila melanogaster. Dev. Biol. 1971;26(4):637-651; PMID:5002603.; http://dx.doi.org/10.1016/0012-1606(71)90146-1
- Woods DF, Bryant PJ. Molecular cloning of the lethal(1)discs large-1 oncogene of Drosophila. Dev. Biol. 1989;134(1):222-35; PMID:2471660; http://dx.doi.org/10.1016/0012-1606(89)90092-4
- Martin P, Martin A, Shearn A. Studies of l(3)c43hs1 a polyphasic, temperature-sensitive mutant of Drosophila melanogaster with a variety of imaginal disc defects. Dev. Biol. 1977;55(2):213-32; PMID:402295; http://dx.doi.org/10.1016/0012-1606(77)90168-3
- Bryant PJ, Schubiger G. Giant and duplicated imaginal discs in a new lethal mutant of Drosophila melanogaster. Dev. Biol. 1971;24(2):233-63; PMID:4994924; http://dx.doi.org/10.1016/0012-1606(71)90097-2
- Rahn P. Untersuchungen zur Entwicklung von Ganz- und Teilimplantaten der Flügelimaginalscheibe vonEphestia kühniella Z. Wilhelm Roux. Arch. Entwickl. Mech. Org. 1972;170(1):48-82; http://dx.doi.org/10.1007/BF00575521
- Dewes E. Regeneration in transplanted halves of male genital disks and its influence upon duration of development in Ephestia kühniella Z. Wilhelm Roux Arch. für Entwicklungsmechanik der Org. 1973;172(4):349-354; http://dx.doi.org/10.1007/BF00577885
- Dewes E. Entwicklungsleistungen implantierter ganzer und halbierter männlicher Genitalimaginalscheiben von Ephestia kühniella Z. und Entwicklungsdauer der Wirtstiere. Wilhelm Roux's Arch. Dev. Biol. 1975;178(2):167-183
- Sehnal F, Bryant PJ. Delayed pupariation in Drosophila imaginal disc overgrowth mutants is associated with reduced ecdysteroid titer. J. Insect Physiol. 1993;39(12):1051-1059; http://dx.doi.org/10.1016/0022-1910(93)90129-F
- Gateff E. Malignant neoplasms of genetic origin in Drosophila melanogaster. Science 1978;200(4349):1448-59; PMID:96525; http://dx.doi.org/10.1126/science.96525
- Kiss I, Molnár I. Metamorphic changes of wild type and mutant Drosophila tissues induced by 20-hydroxy ecdysone in vitro. J. Insect Physiol. 1980;26(6):391-401; http://dx.doi.org/10.1016/0022-1910(80)90010-4
- Yamanaka N, Rewitz KF, O’Connor MB. Ecdysone Control of Developmental Transitions: Lessons from Drosophila Research. 2013 58:497-516; PMID:23072462
- Riddiford LM. Hormone receptors and the regulation of insect metamorphosis. Receptor 1993;3(3):203-9; PMID:8167571
- Buszczak M, Freeman MR, Carlson JR, Bender M, Cooley L, Segraves WA. Ecdysone response genes govern egg chamber development during mid-oogenesis in Drosophila. Development 1999;126(20):4581-9; PMID:10498692
- Carney GE, Bender M. The Drosophila ecdysone receptor (EcR) gene is required maternally for normal oogenesis. Genetics 2000;154(3):1203-11; PMID:10757764
- Chavoshi TM, Moussian B, Uv A. Tissue-autonomous EcR functions are required for concurrent organ morphogenesis in the Drosophila embryo. Mech. Dev. 2010;127(5-6):308-19; PMID:20093179; http://dx.doi.org/10.1016/j.mod.2010.01.003
- Fristrom JW, Logan WR, Murphy C. The synthetic and minimal culture requirements for evagination of imaginal discs of Drosophila melanogaster in vitro. Dev. Biol. 1973;33(2):441-456; PMID:4207978; http://dx.doi.org/10.1016/0012-1606(73)90149-8
- Martin P, Shearn A. Development of Drosophila imaginal discs in vitro: Effects of ecdysone concentration and insulin. J. Exp. Zool. 1980;211(3):291-301.; http://dx.doi.org/10.1002/jez.1402110306
- Postlethwait JH, Schneiderman HA. Induction of metamorphosis by ecdysone analogues. Drosophila imaginal discs cultured in vivo. Biol. Bull. 1970;138(1):47-55; PMID:5437915; http://dx.doi.org/10.2307/1540290
- Guillermet C, Mandaron P. In vitro imaginal disc development and moulting hormone. J Embryol Exp Morphol 1980;57(1):107-118; PMID:6776221
- Mandaron P, Guillerment C, Sengel P. In Vitro Development of Drosophila Imaginal Discs: Hormonal Control and Mechanism of Evagination. Integr. Comp. Biol. 1977;17(3):661-670; http://dx.doi.org/10.1093/icb/17.3.661
- Ashburner M. Sequential gene activation by ecdysone in polytene chromosomes of Drosophila melanogaster. Dev. Biol. 1974;39(1):141-157; PMID:4209831.; http://dx.doi.org/10.1016/S0012-1606(74)80016-3
- Ashburner M, Chihara C, Meltzer P, Richards G. Temporal control of puffing activity in polytene chromosomes. Cold Spring Harb. Symp. Quant. Biol. 1974;38:655-62; http://dx.doi.org/10.1101/SQB.1974.038.01.070
- Boyd M, Ashburner M. The hormonal control of salivary gland secretion in Drosophila melanogaster: Studies in vitro. J. Insect Physiol. 1977;23(4):517-523; http://dx.doi.org/10.1016/0022-1910(77)90263-3
- Kraminsky GP, Clark WC, Estelle MA, Gietz RD, Sage BA, O'Connor JD, Hodgetts RB. Induction of translatable mRNA for dopa decarboxylase in Drosophila: an early response to ecdysterone. Proc. Natl. Acad. Sci. U. S. A. 1980;77(7):4175-9; PMID:6776524; http://dx.doi.org/10.1073/pnas.77.7.4175
- Schwartz LM, Truman JW. Hormonal control of muscle atrophy and degeneration in the moth Antheraea polyphemus. J. Exp. Biol. 1984;111:13-30; PMID:6491588
- Ninov N, Manjón C, Martín-Blanco E. Dynamic control of cell cycle and growth coupling by ecdysone, EGFR, and PI3K signaling in Drosophila histoblasts. PLoS Biol. 2009;7(4):e1000079; PMID:19355788; http://dx.doi.org/10.1371/journal.pbio.1000079
- Smith WA, Gilbert LI, Bollenbacher WE. Calcium-cyclic AMP interactions in prothoracicotropic hormone stimulation of ecdysone synthesis. Mol. Cell. Endocrinol. 1985;39(1):71-8; PMID:2982678; http://dx.doi.org/10.1016/0303-7207(85)90093-0
- Smith WA, Combest WL, Gilbert LI. Involvement of cyclic AMP-dependent protein kinase in prothoracicotropic hormone-stimulated ecdysone synthesis. Mol. Cell. Endocrinol. 1986;47(1-2):25-33; PMID:3017787; http://dx.doi.org/10.1016/0303-7207(86)90012-2
- Gilbert LI, Combest WL, Smith WA, Meller VH, Rountree DB. Neuropeptides, second messengers and insect molting. Bioessays 1988;8(5):153-7; PMID:3044357; http://dx.doi.org/10.1002/bies.950080506
- McBrayer Z, Ono H, Shimell M, Parvy JP, Beckstead RB, Warren JT, Thummel CS, Dauphin-Villemant C, Gilbert LI, O'Connor MB. Prothoracicotropic hormone regulates developmental timing and body size in Drosophila. Dev. Cell 2007;13(6):857-71; PMID:18061567
- Zdarek J, Fraenkel G. Neurosecretory control of ecdysone release during puparium formation of flies. Gen. Comp. Endocrinol. 1971;17(3):483-489; PMID:5128300; http://dx.doi.org/10.1016/0016-6480(71)90183-3
- Roberts B, Gilbert LI, Bollenbacher WE. In vitro activity of dipteran ring glands and activation by the prothoracicotropic hormone. Gen. Comp. Endocrinol. 1984;54(3):469-77; PMID:6735164; http://dx.doi.org/10.1016/0016-6480(84)90164-3
- Harvie PD, Filippova M, Bryant PJ. Genes expressed in the ring gland, the major endocrine organ of Drosophila melanogaster. Genetics 1998;149(1):217-31; PMID:9584098
- Neubueser D, Warren JT, Gilbert LI, Cohen SM. molting defective is required for ecdysone biosynthesis. Dev. Biol. 2005;280(2):362-72; PMID:15882578; http://dx.doi.org/10.1016/j.ydbio.2005.01.023
- Wismar J, Habtemichael N, Warren JT, Dai JD, Gilbert LI, Gateff E. The mutation without children(rgl) causes ecdysteroid deficiency in third-instar larvae of Drosophila melanogaster. Dev. Biol. 2000;226(1):1-17; PMID:10993670; http://dx.doi.org/10.1006/dbio.2000.9811
- Takeuchi H, Chen J, O’Reilly D, Rees H, Turner P. Regulation of ecdysteroid signalling: molecular cloning, characterization and expression of 3-dehydroecdysone 3α-reductase, a novel eukaryotic member of the short-chain dehydrogenases/reductases superfamily from the cotton leafworm, Spodoptera littoralis. 2000;349(Pt 1):239-245; PMID:10861234
- Hadorn E. An Accelerating Effect of Normal “Ring-Glands” on Puparium-Formation in Lethal Larvae of Drosophila Melanogaster. Proc. Natl. Acad. Sci. U. S. A. 1937;23(9):478-84; PMID:16577798; http://dx.doi.org/10.1073/pnas.23.9.478
- Waddington CH, Robertson E. Determination, activation and actinomycin D insensitivity in the optic imaginal disk of Drosophila. Nature 1969;221(5184):933-5; PMID:5765504; http://dx.doi.org/10.1038/221933a0
- Colombani J, Andersen DS, Léopold P. Secreted peptide Dilp8 coordinates Drosophila tissue growth with developmental timing. Science 2012;336(6081):582-5; PMID:22556251; http://dx.doi.org/10.1126/science.1216689
- Shingleton AW, Das J, Vinicius L, Stern DL. The temporal requirements for insulin signaling during development in Drosophila. PLoS Biol. 2005;3(9):e289; PMID:16086608; http://dx.doi.org/10.1371/journal.pbio.0030289
- Böhni R, Riesgo-Escovar J, Oldham S, Brogiolo W, Stocker H, Andruss BF, Beckingham K, Hafen E. Autonomous control of cell and organ size by CHICO, a Drosophila homolog of vertebrate IRS1-4. Cell 1999;97(7):865-75; PMID:10399915; http://dx.doi.org/10.1016/S0092-8674(00)80799-0
- Kramer JM, Davidge JT, Lockyer JM, Staveley BE. Expression of Drosophila FOXO regulates growth and can phenocopy starvation. BMC Dev. Biol. 2003;3:5; PMID:12844367.; http://dx.doi.org/10.1186/1471-213X-3-5
- Shearn A, Rice T, Garen A, Gehring W. Imaginal disc abnormalities in lethal mutants of Drosophila. Proc. Natl. Acad. Sci. U. S. A. 1971;68(10):2594-8; PMID:5002822; http://dx.doi.org/10.1073/pnas.68.10.2594
- Szabad J, Bryant PJ. The mode of action of “discless” mutations in Drosophila melanogaster. Dev. Biol. 1982;93(1):240-256; PMID:6813163.; http://dx.doi.org/10.1016/0012-1606(82)90256-1
- Mensch J, Lavagnino N, Carreira VP, Massaldi A, Hasson E, Fanara JJ. Identifying candidate genes affecting developmental time in Drosophila melanogaster: pervasive pleiotropy and gene–by-environment interaction. BMC Dev. Biol. 2008;8(1):78; PMID:18687152; http://dx.doi.org/10.1186/1471-213X-8-78
- Johnston LA, Prober DA, Edgar BA, Eisenman RN, Gallant P. Drosophila myc regulates cellular growth during development. Cell 1999;98(6):779-90; PMID:10499795; http://dx.doi.org/10.1016/S0092-8674(00)81512-3
- Jacobs HW, Knoblich JA, Lehner CF. Drosophila Cyclin B3 is required for female fertility and is dispensable for mitosis like Cyclin B. Genes Dev. 1998;12(23):3741-51; PMID:9851980; http://dx.doi.org/10.1101/gad.12.23.3741
- Manak JR, Mitiku N, Lipsick JS. Mutation of the Drosophila homologue of the Myb protooncogene causes genomic instability. Proc. Natl. Acad. Sci. U. S. A. 2002;99(11):7438-43; PMID:12032301; http://dx.doi.org/10.1073/pnas.122231599
- Stevens ME, Bryant PJ. Temperature-dependent expression of the apterous phenotype in Drosophila melanogaster. Genetics 1986;112(2):217-28; PMID:3079719
- Shi W, Stampas A, Zapata C, Baker NE. The pineapple eye gene is required for survival of Drosophila imaginal disc cells. Genetics 2003;165(4):1869-79; PMID:14704172
- Dolecek R, Dvorácek C, Jezek M, Kubis M, Sajnar J, Závada M. Very low serum testosterone levels and severe impairment of spermatogenesis in burned male patients. Correlations with basal levels and levels of FSH, LH and PRL after LHRH + TRH. Endocrinol. Exp. 1983;17(1):33-45; PMID:6409583
- Pozo J, Argente J. Delayed puberty in chronic illness. Best Pract. Res. Clin. Endocrinol. Metab. 2002;16(1):73-90; PMID:11987900; http://dx.doi.org/10.1053/beem.2002.0182
- Simon D. Puberty in chronically diseased patients. Horm. Res. 2002;57 Suppl 2:53-6; PMID:12065928; http://dx.doi.org/10.1159/000058102
- Mavoungou D, Poaty-Mavoungou V, Ongali B, Akoume MY, Maka G, Mavoungou E. Hypothalamic-pituitary gonadal axis and immune response imbalance during chronic filarial infections. Trop. Med. Int. Health 2005;10(11):1180-6; PMID:16262744; http://dx.doi.org/10.1111/j.1365-3156.2005.01499.x
- Dolecek R. Endocrine changes after burn trauma–a review. Keio J. Med. 1989;38(3):262-76; PMID:2511373; http://dx.doi.org/10.2302/kjm.38.262
- Emanuele NV, LaPaglia N, Kovacs EJ, Gamelli RL, Emanuele MA. Profound effects of burn and ethanol on proinflammatory cytokines of the reproductive axis in the male mouse. J. Burn Care Res. 29(3):531-40; PMID:18388566; http://dx.doi.org/10.1097/BCR.0b013e3181711273
- Rewitz KF, Rybczynski R, Warren JT, Gilbert LI. The Halloween genes code for cytochrome P450 enzymes mediating synthesis of the insect moulting hormone. Biochem. Soc. Trans. 2006;34(Pt 6):1256-60; PMID:17073797