
ABSTRACT
Androgen deprivation therapy, also known as chemical castration, has been used as an adjunct to psychotherapy for sex offenders. Goserelin and bicalutamide are drugs used for chemical castration. Serotonin (5-hydroxytryptamine, 5-HT) is a key neurotransmitter involved in mood changes, such as depression. We investigated the effects of surgical and chemical castration on depressive symptoms in rats. Surgical castration was performed through a bilateral orchiectomy. Bicalutamide was administrated orally once a day for 84 consecutive days. Goserelin acetate was implanted subcutaneously into the anterior abdominal wall, and this implantation was repeated 3 times at 28-day intervals. Testosterone levels were detected by enzyme-linked immunosorbent assays. Sexual behaviors were analyzed by measuring mount latency, mount frequency, intromission latency, and intromission frequency. The forced swimming test was performed to evaluate rats’ depression status. To detect 5-HT and tryptophan hydroxylase (TPH)-positive cells in the dorsal raphe, immunohistochemistry for 5-HT and TPH and western blotting for 5-HT1A receptors and TPH were performed. Surgical castration and goserelin decreased testosterone levels and suppressed sexual behaviors. However, bicalutamide did not inhibit sexual behaviors, although it reduced testosterone levels to a limited extent. Both surgical and chemical castration induced depression in rats. The expression of 5-HT, TPH, and 5-HT1A receptors in the dorsal raphe was significantly decreased by both surgical castration and chemical castration via bicalutamide and goserelin. The present results showed that surgical and chemical castration for 12 weeks induced a depressive state in rats by inhibiting serotonergic function through 5-HT1A receptors.
1. Introduction
Androgen deprivation therapy, also known as chemical castration, has been used as an adjunct to psychotherapy for sex offenders (Gooren Citation2011). Luteinizing hormone-releasing hormone (LHRH) agonists and anti-androgens have been used for androgen-deprivation treatment. Goserelin and bicalutamide are drugs used for chemical castration. Goserelin is a synthetic long-acting agonist of gonadotropin-releasing hormone, and is used for the treatment of malignant neoplasms of the prostate, uterine fibromas, and metastatic breast cancer. Bicalutamide is an androgen receptor inhibitor that is used for the treatment of advanced prostate cancer. These drugs are known to produce clinical effects similar to surgical castration (Lehmusvaara et al. Citation2012). Despite the medical usefulness of these treatments, it has been suggested that chemical castration can cause some side effects such as muscle wasting, anemia, osteoporosis, depression, and reduced sexual desire (Berlin Citation2009). Psychological factors such as depression and loss of libido can have a major negative impact on quality of life.
Serotonin (5-hydroxytryptamine, 5-HT) plays a key role in many physiological processes, including cognition and mood, and 5-HT levels in the brain are affected by depression and anxiety disorders (Biskup et al. Citation2012). Reduced activity of the brain serotonergic system is closely implicated in the pathophysiology of depression (Christiansen et al. Citation2007). Tryptophan hydroxylase (TPH) is a rate-limiting enzyme in serotonin biosynthesis in the raphe nucleus, and modulates serotonergic functions. Thus, TPH is a major candidate for the treatment of psychiatric and behavioral disorders, particularly depression (Roy et al. Citation1999). In addition, tryptophan depletion has been found to increase symptom severity in depressed patients by downregulating 5-HT biosynthesis (Ruhé et al. Citation2007).
The 5-HT receptor family is comprised of 7 receptor classes (Barnes and Sharp Citation1999). Of these, the 5-HT1A receptors are located on serotonin cell bodies in the dorsal raphe, and activation of these receptors suppresses the firing of serotonin neurons and reduces the release of this neurotransmitter in the prefrontal cortex (Albert et al. Citation1996). The inhibition of 5-HT1A receptors in the dorsal raphe may be a mechanism by which serotonergic neurotransmission is reduced in suicide victims with major depression (Stockmeier et al. Citation1998). Antidepressant treatment alleviated depression by enhancing the tonic activation of forebrain postsynaptic 5-HT1A receptors (Haddjeri et al. Citation1998). Moreover, 5-HT1A receptors are strongly implicated in anxiety, depression, and suicide (Popova and Naumenko Citation2013).
Previously, we reported that surgical castration caused deterioration of spatial learning ability, while in contrast, treatment with goserelin or bicalutamide for 28 days did not impair spatial learning ability (Shin et al. Citation2016). In the present study, we investigated the effects of surgical and chemical castration on depressive symptoms using rats. We focused on the long-term administration of goserelin and bicalutamide in relation with 5-HT1A receptors in the dorsal raphe.
2. Materials and methods
2.1. Animals
Adult male Sprague-Dawley rats weighing 300 ± 10 g (8 weeks of age) were obtained from a commercial breeder (Orient Co., Seoul, Korea). The experimental procedures were performed in accordance with the animal care guidelines of the National Institutes of Health, and approved by the Kyung Hee University Institutional Animal Care and Ethics Committee [KHUASP(SE)-15-011]. The animals were housed under controlled temperature (20°C ± 2°C) and lighting (from 07:00 to 19:00) conditions, with food and water made available ad libitum. The male rats were divided into 4 groups (n = 10 in each group): a control group, an orchiectomy group, a bicalutamide treatment group, and a goserelin treatment group.
2.2. Induction of surgical castration
Bilateral orchiectomy was performed according to the previously described method (Shin et al. Citation2016). The rats were anesthetized with 3% isoflurane in 20% O2 and 77% N2. Incisions were made on the center of the scrotum, followed by exposure of each testicle. The vas deferens and main testicular vessels were isolated, ligated, and severed, allowing removal of the testes and epididymis. In the control group, the testes were exposed without removal.
2.3. Drugs
The rats in the bicalutamide treatment group received 10 mg/kg of bicalutamide (Casodex, AstraZeneca) orally once a day for 84 consecutive days. Goserelin acetate (Zoladex, AstraZeneca, Bedfordshire, UK, 3.6 mg; 5 mm long × 1 mm wide biodegradable cylinder) was implanted subcutaneously into the anterior abdominal wall of rats in the goserelin treatment group, and this implantation of goserelin was repeated 3 times at 28-day intervals (drug administration sustained for 84 days.).
2.4. Analysis of sexual behaviors
Sexual behaviors were measured using a video monitoring system, according to a previously described method (JianFeng et al. Citation2012). The female rats were made receptive by sequential administration of estradiol benzoate (10 μg⁄100 g, Tokyo Chemical Industry Co., Tokyo, Japan) and progesterone (0.5 mg/100 g, Tokyo Chemical Industry Co.) by subcutaneous injection at 48 and 4 h prior to pairing. The female rats were introduced to each male rat in a clear cylindrical acrylic container (46 cm × 35 cm) for 30 min (adaptation period) after drug administration.
The following male sexual behavior parameters were calculated over the course of 30 min. Mount latency was defined as the time from the introduction of the female until the first mount. Mount frequency was defined as the number of mounts preceding ejaculation. Intromission latency was defined as the time from the introduction of the female until the first intromission. Intromission frequency was defined as the number of intromissions preceding ejaculation.
2.5. Forced swimming test
The forced swimming test was performed to evaluate the rats’ depressive state, according to a previously described method (Sung et al. Citation2010; Roh et al. Citation2016). On the day after the sexual behavior test, the rats were exposed to a 6-minute test, in which they were placed individually in a vertical Plexiglas cylinder (height, 45 cm; diameter, 19 cm) filled to a height of 30 cm with water at 26°C ± 1°C. Immobility was defined as no activity other than that needed to keep the rat's head above water. Climbing was defined as occurring when the rat was in active vertical motion with its forelegs above the water level. During the test, the duration of immobility and climbing was analyzed using a Smart version 2.5 video tracking system (Panlab, Barcelona, Spain).
2.6. Testosterone measurement
After the forced swimming test, blood from each rat was collected by cardiac puncture, and serum testosterone was analyzed using commercially available testosterone enzyme-linked immunosorbent assay (ELISA) kits (IBL International Gmbh, Hamburg, Germany), according to a previously described method (Shin et al. Citation2016). The absorbance of the content of each well was measured at a wavelength of 450 nm using SpectraMax 190 ELISA reader (Molecular Devices, Sunnyvale, CA, USA).
2.7. Tissue preparation
Transcardial perfusion with 50-mM phosphate-buffered saline (PBS) was performed under anesthesia with Zoletil 50® (10 mg/kg, intraperitoneal; Vibac Laboratories, Carros, France). A freshly prepared solution consisting of 4% paraformaldehyde in 100-mM phosphate buffer (pH 7.4) was used for fixation. Fixed brains were cryoprotected in 30% sucrose for 3 days, and cut into coronal 40-μm thick sections using a freezing microtome (Leica, Nussloch, Germany).
2.8. Immunohistochemistry for 5-HT and TPH
To detect 5-HT-positive and TPH-positive cells in the dorsal raphe, immunohistochemistry was performed, following a previously described method (Seo et al. Citation2013; Roh et al. Citation2016). On average, 6 sections were selected in each brain. Then the sections were incubated in PBS for 10 min and washed 3 times, again with PBS. The sections were then incubated in 1% hydrogen peroxide for 30 min. Next, the sections were incubated overnight with rabbit anti-5-HT antibody (ImmunoStar, Hudson, WI, USA) at a dilution of 1:500 or mouse anti-TPH antibody (Calbiochem, Darmstadt, Germany) at a dilution of 1:500. Then, the sections were incubated for 1 h with biotinylated anti-rabbit secondary antibody or anti-mouse secondary antibody (Vector Laboratories, Burlingame, CA, USA). The sections were subsequently incubated with avidin-biotin-peroxidase complex (Vector Laboratories) for 1 h at room temperature. Immunoreactivity was visualized by incubating the sections in a solution consisting of 0.05% 3,3′-diaminobenzidine and 0.01% hydrogen peroxide in 50-mM Tris buffer (pH 7.6) for approximately 3 min. The sections were then mounted on gelatin-coated glass slides. The slides were air-dried overnight at room temperature, and the coverslips were mounted using Permount® (Thermo Fisher Scientific Inc., Waltham, MA, USA).
2.9. Western blotting for TPH and 5-HT1A receptors
Western blotting for TPH and 5-HT1A receptors was performed, according to a previously described method (Kim et al. Citation2015). Sample tissues were lysed in ice-cold lysate buffer containing 50-mM HEPES (pH, 7.5), 150-mM NaCl, 10% glycerol, 1% Triton X-100, 1.5-mM magnesium chloride hexahydrate, 1-mM ethyleneglycol-bis-(β-aminoethyl ether)-N,N′-tetraacetic acid, 1-mM phenylmethylsulfonyl fluoride, 2 μg/mL of leupeptin, 1 μg/mL of pepstatin, 1-mM sodium orthovanadate, and 100-mM sodium fluoride, after which the mixture was incubated for 30 min at 4°C. The tissue debris was then removed by microcentrifugation, followed by quick freezing of the supernatant. The protein concentration was measured using a Bio-Rad colorimetric protein assay kit (Bio-Rad, Hercules, CA, USA). In this procedure, 40 μg of protein was separated on sodium dodecyl sulfate-polyacrylamide gels and transferred onto a nitrocellulose membrane (Whatman, Clifton, NJ, USA). Mouse anti-TPH antibody (1:1000; Oncogene Research Product, Cambridge, UK) and rabbit anti-5-HT1A antibody (1:1000; Abcam, Cambridge, UK) were used as the primary antibodies. Horseradish peroxidase-conjugated anti-mouse antibody (1:3000; Vector Laboratories) and horseradish peroxidase-conjugated anti-rabbit antibody (1:3000; Vector Laboratories) were used as the secondary antibodies to probe TPH and 5-HT1A. Band detection was performed using the enhanced chemiluminescence detection system (Santa Cruz Biotechnology, Santa Cruz, CA, USA). The bands were quantified using an Image-Pro Plus computer-assisted image analysis system (Media Cybernetics Inc., Bethesda, MD, USA).
2.10. Data analysis
Statistical analysis was performed using 1-way analysis of variance followed by the Duncan post hoc test, and the results were expressed as the mean ± standard error of the mean. Significance was set as p < 0.05.
3. Results
3.1. Testosterone concentrations
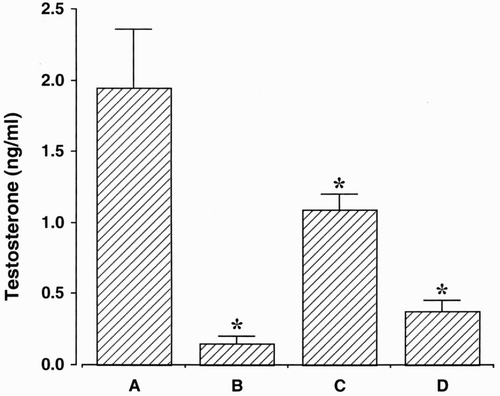
Testosterone concentrations are presented in . The testosterone concentrations were reduced by both surgical castration and chemical castration. Bicalutamide exerted the smallest inhibiting effect on testosterone concentration.
Figure 1. Testosterone concentration. (A) Control group, (B) orchiectomy group, (C) bicalutamide treatment group, and (D) goserelin treatment group. The data is presented as the mean ± standard error of the mean (SEM). * represents p < 0.05 compared to the control group.
3.2. Sexual behaviors
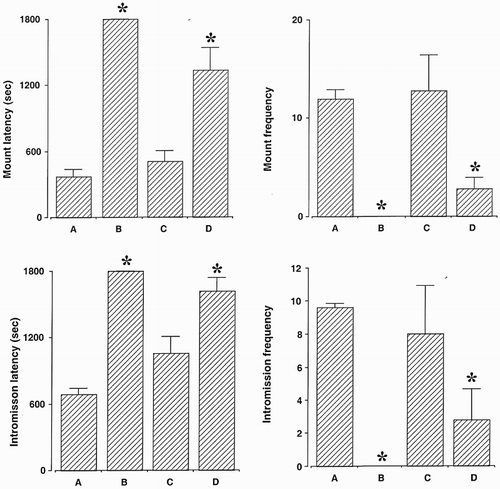
Sexual behaviors are presented in . Mount latency was increased and mount frequency was reduced by surgical castration and goserelin treatment. Intromission latency was increased and intromission frequency was reduced by surgical castration and goserelin treatment. Orchiectomy and goserelin treatment inhibited sexual behaviors, while bicalutamide treatment exerted no significant effect on sexual behaviors.
Figure 2. Sexual behaviors. (A) Control group, (B) orchiectomy group, (C) bicalutamide treatment group, and (D) goserelin treatment group. The results are presented as the mean ± standard error of the mean (SEM). * represents p < 0.05 compared to the control group.
3.3. Depressive symptoms
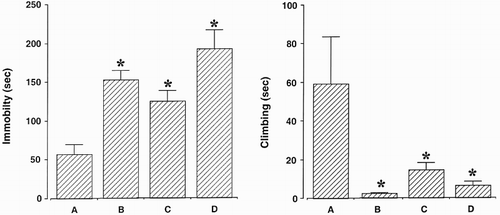
The results of the forced swimming test are presented in . Immobility time was increased and climbing time was reduced by both surgical castration and chemical castration. The present results showed that orchiectomy, bicalutamide, and goserelin induced depressive symptoms. Bicalutamide exerted the smallest effect on depressive symptoms.
Figure 3. Immobility and climbing time. (A) Control group, (B) orchiectomy group, (C) bicalutamide treatment group, and (D) goserelin treatment group. The results are presented as the mean ± standard error of the mean (SEM). * represents p < 0.05 compared to the control group.
3.4. The numbers of 5-HT and TPH in the dorsal raphe
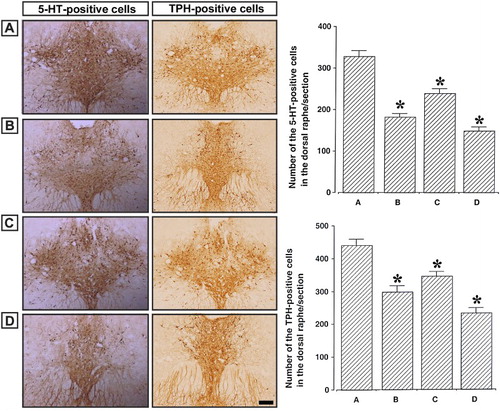
Photomicrographs of 5-HT-positive and TPH-positive cells are presented in . Both surgical and chemical castration inhibited the numbers of 5-HT-positive cells and TPH-positive cells in the dorsal raphe. Bicalutamide showed the smallest inhibitory effect on the numbers of 5-HT-positive cells and TPH-positive cells among the 3 groups.
Figure 4. 5-Hydroxytryptamine (5-HT) and tryptophan hydroxylase (TPH). Photomicrographs showing TPH and 5-HT expressions in the dorsal raphe. The scale bar represents 100 µm. (A) Control group, (B) orchiectomy group, (C) bicalutamide treatment group, and (D) goserelin treatment group. The results are presented as the mean ± standard error of the mean (SEM). * represents p < 0.05 compared to the control group.
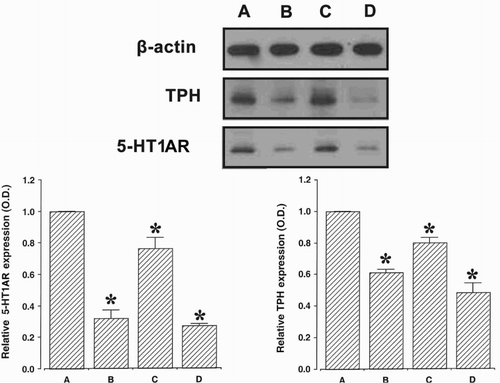
3.5. Expression of the 5-HT1A receptor and TPH in the dorsal raphe
The expression of the 5-HT1A receptor and TPH is presented in . The expression of the 5-HT1A receptor and TPH in the dorsal raphe was suppressed by both surgical castration and chemical castration. Bicalutamide showed the smallest inhibitory effect on the expression of the 5-HT1A receptor and TPH among the 3 groups.
Figure 5. 5-Hydroxytryptamine-1A receptors (5-HT1AR) and tryptophan hydroxylase (TPH). (A) Control group, (B) orchiectomy group, (C) bicalutamide treatment group, and (D) goserelin treatment group. Data are presented as the means ± standard error of the mean (SEM). * represents p < 0.05 compared to the control group.
4. Discussion
Testosterone deficiency is linked with depression in men, since testosterone plays an important role in mood regulation (Zitzmann Citation2006). Carrier and Kabbaj (Citation2012) found that testosterone ameliorated depressive-like behavior in male rats. Ko et al. (Citation2016) demonstrated a correlation between serum testosterone levels and chemical castration regimens. In the present study, serum testosterone levels in the surgical castration group and the goserelin group were lower than in the control group. These results suggest that castration was induced by both orchiectomy and goserelin treatment. However, bicalutamide treatment reduced testosterone levels slightly. Bicalutamide is known to exert its effect through the competitive inhibition of androgen receptors, and it is known to have little effect on serum testosterone levels (Furr and Tucker Citation1996). Motofei et al. (Citation2011) reported that bicalutamide exerted no significant effect on libido in heterosexual men with prostate cancer.
According to the data found in the present study, surgical castration and goserelin lowered testosterone levels and suppressed sexual activity. However, bicalutamide did not inhibit sexual behavior and showed only some inhibition of testosterone.
Decreases in androgens due to aging are associated with negative mood and anxiety in humans (Seidman Citation2003). A high incidence of major depressive disorder was observed in patients with low serum testosterone levels, and testosterone therapy effectively improved their mood (McIntyre et al. Citation2006). Testosterone has a protective effect against the development of depression, and testosterone administration showed an antidepressant effect on aged mice (Frye and Walf Citation2009).
Behavioral parameters such as climbing time and immobility time are interpreted as indicators of depression in the forced swimming test. Increased immobility time has been interpreted as a failure of persistence in escape-oriented behavior (Cryan et al. Citation2002). A decrease in climbing time and an increase in immobility time in maternal rats isolated from their offspring was interpreted as an indicator of depression-like behaviors (Sung et al. Citation2010). Alcohol administration also resulted in depressed behaviors, such as decreased climbing time and increased immobility time, in gerbils (Kim et al. Citation2012). Orchiectomy significantly increased immobility and decreased swimming and climbing until 60 days after castration. In contrast, nicotine reduced immobility and increased swimming in rats (Bonilla-Jaime et al. Citation2010). Orchiectomy suppressed the antidepressant effect of nicotine (Bonilla-Jaime et al. Citation2010). Ahmadi-Davis et al. (Citation2014) reported that goserelin treatment caused major depressive disorder in a man with prostate cancer. In the present study, the immobility time was prolonged and swimming time was shortened by surgical castration and chemical castration via bicalutamide and goserelin. These results indicate that both surgical castration and chemical castration induced depression in rats.
Baumann et al. (Citation2002) found that the number of neurons was decreased in the dorsal raphe of depressed patients. The dorsal raphe is closely related to mood disorders such as depression (Michelsen et al. Citation2008). Dysfunction of the serotonergic system is considered to be a hallmark of depressive disorder (Jans et al. Citation2007). TPH has two isoforms: TPH-1 and TPH-2. TPH2 mRNA expression was found to be increased in depressed individuals who committed suicide (Bach-Mizrachi et al. Citation2006). TPH-1 showed a strong correlation with stress-induced depression, but TPH-2 was not associated with major depression (Gizatullin et al. Citation2008). Maternal rats isolated from their offspring showed a depressed state, with decreased expression of 5-HT and TPH in the dorsal raphe (Sung et al. Citation2010). Alcohol consumption induced a depressed state by increasing 5-HT and TPH expression in the dorsal raphe of rats (Kim et al. Citation2012). In the present study, the expression of 5-HT and TPH in the dorsal raphe was significantly decreased by surgical castration and chemical castration via bicalutamide and goserelin.
It has been well established that serotonergic dysfunction, especially dysfunction in 5-HT1A receptors, plays an important role in the development of major depressive disorder (Kaufman et al. Citation2016). It was found that 5-HT expression in the dorsal raphe was enhanced by upregulation of the 5-HT1A receptors, which improved stress-related depression (Kim et al. Citation2015). The antidepressant effect of exercise was found to be achieved via enhanced serotonin function due to upregulation of the 5-HT1A receptors in the dorsal raphe (Shin et al. Citation2017). In the present study, the expression of 5-HT1A receptors in dorsal raphe was significantly decreased by surgical castration and chemical castration via bicalutamide and goserelin.
The present results showed that surgical castration and chemical castration for 12 weeks induced a depressive state by enhancing serotonergic function through 5-HT1A receptors in rats. The results of this experiment suggest the need for ancillary treatment for depression when using chemical castration.
Disclosure statement
No potential conflict of interest was reported by the authors.
ORCID
Sung-Eun Kim http://orcid.org/0000-0003-3742-0271
Additional information
Funding
References
- Ahmadi-Davis S, Velasco R, Stewart JT. 2014. Goserelin-induced depression in a man with prostate cancer. Psychosomatics. 55:720–722. doi: 10.1016/j.psym.2013.12.008
- Albert PR, Lembo P, Storring JM, Charest A, Saucier C. 1996. The 5-HT1A receptor: signaling, desensitization, and gene transcription. Neuropsychopharmacology. 14:19–25. doi: 10.1016/S0893-133X(96)80055-8
- Bach-Mizrachi H, Underwood MD, Kassir SA, Bakalian MJ, Sibille E, Tamir H, Mann JJ, Arango V. 2006. Neuronal tryptophan hydroxylase mRNA expression in the human dorsal andmedian raphe nuclei: major depression and suicide. Neuropsychopharmacology. 31:814–824. doi: 10.1038/sj.npp.1300897
- Barnes NM, Sharp T. 1999. A review of central 5-HT receptors and their function. Neuropharmacology. 38:1083–1152. doi: 10.1016/S0028-3908(99)00010-6
- Baumann B, Bielau H, Krell D, Agelink MW, Diekmann S, Wurthmann C, Trübner K, Bernstein HG, Danos P, Bogerts B. 2002. Circumscribed numerical deficit of dorsal raphe neurons in mood disorders. Psychol Med. 32:93–103. doi: 10.1017/S0033291701004822
- Berlin FS. 2009. Risk/benefit ratio of androgen deprivation treatment for sex offenders. J Am Acad Psychiatry Law. 37:59–62.
- Biskup CS, Sánchez CL, Arrant A, Van Swearingen AE, Kuhn C, Zepf FD. 2012. Effects of acute tryptophan depletion on brain serotonin function andconcentrations of dopamine and norepinephrine in C57BL/6J and BALB/cJmice. PLoS One. 7:e35916. doi: 10.1371/journal.pone.0035916
- Bonilla-Jaime H, Limón-Morales O, Arteaga-Silva M, Hernández-González M, Guadarrama-Cruz G, Alarcón-Aguilar F, Vázquez-Palacios G. 2010. Orchiectomy modifies the antidepressant-like response of nicotine in the forced swimming test. Physiol Behav. 101:456–461. doi: 10.1016/j.physbeh.2010.07.011
- Carrier N, Kabbaj M. 2012. Extracellular signal-regulated kinase 2 signaling in the hippocampal dentate gyrus mediates the antidepressant effects of testosterone. Biol Psychiatry. 71:642–651. doi: 10.1016/j.biopsych.2011.11.028
- Christiansen L, Tan Q, Iachina M, Bathum L, Kruse TA, McGue M, Christensen K. 2007. Candidate gene polymorphisms in the serotonergic pathway: influence on depression symptomatology in an elderly population. Biol Psychiatry. 61:223–230. doi: 10.1016/j.biopsych.2006.03.046
- Cryan JF, Markou A, Lucki I. 2002. Assessing antidepressant activity in rodents: recent developments and future needs. Trends Pharmacol Sci. 23:238–245. doi: 10.1016/S0165-6147(02)02017-5
- Frye CA, Walf AA. 2009. Depression-like behavior of aged male and female mice is ameliorated with administration of testosterone or its metabolites. Physiol Behav. 97:266–269. doi: 10.1016/j.physbeh.2009.02.022
- Furr BJ, Tucker H. 1996. The preclinical development of bicalutamide: pharmacodynamics and mechanism of action. Urology. 47(1A Suppl):13–25. doi: 10.1016/S0090-4295(96)80003-3
- Gizatullin R, Zaboli G, Jönsson EG, Asberg M, Leopardi R. 2008. The tryptophan hydroxylase (TPH) 2 gene unlike TPH-1 exhibits no association with stress-induced depression. J Affect Disord. 107:175–179. doi: 10.1016/j.jad.2007.07.005
- Gooren LJ. 2011. Clinical review: ethical and medical considerations of androgen deprivation treatment of sex offenders. The J Clin Endocrinol Metab. 96:3628–3637. doi: 10.1210/jc.2011-1540
- Haddjeri N, Blier P, de Montigny C. 1998. Long-term antidepressant treatments result in a tonic activation of forebrain 5-HT1A receptors. J Neurosci. 18:10150–10156.
- Jans LA, Riedel WJ, Markus CR, Blokland A. 2007. Serotonergic vulnerability and depression: assumptions, experimental evidence and implications. Mol Psychiatry. 12:522–543. doi: 10.1038/sj.mp.4001920
- JianFeng C, PengYing Z, ChengWei X, TaoTao H, YunGui B, KaoShan C. 2012. Effect of aqueous extract of Arctium lappa L. (burdock) roots on the sexual behavior of male rats. BMC Complement Altern Med. 12:8. doi: 10.1186/1472-6882-12-8
- Kaufman J, DeLorenzo C, Choudhury S, Parsey RV. 2016. The 5-HT1A receptor in major depressive disorder. Eur Neuropsychopharmacol. 26:397–410. doi: 10.1016/j.euroneuro.2015.12.039
- Kim JE, Ji ES, Seo JH, Lee MH, Cho S, Kim Park Y, Seo TB, Kim CJ. 2012. Alcohol exposure induces depression-like behavior by decreasing hippocampal neuronal proliferation through inhibition of the BDNF-ERK pathway in gerbils. Anim Cells Syst. 16:190–197. doi: 10.1080/19768354.2011.640352
- Kim TW, Lim BV, Baek D, Ryu DS, Seo JH. 2015. Stress-induced depression is alleviated by aerobic exercise through up-regulation of 5-hydroxytryptamine 1A receptors in rats. Int Neurourol J. 19:27–33. doi: 10.5213/inj.2015.19.1.27
- Ko DH, Lee K, Jeon SH, Song SH, Yun YM, Chun S, Kim HS, Kim JY, In MK, Song J. 2016. Simultaneous measurement of serum chemical castration agents and testosterone levels using ultra-performance liquid chromatography-tandem mass spectrometry. J Anal Toxicol. 40:294–303. doi: 10.1093/jat/bkw017
- Lehmusvaara S, Erkkilä T, Urbanucci A, Waltering K, Seppälä J, Larjo A, Tuominen VJ, Isola J, Kujala P, Lähdesmäki H, et al. 2012. Chemical castration and anti-androgens induce differential gene expression in prostate cancer. J Pathol. 227:336–345. doi: 10.1002/path.4027
- McIntyre RS, Mancini D, Eisfeld BS, Soczynska JK, Grupp L, Konarski JZ, Kennedy SH. 2006. Calculated bioavailable testosterone levels and depression in middle-aged men. Psychoneuroendocrinology. 31:1029–1035. doi: 10.1016/j.psyneuen.2006.06.005
- Michelsen KA, Prickaerts J, Steinbusch HW. 2008. The dorsal raphe nucleus and serotonin: implications for neuroplasticity linked to major depression and Alzheimer’s disease. Prog Brain Res. 172:233–264. doi: 10.1016/S0079-6123(08)00912-6
- Motofei IG, Rowland DL, Popa F, Kreienkamp D, Paunica S. 2011. Preliminary study with bicalutamide in heterosexual and homosexual patients with prostate cancer: a possible implication of androgens in male homosexual arousal. BJU Int. 108:110–115. doi: 10.1111/j.1464-410X.2010.09764.x
- Popova NK, Naumenko VS. 2013. 5-HT1A receptor as a key player in the brain 5-HT system. Rev Neurosci. 24:191–204.
- Roh JH, Ko IG, Kim SE, Lee JM, Ji ES, Kim JH, Chang HK, Lee SK, Kim KH. 2016. Treadmill exercise ameliorates intracerebral hemorrhage-induced depression in rats. J Exerc Rehabil. 12:299–307. doi: 10.12965/jer.1632692.346
- Roy A, Nielsen D, Rylander G, Sarchiapone M, Segal N. 1999. Genetics of suicide in depression. J Clin Psychiatry. 60:12–17.
- Ruhé HG, Mason NS, Schene AH. 2007. Mood is indirectly related to serotonin, norepinephrine and dopamine levels in humans: a meta-analysis of monoamine depletion studies. Mol Psychiatry. 12:331–359. doi: 10.1038/sj.mp.4001949
- Seidman SN. 2003. Testosterone deficiency and mood in aging men: pathogenic and therapeutic interactions. World J Biol Psychiatry. 4:14–20. doi: 10.3109/15622970309167905
- Seo JH, Kim TW, Kim CJ, Sung YH, Lee SJ. 2013. Treadmill exercise during pregnancy ameliorates post-traumatic stress disorder-induced anxiety-like responses in maternal rats. Mol Med Rep. 7:389–395. doi: 10.3892/mmr.2012.1197
- Shin MS, Chung KJ, Ko IG, Kim SH, Jin JJ, Kim SE, Lee JM, Ji ES, Kim TW, Cho HS, et al. 2016. Effects of surgical and chemical castration on spatial learning ability in relation to cell proliferation and apoptosis in hippocampus. Int Urol Nephrol. 48:517–527. doi: 10.1007/s11255-015-1200-0
- Shin MS, Park SS, Lee JM, Kim TW, Kim YP. 2017. Treadmill exercise improves depression-like symptoms by enhancing serotonergic function through upregulation of 5-HT1A expression in the olfactory bulbectomized rats. J Exerc Rehabil. 13:36–42. doi: 10.12965/jer.1734918.459
- Stockmeier CA, Shapiro LA, Dilley GE, Kolli TN, Friedman L, Rajkowska G. 1998. Increase in serotonin-1A autoreceptors in the midbrain of suicide victims with major depression-postmortem evidence for decreased serotonin activity. J Neurosci. 18:7394–7401.
- Sung YH, Shin MS, Cho S, Baik HH, Jin BK, Chang HK, Lee EK, Kim CJ. 2010. Depression-like state in maternal rats induced by repeated separation of pups is accompanied by a decrease of cell proliferation and an increase of apoptosis in the hippocampus. Neurosci Lett. 470:86–90. doi: 10.1016/j.neulet.2009.12.063
- Zitzmann M. 2006. Testosterone and the brain. Aging Male. 9:195–199. doi: 10.1080/13685530601040679