
ABSTRACT
Shigella flexneri causes severe diarrheal disease worldwide. While many aspects of pathogenesis have been elucidated, significant knowledge gaps remain regarding the role of putative chromosomally-encoded virulence genes. The uncharacterized sap gene encoded on the chromosome has significant nucleotide sequence identity to the fluffy (flu) antigen 43 autotransporter gene in pathogenic Escherichia coli. Here, we constructed a Δsap mutant in S. flexneri strain 2457T and examined the effects of this mutation on bacterial cell aggregation, biofilm formation, and adherence to colonic epithelial cells. Analyses included the use of growth media supplemented with glucose and bile salts to replicate small intestinal signals encountered by S. flexneri. Deletion of the sap gene in 2457T affected epithelial cell adherence, resulted in quicker bacterial cell aggregation, but did not affect biofilm formation. This work highlights a functional role for the sap gene in S. flexneri pathogenesis and further demonstrates the importance of using relevant and appropriate gastrointestinal signals to characterize virulence genes of enteropathogenic bacteria.
Introduction
Shigella is a genus of Gram-negative, facultative anaerobic bacteria that includes four species and more than 50 serotypes. Shigella is an important diarrheal pathogen afflicting the globe, particularly in lower- and middle-income countries.Citation1 The infectious dose is very low, in which as little as 10 to 100 bacteria can cause infection.Citation2 The Global Burden of Disease estimates that Shigella is the second most common cause of diarrheal deaths, with approximately 165,000 annual deaths worldwide, particularly in children, the elderly, and immunocompromised patients.Citation3,Citation4 This disease burden is further complicated by an increasing prevalence of antibiotic resistance and the lack of an effective, approved vaccine.Citation5
The primary mode of transmission of Shigella is through the fecal-oral route typically by ingestion of contaminated food or water.Citation1 After Shigella survives the acidic conditions in the stomach, the bacteria resist bile salts during transit of the small intestine.Citation6,Citation7 Bile salts are the principal components of bile, which aids in digestion by absorbing lipids and fat-soluble vitamins. Moreover, bile salts have antimicrobial properties that compromise the membrane of bacteria.Citation8 However, Shigella and many other enteric pathogens resist bile salts and alter transcriptional profiles in response to exposure.Citation6,Citation7 After reaching the colonic epithelium, Shigella induces transcytosis of M cells, specialized antigen-presenting cells of the colonic epithelium, to invade epithelial cells at the basolateral pole.Citation1 Invasion, cytoskeleton modifications, and host cell signaling manipulation require a series of protein effectors that are delivered into the cytoplasm of the host cell by the needle-like type-III secretion system (T3SS).Citation9 As infection proceeds, interleukin-8 (IL-8) is secreted from infected epithelial cells, resulting in a significant recruitment of neutrophils to the site of infection, ultimately leading to tissue destruction that exacerbates the symptoms of the disease.Citation1,Citation9 Despite extensive research on virulence genes encoded on the Shigella virulence plasmid, the role of several putative virulence genes encoded on the chromosome remains unknown. Characterization of these additional chromosomally-encoded virulence genes may help advance our understanding of infection and illuminate new therapeutic strategies to eradicate shigellosis.
Most S. flexneri serotype 2a strains harbor the she pathogenicity island (PAI), which is a genomic region of 31 open reading frames encoded on the chromosome.Citation10 There is an unusual clustering of three genes that each encode proteins classified as autotransporters, including the putative autotransporter sap (Shigella autotransporter-like protein) gene.Citation10,Citation11 Autotransporters or proteins of the type-V secretion system, are large proteins consisting of an N-terminal signal sequence and a passenger domain that is secreted to the bacterial outer surface through a unique secretion mechanism utilizing the C-terminal β-barrel domain of the protein that embeds into the outer membrane of the bacterial cell.Citation12 The passenger domain is the structural motif that harbors the biological function and is often directly involved in virulence mechanisms such as bacterial cell aggregation, biofilm formation, adhesion to epithelial cells, host immune evasion, and other cytopathic effects.Citation13–17 The sap gene has 99% nucleotide identity to the Escherichia coli flu gene that encodes the autotransporter antigen 43 (Ag43).Citation10,Citation14 Ag43 in E. coli has been characterized to have several functions associated with virulence in different E. coli pathovars, including bacterial cell aggregation and alterations of bacterial shape in enterohemorrhagic Escherichia coli (EHEC) serotype O157:H7.Citation18 Ag43 also contributes to the adhesion of uropathogenic Escherichia coli (UPEC) strain CFT073, which increases long-term persistence of the pathogen in the bladder of a mouse model of urinary tract infection.Citation19 Finally, Ag43 has been shown to enhance the survival of E. coli by forming bacterial aggregates inside neutrophils.Citation20
Despite the unknown function of the sap gene in S. flexneri 2457T serotype 2a, in silico analysis identified a highly immunogenic peptide within the Sap protein, which was recently included in a multi-epitope recombinant fusion vaccine candidate with two additional immunogenic peptides, the autotransporters Pic and SigA from S. flexneri 2457T. Inclusion of the Sap peptide was due to the autotransporter-like properties and the predicted outer membrane localization of the protein.Citation11 Vaccine efficacy was subsequently examined in an intranasal murine infection model and found to increase construct-specific antibody (serum IgG and fecal IgA) and cytokine (TNF-α, IL-17, and IFN-γ) titers. The vaccine candidate conferred 100% protection against S. flexneri 2457T in the murine model, highlighting the importance of mucosal immunity activation, of which the autotransporters appear to be a critical component.Citation21 Since the function of the sap gene in S. flexneri remains largely unknown, the goal of this study was to evaluate a sap mutant to determine a possible role in S. flexneri virulence.
Results
Inspection of the sap coding region with previous RNA-sequencing data and Δsap mutant construction
Since the sap gene (S3195) in S. flexneri serotype 2a strain 2457T (GenBank Accession Number AE014073.1) is currently annotated as a pseudogene, we inspected the sap coding region using previously generated data from high-throughput RNA-sequencing-based gene expression analyses.Citation6 The gene expression data were obtained from S. flexneri 2457T grown in tryptic soy broth (TSB) with and without bile salts under both shaking and static growth conditions at 37°C.Citation6 The analyses of sap gene expression were consistent in the four different growth conditions, and there were no significant differences among the gene expression patterns (). Sequence comparison of the sap gene with the E. coli Ag43 flu gene indicated 99% nucleotide sequence identity.Citation14 However, in S. flexneri 2457T, there is a stop codon in the passenger domain of the sap gene at nucleotide position 894 that also corresponds with a break in the RNA-seq transcript reads (). We further analyzed the sequence region and discovered a putative Rho-dependent termination sequence with the RhoTermPredict algorithm.Citation22 A putative Rho utilization (RUT) site at nucleotides 874–952 and a corresponding pause site with appropriate hairpin structures in the 150 nucleotides immediately downstream of the RUT site were detected,Citation22,Citation23 which could explain the transcriptional profile and break in the RNA-seq reads.
Figure 1. Analyses of the sap coding region and gene deletion strategy. (a) the sap coding region, which is located at the twentieth open reading frame (orf) within the she pathogenicity island, was analyzed using previous RNA-sequencing gene expression data.Citation6 RNA was extracted from S. flexneri 2457T cultures grown in TSB with and without bile salts under both shaking and static growth conditions, each with technical duplicate cultures (A and B). The RNA-seq trace read data were generated using the integrative viewer (IGV) software (version 2.14). There were no significant expression changes in the sap gene in the different growth conditions. The corresponding sap coding region is provided at the bottom of the diagram, and the gap in sequencing corresponds to the annotated stop codon at position 894. (b) Comparison of the sap gene sequence with the Ag43 flu gene sequence of E. coli Citation14 indicated 99% identity of the signal sequence (nucleotides 1–156), the passenger domain (nucleotides 157–1,656) and the β-barrel domain (nucleotides 1,657–3,120). The only sequence difference is the stop codon at position 894 in the S. flexneri 2457T sequence. The lambda red linear recombination method was used to replace the 5’ region of the sap gene, specifically the signal sequence and the passenger domains (1.7 kb), with a kanamycin resistance cassette. The mutation was confirmed by PCR analysis of the wild type and ∆sap kanamycin resistant colonies.
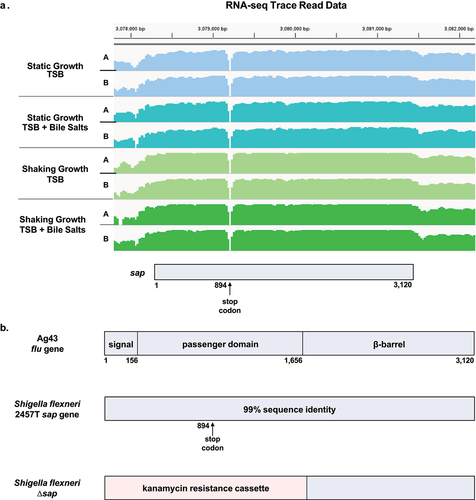
We utilized the stop codon to evaluate sequence divergence of the sap gene across different S. flexneri 2a isolates. Interestingly, the S. flexneri serotype 2a strain YSH6000T sap gene homolog (orf20) does not contain a stop codon (GenBank Accession Number AF200692.2); however, the sap gene (SF2990) in S. flexneri serotype 2a strain 301 (GenBank Accession Number AE005674.2) is annotated as an intact gene from nucleotide 1 to the same premature stop codon, while the C-terminal domain is annotated as a separate outer membrane fluffing protein gene (SF2991). Based on the variable annotations across the S. flexneri serotype 2a strains, we evaluated the sap region in the global collection of S. flexneri isolates analyzed in the GEMS collection ().Citation24 In this global collection of S. flexneri isolates, only 150 (19%) of the 806 sequenced strains encode the sap gene, most of which (17%, 137/806) contain the premature stop codon. An additional 36% of the isolates (291/806) contained a sequence that was significantly similar to the sap gene region but divergent (35–70% nucleotide identity), whereas 45% (365/806) showed little or no primary sequence similarity to the sap gene (<30% nucleotide identity). This in silico analysis highlights the significant sap gene sequence divergence across the S. flexneri species.
Table 1. The presence of the sap gene in the global collection of S. flexneri isolates (GEMS).
To further evaluate the potential functions of the sap gene in the pathogenesis of S. flexneri strain 2457T, we used the lambda red linear recombination methodCitation25 to disrupt the 5’ end of the sap gene, which encodes the N-terminal signal sequence and the passenger domain of the putative Sap protein. This 1.7 kb region was replaced with a kanamycin antibiotic resistance cassette (), which was verified by PCR. The ∆sap mutant was also sequenced to ensure the insertion of the kanamycin cassette was the only significant genetic change to the strain. Subsequent growth curve analyses of the Δsap mutant compared to wild type 2457T in TSB media with and without 0.4% weight/volume (w/v) bile salts indicated no effect of the mutation on the growth of the bacteria or bile salts resistance.
The effect of the deletion of the sap passenger domain on bacterial cell aggregation and biofilm formation in S. flexneri
Bacterial cell aggregation, or autoaggregation, is a phenotype in which bacteria of the same clone or species form multicellular aggregates. This process is typically mediated by self-recognizing proteins that enable bacterial cells to bind together, or through exopolysaccharides such as lipopolysaccharides (LPS) in Gram-negative bacteria.Citation26 Ag43 in E. coli is the most common autotransporter protein involved in this self-recognizing mechanism.Citation14,Citation27 Ag43 is also associated with biofilm formation in some strains of E. coli.Citation14,Citation18 Biofilms are organized bacterial communities that begin with bacterial attachment and microcolony formation followed by the expression of an extracellular polymeric substance (EPS) matrix, which allows bacteria to selectively interact with the extracellular environment and resist harsh conditions.Citation28 Previous literature has not evaluated the ability of S. flexneri to use self-recognizing proteins for autoaggregation. However, cellular aggregation and biofilm formation occur after bile salts and glucose exposure during subculture. The structural proteins of the long polar fimbriae (LpfA), type 1 fimbriae (FimA), and curli (CsgAB) adherence factors, which are also expressed in the presence of both glucose and bile salts, facilitate the adherence stage of biofilm formation in S. flexneri.Citation29 Biofilm formation is further enhanced by robust EPS matrix production starting at approximately 4 h of subculture.Citation6,Citation30 Furthermore, recent work has demonstrated that the S. flexneri autotransporter protein IcsA, a protein that mediates actin-based motility in the cytoplasm of infected host cells,Citation9 facilitates aggregation and bacterial cell-to-cell contact during biofilm formation.Citation31 Importantly, these phenotypes do not occur in Luria broth (LB) media, which is the most common enteric growth media.Citation6 Therefore, and given the function of Ag43 in E. coli, we wanted to evaluate the ∆sap mutant in autoaggregation and biofilm formation assays.
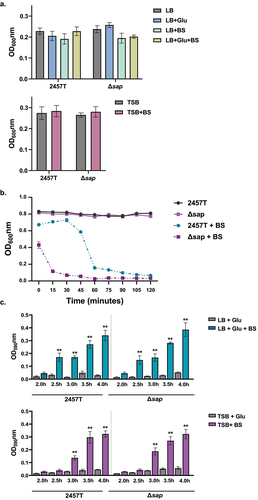
Starting with the self-recognizing mechanism of autoaggregation, this phenotype was examined using a clumping assay that measures bacterial aggregates at the bottom of culture tubes.Citation32 To properly consider autoaggregation both before and after biofilm formation, wild type 2457T and the Δsap mutant were cultured in LB media supplemented with or without glucose and/or bile salts, as well as TSB supplemented with or without bile salts at different time points. Given the range of glucose concentrations in the small intestine,Citation33,Citation34 LB media supplemented with 2% w/v glucose was used to replicate high glucose concentrations, while TSB media already contains added glucose at 0.5% w/v and replicates low glucose concentrations as previously described.Citation6 Under glucose and bile salts subculturing conditions of approximately 2.5 to 3 h (depending on the glucose concentration), EPS matrix production of the biofilm is not yet detected.Citation6 Thus, autoaggregation prior to EPS matrix production was examined in all media types. At this early time point, autoaggregation was not observed for either 2457T or the ∆sap mutant, and there were no significant differences between the strains (). However, after EPS matrix production started, autoaggregation was observed. Interestingly, the ∆sap mutant aggregated quicker relative to 2457T (, note the different decreases in OD600 readings), indicating that autoaggregation during biofilm formation may be independent of or inhibited by the sap gene product. For complete biofilm formation, we compared the abilities of wild type 2457T and the ∆sap mutant to form biofilms during incubation times of 2–4 h using our 96-well plate assay as previously described.Citation6,Citation30 These results demonstrated that biofilm formation progressively increased after bile salts exposure in the presence of glucose, for both 2457T and the ∆sap mutant. There were no significant differences between the strains during the time periods examined (), or longer incubation times up to 24 h (data not shown). These data indicate that the sap gene product is not directly involved in autoaggregation or biofilm formation, including the early stages of formation, in S. flexneri strain 2457T.
Figure 2. Bacterial cell aggregation and biofilm formation analyses. Overnight cultures of S. flexneri wild type strain 2457T and the Δsap mutant were standardized and inoculated into different media formulations at 1:50 dilutions, either LB supplemented with glucose (glu) and/or bile salts (BS) or TSB supplemented with bile salts, and subcultured. All data are plotted as the average of three biological independent experiments, ± the standard error of the mean. (a) To measure bacterial cell aggregation prior to EPS matrix formation, subcultures were grown in shaking conditions until growth reached an OD600 of 0.7. Bacterial cultures were then centrifuged, resuspended in 2 ml of 1X PBS, and placed on the benchtop for 18 h. The OD600 was measured 0.5 cm from the top of the supernatants at the beginning (time 0) and after the 18 h incubation. Bacterial cell aggregation was measured as the final OD600 readings at 18 h subtracted from the initial OD600 readings for each condition. No significant differences were detected in any of the conditions tested. (b) To measure bacterial cell aggregation after EPS matrix formation, subcultures were grown as noted above for 4 hours in TSB with and without bile salts supplementation. While there was no autoaggregation in 2457T or the Δsap mutant in TSB, both strains aggregated in TSB + BS (p < 0.01 for all time points), and the Δsap mutant aggregated quicker compared to 2457T (p ≤0.001 for all time points). Analyses were performed in LB + glu ± BS and the same results were obtained, but the higher glucose concentrations resulted in the Δsap mutant aggregating within five minutes (data not shown). (c) Biofilm formation was monitored every 30 minutes between hours 2 to 4 of incubation to capture the early phase of formation. There were significant increases in the biofilm formation in all media containing both glucose and bile salts in 2457T and the Δsap mutant (p < 0.005) as indicated by the asterisks. EPS matrix production started at 2.5 hours for LB + glu + BS and at 3.0 hours for TSB + BS in which the delay is due to the lower glucose concentration. There were no significant differences between 2457T and the Δsap mutant.
The sap gene product has a role in S. flexneri adherence to HT-29 intestinal epithelial cells
Colonization of the gastrointestinal tract by commensal or pathogenic bacteria is typically mediated by surface structures such as pili or fimbriae to initiate contact with host cells.Citation35 Ag43 has been shown to facilitate adherence of E. coli.Citation19 S. flexneri adherence to colonic epithelial cells is mediated by surface proteins, including LpfA, FimA, and CsgAB, as well as the T3SS effectors OspE1 and OspE2, in which adherence and surface localization of these proteins are induced after exposure to physiological concentrations of bile salts and glucose.Citation6,Citation29,Citation36 To determine a possible role of the sap gene product in the adherence of S. flexneri to colonic epithelial cells, adherence assays with HT-29 cells were performed for wild type 2457T and the Δsap mutant. The adherence of both bacterial strains was evaluated following bacterial subculturing in high (LB + glucose) and low (TSB) glucose concentrations, both with or without bile salts. It is essential to note that glucose and bile salts are important small intestinal signals. For subsequent infection analyses of colonic epithelial cells, the bacteria were washed and resuspended in standard tissue culture media to mimic the transition into the colon prior to applying the bacteria to the HT-29 cells.Citation6,Citation29,Citation36 Our results demonstrate that wild type 2457T and the Δsap mutant both displayed increased adherence after bile salts exposure in both media types relative to media without bile salts (p < 0.005; ). Interestingly, despite the same pattern of induced adherence following bile salts exposure, the adherence rate of the Δsap mutant relative to wild type 2457T was significantly lower, by approximately 50%, only when the bacteria were subcultured in bile salts (p < 0.005; ). These data indicate that the sap gene product has a role in the adherence phenotype of S. flexneri to HT-29 cells, which was detected only after subculturing the bacteria in the presence of small intestinal signals.
Figure 3. Analysis of the ∆sap mutant in adherence to and invasion of HT-29 epithelial cells. Infection analyses were performed following bacterial subculture in LB media supplemented with glucose (glu), with or without bile salts (BS), as well as TSB media with or without bile salts. The average percent recoveries ± standard error of the mean from three biological replicates are plotted relative to the recovery rate for 2457T from the bile salts subcultures (set at 100%). (a) In adherence assays following subculture in either LB + glu ± BS or TSB ± BS, the adherence rates were induced for both wild type 2457T and the Δsap mutant following bile salts exposure (**, p < 0.005). However, the Δsap mutant had lower adherence following subculture in bile salts compared to 2457T in the same condition (ǂ, p < 0.005). There were no significant differences in adherence between 2457T or the Δsap mutant following subculturing in media without bile salts. While there is a decrease in adherence in the Δsap mutant relative to 2457T following TSB subculture without bile salts, the difference was not statistically significant. (b) For invasion, there were no significant differences between 2457T and the Δsap mutant following subculture in LB + glu ± BS. Both strains had significant increases in invasion following exposure to bile salts (**, p < 0.005).
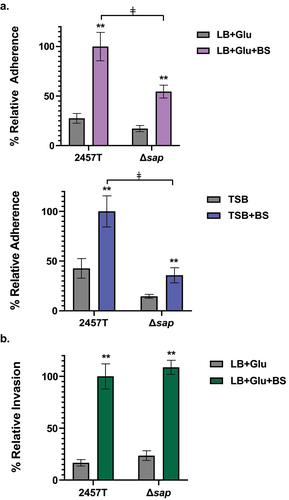
The invasion of colonic epithelial cells is a critical step in the S. flexneri lifecycle, in which entry into the cellular cytoplasm is required to establish a replicative niche of S. flexneri. Given the importance of host cell invasion, we evaluated the ability of the Δsap mutant to invade the epithelial cells. For Ag43, there is no evidence that this autotransporter plays a role in invasion,Citation37 especially since most E. coli pathovar isolates are noninvasive.Citation38 Thus, we hypothesized that the mutation would not affect invasion of S. flexneri. To test this hypothesis, we performed an invasion assay with HT-29 cells following the same subculture conditions as the adherence assays.Citation6,Citation39 The bacteria were centrifuged onto the HT-29 cells to facilitate bacterial contact with the epithelial cells and bypass the adherence step.Citation39 As expected, bile salts exposure increased the invasion ratesCitation40,Citation41 for both wild type 2457T and the ∆sap mutant, but there were no statistical differences (), indicating that the sap gene is not directly involved in the invasion of S. flexneri into colonic epithelial cells.
Discussion
Bacillary dysentery caused by S. flexneri is a bloody diarrheal disease that is the second leading cause of mortality.Citation3 Despite understanding important concepts in pathogenesis, particularly regarding host cell invasion and immune evasion, many gaps in knowledge remain. The chromosomal she PAI of S. flexneri harbors autotransporter genes such as sap that encode highly immunogenic proteins, particularly the amino acid sequence of the passenger domain.Citation11 Given the immunogenic effect of a recombinant protein that included a Sap peptide in a S. flexneri vaccine candidate,Citation21 we sought to study the role of the sap gene in S. flexneri pathogenesis. We evaluated the ∆sap mutant in autoaggregation, biofilm formation, and adherence assays due to the homology of sap to the flu gene encoding the autotransporter Ag43.Citation10,Citation14–16,Citation19 Furthermore, since many of these phenotypes require supplemented laboratory media for the appropriate analysis of S. flexneri,Citation6,Citation29 we performed our analyses with standard laboratory media, as well as media supplemented with bile salts and glucose based on our previous work.
Autoaggregation is defined as multicellular clumps of bacteria that confer protection against environmental stresses or host responses, and can also help initiate biofilm formation in a self-recognizing manner.Citation26 Thus, it is important to evaluate both autoaggregation and biofilm formation in the ∆sap mutant. Our analyses demonstrated that the S. flexneri sap gene product is not directly involved in either process. We tested for autoaggregation at 2 h of subculturing, which is prior to when the EPS matrix is secreted during biofilm formation (see the increases in OD590 readings in at 2.5 and 3 h, depending on the glucose concentration and our previous work.Citation6,Citation29 In experiments in which autoaggregation was tested after secretion of the EPS matrix, the ∆sap mutant aggregated quicker than wild type 2457T (). Given that sap gene expression was not affected by bile salts exposure (), we were not surprised by the lack of a direct effect of the sap deletion on autoaggregation or biofilm formation, but we were surprised by the quicker aggregative phenotype of the Δsap mutant following EPS secretion. A recent study demonstrated that the S. flexneri autotransporter icsA gene product can mediate bacterial cell-to-cell contact and aggregative growth in the presence of bile salts, which facilitates biofilm formation.Citation31 Interestingly, the expression of icsA was not altered by bile salts exposure,Citation6 suggesting that constitutive expression of icsA is sufficient to facilitate biofilm formation, possibly by interacting with other proteins or the EPS matrix.Citation6,Citation29 We hypothesize Sap either inhibits autoaggregation or that the absence of Sap in these conditions enables more efficient surface localization of other proteins such as IcsA, the adherence factors LpfA, FimA, and CsgAB, and/or EPS matrix components to aid autoaggregation during biofilm formation. In fact, it has been documented that type 1 fimbriae can block Ag43-mediated autoaggregation.Citation14,Citation42 Future peptide-specific analyses will verify these results, which should consider potential phase variation of the sap gene under different conditions given the homology to the Ag43 flu gene.Citation14,Citation43 Nevertheless, our data demonstrated that the sap gene product in S. flexneri 2457T is not directly involved in autoaggregation or biofilm formation under the conditions examined.
Despite the lack of an effect of the ∆sap mutation on autoaggregation and biofilm formation, the mutation affected adherence to HT-29 epithelial cells. Adherence enables pathogens to directly contact and colonize the host. Previous work by our laboratory has identified several gene products that directly facilitate S. flexneri adherence to colonic epithelial cells: the virulence plasmid-associated homologs ospE1 and ospE2, as well as the chromosomally encoded lpfA, fimA and csgAB genes.Citation6,Citation29,Citation44 The analyses here indicate that the sap gene product also facilitates S. flexneri adherence, which was not fully detected until the bacteria were subcultured with gastrointestinal signals encountered during small intestinal transit, which were provided prior to evaluating the adherence function to colonic epithelial cells. It is important to emphasize that the bile salts and glucose supplementation occur prior to adding the bacteria to the epithelial cells as previously discussed.Citation6,Citation29,Citation45 Interestingly, the ∆sap mutant had the same induced adherence pattern as wild type 2457T after bile salts and glucose exposure, but the overall adherence rate was reduced relative to 2457T. Therefore, we hypothesize that the adherence function of the putative Sap protein is independent of the OspE1, OspE2, LpfA, FimA, and CsgAB proteins, which are affected by bile salts and glucose exposure.Citation29,Citation44 The results are further validated by the constitutive expression of the sap gene as noted in our RNA-seq analyses. Furthermore, the reduced adherence of the ∆sap mutant relative to 2457T provides additional evidence that the putative Sap protein is expressed on the bacterial surface, which supports the immunogenic nature of the proteinCitation11,Citation21 and the increased rate of autoaggregation of the ∆sap mutant after EPS matrix secretion (). Future protein-specific analyses are needed to determine how the putative Sap protein is secreted and localized to the bacterial outer surface to function as an adherence factor, especially given the presence of the stop codon that potentially disrupts the association of the passenger domain with the C-terminal β-barrel domain. Finally, the ∆sap mutation did not affect the invasion ability of S. flexneri, suggesting that the virulence role of Sap is limited to epithelial cell adherence.
Sequence analysis of the sap gene across a collection of S. flexneri isolates identified a similar stop codon at nucleotide position 894 in the passenger domain of sap that was identified in S. flexneri serotype 2a strain 2457T. This stop codon is also present in the reference S. flexneri serotype 2a strain 301, but not present in S. flexneri serotype 2a strain YSH6000T. The stop codon in strain 301 resulted in a two gene annotation of the sap gene (SF2990) and SF2991 described as an outer membrane fluffing protein. Additionally, upon inspection of the GEMS collection of S. flexneri isolates, most strains either did not possess any sequence identity to the sap gene region or had significant sequence divergence that may result in functional variation. Of the isolates that contain the sap gene region, which may be specific to serotype 2a, the stop codon is in a similar location, suggesting that this annotation is a common feature of the sap gene in S. flexneri. Future analyses of the effects of this sap sequence divergence and genetic variability could identify effects on virulence-associated functions. Furthermore, sequence divergence could provide some protection from the host immune response.Citation20,Citation26 For Ag43, sequence and/or functional variability is well documented among the E. coli species, including functional diversity across different pathovars,Citation27 two identical alleles in EHEC O157:H7 strain EDL933,Citation18 altered functional diversity of two alleles in UPEC strain CFT073,Citation19 and other variations such as Dr adhesin-mediated inhibition of Ag43 in UPEC strain IH11128.Citation16 Thus, the sequence variability found in the S. flexneri sap gene, including the stop codon in the passenger domain of 2457T and other strains, likely affects the function of Sap across the Shigella species and serotypes. This new finding of virulence function related to the sap gene of 2457T offers opportunities to study the effects of sap sequence divergence on S. flexneri virulence.
In conclusion, this study functionally characterizes the S. flexneri 2457T sap gene. Despite the nucleotide sequence homology to the E. coli Ag43 flu gene, the pseudogene annotation in 2457T, and the lack of direct function associated with autoaggregation and biofilm formation, we demonstrated that the sap gene facilitates S. flexneri adherence to colonic epithelial cells. Furthermore, this study highlights the importance of using relevant and appropriate gastrointestinal signals to study the pathogenesis of enteropathogenic bacteria and related host–pathogen interactions. Future areas of study include protein-specific analyses and understanding of the effects of sap gene sequence divergence. This work improves our understanding of S. flexneri adherence, further demonstrates the importance of confirming proper pseudogene annotations, and establishes a new avenue of investigation to improve our understanding of Shigella pathogenesis and ultimately develop novel therapeutic candidates.
Materials and methods
Bacterial strains and growth conditions
The bacterial strains, wild type S. flexneri serotype 2a strain 2457T and 2457T/sap:aph-3 (Δsap, kanamycin resistance), were cultured at 37°C in LB (Lennox) or TSB. LB supplemented with glucose was added at a concentration of 2% w/v, and TSB contains approximately 0.5% w/v glucose relative to LB. Bile salts (catalog number B8756; Sigma-Aldrich) were used at a concentration of 0.4% w/v. Bacterial strains were routinely maintained on plates containing TSB with 1.5% agar and 0.025% Congo red (CR; catalog number C6277; Sigma-Aldrich). All media were autoclaved or filter-sterilized with a 0.22 µm membrane following the addition of bile salts and/or glucose. Kanamycin was used at a concentration of 50 µg/ml, as indicated.
Prevalence of the sap gene in the GEMS collection
RNA-seq data and mapping were obtained as previously described,Citation6 with read data visualized using Integrative Viewer (IGV) software (version 2.14).Citation46,Citation47 For the GEMS isolates, 806 S. flexneri isolates were sequenced and assembled as previously described.Citation24 The sap gene from S. flexneri 2457T (GenBank Accession number NC_004741.1) was searched against the assembled genomes using BLASTN. The sap gene was considered significantly similar if it was > 95% nucleotide identical to > 70% of the complete sap gene region. Variable gene presence was defined as a BLASTN hit that was < 10−100 and > 95% nucleotide identity over at least 30% of the sap gene region. Sequences without significant similarities were considered absent.
Construction of the Δsap mutant targeting the passenger domain of the gene
Deletion of the 5’ signal sequence and passenger domains of the sap gene was performed by allelic exchange using the lambda red linear recombination method.Citation25 Briefly, the plasmid pKD4 was used as a template for the amplification of the kanamycin resistance cassette (aph-3) via PCR using primers sap_kanF (5’-ATGAAACGACATCTGAATACCTGCTACAGGCTGGTATGGAATCACATTACGTGTAGGCTGGAGCTGCTTC-3’) and sap_kanR (5’-GTTATCGGGAATATTCCAGGTGGCACCAGAGGCGAGAGTG ACATTCGTGGCATATGAATACCTCCTTAG-3’). Each primer consisted of a 50-nucleotide sequence homologous to the specified region of sap, followed by 20 nucleotides homologous to aph-3 at the 3’ end. The PCR product was then used to transform S. flexneri 2457T harboring plasmid pKM208, which contains the λ-red gam, beta, and exo genes, to facilitate the homologous recombination of the linear PCR product into the sap region. Positive transformant colonies were selected by kanamycin resistance and confirmed via PCR using primers sap_conF (5’-CCTGCCGGTATCCACATCTG-3’) and sap_conR (5’-TGCCATATCCGGGCGTACAC-3’), which amplified the 5’ upstream region of sap and into the kanamycin cassette to generate a 1.3 kb product to confirm presence of the mutation. Wild type 2457T was used as the negative control for the PCR. Finally, to confirm the ∆sap mutation was the only significant sequence alteration in the ∆sap mutant strain, total genomic DNA was extracted and sequenced at SeqCenter, Pittsburgh, PA. The sap mutant was evaluated using BreSeqCitation48 with default settings with Illumina generated 2 × 151bp paired-end read data as the input for variant calling against the S. flexneri 2457T reference genome (CP100044-CP100048).
Analysis of bacterial cell aggregation
To analyze the possible role of the sap gene product in bacterial cell aggregation, or autoaggregation, the clumping assay was performed as previously described with minor modifications.Citation32 Briefly, overnight cultures of wild type 2457T and the Δsap mutant were standardized and subcultured 1:50 into either LB with 2% w/v glucose or TSB media, each with or without 0.4% w/v bile salts. Cultures were grown at 37°C at 225 rpm for approximately 2.0 to 2.5 h to an OD600 of 0.7. For analysis of autoaggregation after biofilm formation, cultures were grown for 4 h, and the OD600 values were measured. Bacterial subcultures were then centrifuged at 4000 × g, washed, resuspended in 2 ml of 1X PBS, and standardized to an OD600 of 1.0 in polyethylene tubes. The tubes were then placed on a benchtop at room temperature for up to 18 h. The OD600 of the culture supernatants was measured by taking 100 µl at a distance of 0.5 cm below the surface of the culture. Measurements were taken at the initial placement of the culture tubes on the benchtop (time 0) and at each time point. The supernatants were placed in a 96-well plate for absorbance readings using a SpectraMax® plate reader. The final cellular aggregation was measured by subtracting the OD600 at time 0 from the OD600 at the end of the assay.
Analysis of biofilm formation
To evaluate the role of the sap gene product in biofilm formation, a biofilm assay was performed in both LB supplemented with glucose and TSB, both with and without bile salts, as previously described.Citation6,Citation30 Briefly, overnight cultures of wild type 2457T and the Δsap mutant were standardized and 20 µL of each culture was added to 1 ml of each media formulation (1:50 dilution). Next, 130 µL of the inoculated culture was added to three wells (for technical triplicate readings) of a flat-bottomed, tissue culture-treated 96-well plate (Millipore Sigma, USA). Plates were incubated at 37°C, and biofilm formation was measured every 30 min between 2 and 4 h of incubation. Afterward, the media were carefully aspirated, and the plates were washed with 1X PBS and left to dry. The wells were then stained with 0.5% crystal violet for 5 min, washed five times with distilled water, and incubated with 95% ethanol for 30 min at room temperature. The absorbance was measured at an optical density of 590 nm (OD590) using a SpectraMax® plate reader. Samples were blanked to wells that were incubated with media and treated using the same procedure.
HT-29 adherence and invasion assays
The role of the sap gene product in the adherence and invasion of S. flexneri was evaluated using the human colorectal adenocarcinoma cell line HT-29 (ATCC HTB-38) as previously described.Citation6 Briefly, HT-29 cells were cultured in 6-well plates using DMEM with 10% heat-inactivated fetal bovine serum (Thermo Fisher Scientific, Waltham, MA, USA) at 37°C under 5% CO2 until reaching approximately 85% confluency. For the adherence assay, overnight cultures of wild type 2457T and the Δsap mutant were diluted 1:50 and subcultured in LB supplemented with 2% w/v glucose or TSB media, both with or without 0.4% w/v bile salts. Subcultures were grown at 37°C with shaking at 225 rpm for approximately 2 h until the cultures reached an OD600 of 0.7. The subcultures were then standardized to an OD600 of 0.35, centrifuged, washed with 1X PBS, and resuspended in 1 ml of DMEM for application onto HT-29 cells. Prior to the application of the bacterial cultures, HT-29 cells were washed in 1X PBS and resuspended in DMEM without FBS. For application of the bacteria onto the cells, the DMEM was removed, and the standardized bacterial samples resuspended in 1 ml of DMEM were added to the cells. The cells were incubated at 37°C with 5% CO2 for 3 h. Afterward, monolayers were washed five times with 1X PBS and the cells were subsequently lysed with 1% Triton X-100. The number of adherent bacteria was determined by serial dilution and colony counting on TSB agar plates containing CR. The adherence rate was calculated as [recovered bacterial titer/infecting bacterial titer] × 100%.
For the invasion assay, bacteria were prepared as described above. After applying the bacteria onto HT-29 cells, an additional 1 ml of DMEM was added to each well, and the plates were centrifuged at 3000 rpm for 10 min to facilitate bacterial contact with the cells. Afterward, the plates were incubated for 45 min at 37°C with 5% CO2. The HT-29 cells were then washed five times with 1X PBS and incubated with DMEM plus 50 µg/ml gentamicin at 37°C with 5% CO2 for 30 min. The cells were washed again with 1X PBS and incubated with fresh DMEM plus gentamicin for 60 min to ensure lysis of extracellular bacteria. After the incubation, HT-29 cells were washed three times with 1X PBS, and the cells were lysed with 1% Triton X-100. The number of invading bacteria was determined by serial dilution and colony counting on TSB agar plates containing CR. The invasion rate was calculated as [recovery bacterial titer/infecting bacterial titer] × 100%.
Statistical analyses
All experiments were performed with three biological replicates, with either three technical replicates for the biofilm formation assay or two technical replicates for the autoaggregation and infection (adherence and invasion) assays. Data for the infection assays were plotted relative to 2457T from the LB + 2% glucose + 0.4% bile salts or TSB + 0.4% bile salts subcultures, which were set at 100%. Two-way analysis of variance (ANOVA) was used to evaluate differences between treatments and/or wild type 2457T and Δsap mutant in the various assays. Graphs and statistical comparisons were generated using GraphPad Prism 5.04. Statistical significance was set at p value of < 0.05.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Data availability statement
The authors confirm that data supporting the findings of this study are available within the article or upon request from the corresponding author.
Additional information
Funding
References
- Schnupf P, Sansonetti PJ, Cossart P, Roy CR, Sansonetti P. Shigella pathogenesis: new insights through advanced methodologies. Microbiol Spectr. 2019;7(2). doi:10.1128/microbiolspec.BAI-0023-2019.
- Porter CK, Thura N, Ranallo RT, Riddle MS. The Shigella human challenge model. Epidemiol Infect. 2013;141(2):223–14. doi:10.1017/S0950268812001677.
- Khalil IA, Troeger C, Blacker BF, Rao PC, Brown A, Atherly DE, Brewer TG, Engmann CM, Houpt ER, Kang G. et al. Morbidity and mortality due to shigella and enterotoxigenic Escherichia coli diarrhoea: the global burden of disease study 1990–2016. Lancet Infect Dis. 2018;18(11):1229–1240. doi:10.1016/S1473-3099(18)30475-4.
- Troeger C, Forouzanfar M, Rao PC, Khalil I, Brown A, Reiner RC, Fullman N, Thompson RL, Abajobir A, Ahmed M. et al. Estimates of global, regional, and national morbidity, mortality, and aetiologies of diarrhoeal diseases: a systematic analysis for the global burden of disease study 2015. The Lancet Infectious Diseases. 2017;17(9):909–948. doi:10.1016/S1473-3099(17)30276-1.
- Ranjbar R, Farahani A. Shigella: antibiotic-resistance mechanisms and new horizons for treatment. Infect Drug Resist. 2019;12:3137–3167. doi:10.2147/IDR.S219755.
- Nickerson KP, Chanin RB, Sistrunk JR, Rasko DA, Fink PJ, Barry EM, Nataro JP, Faherty CS. Analysis of shigella flexneri resistance, biofilm formation, and transcriptional profile in response to bile salts. Infect Immun. 2017;85(6). doi:10.1128/IAI.01067-16.
- Sistrunk JR, Nickerson KP, Chanin RB, Rasko DA, Faherty CS. Survival of the fittest: how bacterial pathogens utilize bile to enhance infection. Clin Microbiol Rev. 2016;29(4):819–36. doi:10.1128/CMR.00031-16.
- Ridlon JM, Kang DJ, Hylemon PB. Bile salt biotransformations by human intestinal bacteria. J Lipid Res. 2006;47(2):241–59. doi:10.1194/jlr.R500013-JLR200.
- Mattock E, Blocker AJ. How do the virulence factors of shigella work together to cause disease? Front Cell Infect Microbiol. 2017;7:64. doi:10.3389/fcimb.2017.00064.
- Al-Hasani K, Rajakumar K, Bulach D, Robins-Browne R, Adler B, Sakellaris H. Genetic organization of the she pathogenicity island in shigella flexneri 2a. Microb Pathog. 2001;30(1):1–8. doi:10.1006/mpat.2000.0404.
- Leon Y, Zapata L, Salas-Burgos A, Onate A. In silico design of a vaccine candidate based on autotransporters and HSP against the causal agent of shigellosis, shigella flexneri. Mol Immunol. 2020;121:47–58. doi:10.1016/j.molimm.2020.02.008.
- Dautin N. Serine protease autotransporters of enterobacteriaceae (SPATEs): biogenesis and function. Toxins (Basel). 2010;2(6):1179–206. doi:10.3390/toxins2061179.
- Al-Hasani K, Henderson IR, Sakellaris H, Rajakumar K, Grant T, Nataro JP, Robins-Browne R, Adler B. The sigA gene which is borne on the she pathogenicity island of shigella flexneri 2a encodes an exported cytopathic protease involved in intestinal fluid accumulation. Infect Immun. 2000;68(5):2457–2463. doi:10.1128/IAI.68.5.2457-2463.2000.
- van der Woude MW, Henderson IR, van der Woude MW. Regulation and function of Ag43 (flu). Annu Rev Microbiol. 2008;62(1):153–169. doi:10.1146/annurev.micro.62.081307.162938.
- Mendez-Arancibia E, Vargas M, Soto S, Ruiz J, Kahigwa E, Schellenberg D, Gascón J, Urassa H, Schellenberg D. Prevalence of different virulence factors and biofilm production in enteroaggregative Escherichia coli isolates causing diarrhea in children in Ifakara (Tanzania). Am J Trop Med Hyg. 2008;78(6):985–989. doi:10.4269/ajtmh.2008.78.985.
- Zalewska-Pia Tek B, Pia Tek R, Olszewski M, Kur J. Identification of antigen Ag43 in uropathogenic Escherichia coli Dr+ strains and defining its role in the pathogenesis of urinary tract infections. Microbiol. 2015;161(5):1034–1049. doi:10.1099/mic.0.000072.
- Henderson IR, Nataro JP, Portnoy DA. Virulence functions of autotransporter proteins. Infect Immun. 2001;69(3):1231–43. doi:10.1128/IAI.69.3.1231-1243.2001.
- Torres AG, Perna NT, Burland V, Ruknudin A, Blattner FR, Kaper JB. Characterization of Cah, a calcium-binding and heat-extractable autotransporter protein of enterohaemorrhagic Escherichia coli. Mol Microbiol. 2002;45(4):951–66. doi:10.1046/j.1365-2958.2002.03094.x.
- Ulett GC, Valle J, Beloin C, Sherlock O, Ghigo JM, Schembri MA. Functional analysis of antigen 43 in uropathogenic Escherichia coli reveals a role in long-term persistence in the urinary tract. Infect Immun. 2007;75(7):3233–44. doi:10.1128/IAI.01952-06.
- Fexby S, Bjarnsholt T, Jensen PO, Roos V, Hoiby N, Givskov M, Klemm P. Biological Trojan horse: antigen 43 provides specific bacterial uptake and survival in human neutrophils. Infect Immun. 2007;75(1):30–34. doi:10.1128/IAI.01117-06.
- Leon Y, Zapata L, Molina RE, Okanovic G, Gomez LA, Daza-Castro C, Flores-Concha M, Reyes JL, Oñate AA. Intranasal immunization of mice with multiepitope chimeric vaccine candidate based on conserved autotransporters SigA, pic and Sap, confers protection against shigella flexneri. Vaccines (Basel). 2020;8(4):563. doi:10.3390/vaccines8040563.
- Di Salvo M, Puccio S, Peano C, Lacour S, Alifano P. RhoTermPredict: an algorithm for predicting Rho-dependent transcription terminators based on Escherichia coli, Bacillus subtilis and Salmonella enterica databases. BMC Bioinf. 2019;20(1):117. doi:10.1186/s12859-019-2704-x.
- Banerjee S, Chalissery J, Bandey I, Sen R. Rho-dependent transcription termination: more questions than answers. J Microbiol. 2006;44:11–22.
- Bengtsson RJ, Simpkin AJ, Pulford CV, Low R, Rasko DA, Rigden DJ, Hall N, Barry EM, Tennant SM, Baker KS. et al. Pathogenomic analyses of shigella isolates inform factors limiting shigellosis prevention and control across LMICs. Nat Microbiol. 2022;7(2):251–61. doi:10.1038/s41564-021-01054-z.
- Datsenko KA, Wanner BL. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc Natl Acad Sci U S A. 2000;97(12):6640–5. doi:10.1073/pnas.120163297.
- Trunk T, Khalil HS, Leo JC. Bacterial autoaggregation. AIMS Microbiol. 2018;4(1):140–64. doi:10.3934/microbiol.2018.1.140.
- Ageorges V, Schiavone M, Jubelin G, Caccia N, Ruiz P, Chafsey I, Bailly X, Dague E, Leroy S, Paxman J. et al. Differential homotypic and heterotypic interactions of antigen 43 (Ag43) variants in autotransporter-mediated bacterial autoaggregation. Sci Rep. 2019;9(1):11100. doi:10.1038/s41598-019-47608-4.
- Gupta P, Sarkar S, Das B, Bhattacharjee S, Tribedi P. Biofilm, pathogenesis and prevention—a journey to break the wall: a review. Arch Microbiol. 2015;198(1):1–15. doi:10.1007/s00203-015-1148-6.
- Chanin RB, Nickerson KP, Llanos-Chea A, Sistrunk JR, Rasko DA, Kumar DKV, de la Parra J, Auclair JR, Ding J, Li K. et al. Shigella flexneri adherence factor expression in In vivo-like conditions. mSphere. 2019;4:e00751–19. doi:10.1128/mSphere.00751-19.
- Nickerson KP, Faherty CS. Bile Salt-induced Biofilm Formation in Enteric Pathogens: Techniques for Identification and Quantification. J Vis Exp. 2018;135. Forthcoming.10.3791/57322.
- Koseoglu VK, Hall CP, Rodriguez-Lopez EM, Agaisse H, Roy CR. The autotransporter IcsA promotes shigella flexneri biofilm formation in the presence of bile salts. Infect Immun. 2019;87(7). doi:10.1128/IAI.00861-18.
- Xu D, Zhang W, Zhang B, Liao C, Shao Y. Characterization of a biofilm-forming Shigella flexneri phenotype due to deficiency in hep biosynthesis. PeerJ. 2016;4:e2178. doi:10.7717/peerj.2178.
- Kamada N, Chen GY, Inohara N, Nunez G. Control of pathogens and pathobionts by the gut microbiota. Nat Immunol. 2013;14(7):685–90. doi:10.1038/ni.2608.
- Thomson AB, Keelan M, Thiesen A, Clandinin MT, Ropeleski M, Wild GE. Small bowel review: normal physiology part 1. Dig Dis Sci. 2001;46(12):2567–87. doi:10.1023/A:1012794505897.
- Kline KA, Dodson KW, Caparon MG, Hultgren SJ. A tale of two pili: assembly and function of pili in bacteria. Trends Microbiol. 2010;18(5):224–32. doi:10.1016/j.tim.2010.03.002.
- Faherty CS, Harper JM, Shea-Donohue T, Barry EM, Kaper JB, Fasano A, Nataro JP. Chromosomal and plasmid-encoded factors of shigella flexneri induce secretogenic activity ex vivo. PloS One. 2012;7(11):e49980. doi:10.1371/journal.pone.0049980.
- de Luna M, Scott-Tucker A, Desvaux M, Ferguson P, Morin NP, Dudley EG, de Luna MDG, Turner S, Nataro JP, Owen P. et al. The Escherichia coli biofilm-promoting protein Antigen 43 does not contribute to intestinal colonization. FEMS Microbiol Lett. 2008;284(2):237–246. doi:10.1111/j.1574-6968.2008.01207.x.
- Kaper JB, Nataro JP, Mobley HL. Pathogenic Escherichia coli. Nat Rev Microbiol. 2004;2(2):123–40. doi:10.1038/nrmicro818.
- Emanuele AA, Adams NE, Chen YC, Maurelli AT, Garcia GA. Potential novel antibiotics from HTS targeting the virulence-regulating transcription factor, VirF, from Shigella flexneri. J Antibiot (Tokyo). 2014;67(5):379–86. doi:10.1038/ja.2014.10.
- Pope LM, Reed KE, Payne SM. Increased protein secretion and adherence to HeLa cells by Shigella spp. following growth in the presence of bile salts. Infect Immun. 1995;63(9):3642–8. doi:10.1128/iai.63.9.3642-3648.1995.
- Olive AJ, Kenjale R, Espina M, Moore DS, Picking WL, Picking WD. Bile salts stimulate recruitment of IpaB to the Shigella flexneri surface, where it colocalizes with IpaD at the tip of the type III secretion needle. Infect Immun. 2007;75(5):2626–9. doi:10.1128/IAI.01599-06.
- Hasman H, Chakraborty T, Klemm P. Antigen-43-mediated autoaggregation of Escherichia coli is blocked by fimbriation. J Bacteriol. 1999;181(16):4834–41. doi:10.1128/JB.181.16.4834-4841.1999.
- Chauhan A, Sakamoto C, Ghigo JM, Beloin C, Semsey S. Did I pick the right colony? Pitfalls in the study of regulation of the phase variable antigen 43 adhesin. PloS One. 2013;8(9):e73568. doi:10.1371/journal.pone.0073568.
- Faherty CS, Redman JC, Rasko DA, Barry EM, Nataro JP. Shigella flexneri effectors OspE1 and OspE2 mediate induced adherence to the colonic epithelium following bile salts exposure. Mol Microbiol. 2012;85(1):107–21. doi:10.1111/j.1365-2958.2012.08092.x.
- Nickerson KP, Llanos-Chea A, Ingano L, Serena G, Miranda-Ribera A, Perlman M, Lima R, Sztein MB, Fasano A, Senger S. et al. A versatile human intestinal organoid-derived epithelial monolayer model for the study of enteric pathogens. Microbiol Spectr. 2021;9(1):e0000321. doi:10.1128/Spectrum.00003-21.
- Robinson JT, Thorvaldsdottir H, Winckler W, Guttman M, Lander ES, Getz G, Mesirov JP. Integrative genomics viewer. Nat Biotechnol. 2011;29(1):24–26. doi:10.1038/nbt.1754.
- Thorvaldsdottir H, Robinson JT, Mesirov JP. Integrative genomics viewer (IGV): high-performance genomics data visualization and exploration. Brief Bioinform. 2013;14(2):178–92. doi:10.1093/bib/bbs017.
- Deatherage DE, Barrick JE. Identification of mutations in laboratory-evolved microbes from next-generation sequencing data using breseq. Methods Mol Biol. 2014;1151:165–188.