
Abstract
Objectives: The aim of this review was to highlight recent advances in our understanding of the pathogenesis of malignant transformation of endometriosis.
Methods: This study reviewed the English-language literature concerning basic science studies of the potential promotion of carcinogenesis.
Results: Repeated episodes of hemorrhage occur in endometriosis at the onset of menstruation. Extracellular hemoglobin, heme, and iron derivatives in endometriosis cause DNA damage and mutations, which create increased cellular susceptibility to oxidant-mediated cell killing. Excess DNA damage and mutations are linked to cell death, but not carcinogenesis. In response to an oxidative and inflammatory microenvironment, endometriotic cells and macrophages secrete antioxidants that control excess oxidative stress in the surrounding environment. Exposure of endometriotic cells to a sublethal level of oxidative stress may lead to carcinogenesis. Macrophages also secrete immunosuppressive factors that lead to promotion of malignant transformation.
Discussion: At least two potential scenarios could result in ovarian cancer arising from endometriosis. The first step: extracellular hemoglobin, heme, and iron cause cellular oxidative damage by promoting reactive oxygen species formation, which results in DNA damage and mutations (ovarian cancer initiation from endometriosis). The second step: cancer progression may be associated with persistent antioxidant production favoring a protumoral microenvironment.
Introduction
A new paradigm for the pathogenesis of ovarian cancer presupposes two distinct types of molecular profiles: type I and type II cancers. Type I cancer includes clear cell carcinoma, mostly arising from endometriosis. Oxidative stress within endometriosis is likely to contribute to the malignant transformation process. Therefore, in the present review, we focused on the scheme of possible involvement in the development of endometriosis-associated ovarian cancer.
Endometriosis is associated with pelvic pain, female infertility and, rarely, malignant transformation. The malignant transformation of endometriosis is an uncommon event, but happens in ∼1.0% of the cases in Japan.Citation1 Endometriosis increases the risk for endometrioid and clear cell ovarian cancers.Citation1 Ovarian cancers exhibit diverse phenotypes depending on their distinct physical, pathological, clinical, and therapeutic aspects.Citation1 Mechanistic explanations of the pathogenesis of malignant transformation are challenging, but its pathophysiology may be closely associated with fine-tuning of the balance between pro-inflammatory (iron overload) and anti-inflammatory (antioxidant defense) environments in endometriosis.Citation2,Citation3
Menstruation is associated with an increased concentration of red and white blood cells in peritoneal fluid.Citation4 Although retrograde flux at menstruation might give rise to both non-endometriosis and endometriosis in women, the retrograde menstruation theory represents the pathogenetic basis of endometriosis.Citation4 The major physiological function of hemoglobin is to transport oxygen to the peripheral tissues.Citation5 Heme is incorporated into four globin subunits (α2β2) that are required to generate stable hemoglobin tetramers. Heme consists of protoporphyrin IX and a ferrous (Fe2+) iron and constitutes 95% of functional iron in the human body.Citation5 The majority of heme proteins is identified as hemoglobin in animals and humans. Other heme proteins include myoglobin, cytochromes, and some hemoproteins such as catalytic enzymes, which play critical physiological roles including oxygen transport and mitochondrial respiration.Citation5 Hemolysis results in the transition of ferrous to ferric (Fe3+) forms of hemoglobin, which leads to oxidation. Extracellular free hemoglobin, heme, and iron derivatives are critical components of the pathological process and induce a potential toxicity through catalytic activities during hemolysis.Citation6 These compounds are abundantly present in the endometriotic cyst fluid or peritoneal fluid during menstruation.Citation7,Citation8
Hemoglobin derivatives, particularly oxyhemoglobin (hemoglobin containing ferrous iron Fe2+) and methemoglobin (hemoglobin containing ferric iron Fe3+), have multiple pathophysiologic effects when released into the intracystic or intraperitoneal space during hemolysis.Citation9 Free hemoglobin increases oxidative stress via its catalytic activity of heme and iron to generate hydroxyl radicals.Citation7 Endometriosis is vulnerable to oxidative stress and induces DNA damage and cell death. On the other hand, this effect includes induction of genomic instability and oncogenic transformation in endometriotic tissues.Citation9 Taken together, pathophysiological factors in endometriosis lead to DNA damage and mutations, which contribute to either beneficial (cell death) or detrimental (carcinogenesis) outcomes.
This conceptual dichotomy is explored in this review, with a focus on fine-tuning of the balance between oxidative stress and antioxidant effects. This review also outlines several suggested mechanisms for the pathogenesis of endometriosis carcinogenesis. Understanding this mechanism is fundamental for early detection and identification of critical molecular features for carcinogenesis and targeted therapeutics.
Search strategy and selection criteria
A computerized literature search was performed to identify relevant studies reported in the English language. We searched MEDLINE electronic databases (http://www.ncbi.nlm.nih.gov/sites/entrez) published between 1966 and 2014, combining the keywords ‘endometriosis’, ‘ovarian cancer’, ‘hemoglobin’, ‘heme’, ‘iron’, ‘oxidative stress’, ‘antioxidant’, and ‘pathogenesis’. Various combinations of the terms were used, depending on the database searched. Each gene was also linked to NCBI Entrez Gene pages (http://www.ncbi.nlm.nih.gov/sites/entrez). In addition, references in each article were searched to identify potentially missed studies. There were 592 articles identified by the search. About 134 articles were potentially relevant. Thirty-seven publications were chosen based on the final selection taking into account the title and the summary analysis.
Free hemoglobin and heme toxicities
Red blood cells in the extravascular space tend to lyse quickly.Citation9 When hemoglobin appears in the extracellular milieu, it is prone to autoxidation and easily oxidized from ferrous hemoglobin (oxyhemoglobin) to ferric hemoglobin (methemoglobin), which readily releases heme and superoxide (O2−) or perhydroxy (HO2−) radical.Citation9 Heme is highly cytotoxic, potently hemolytic, and destabilizes the cytoskeleton, mitochondria, nuclei, and cell membrane, as well as subsequently inducing lipid peroxidation and DNA damage.Citation6,Citation10 Free heme promotes oxidative damage and formation of reactive oxygen species (ROS), which, in turn, split the heme ring and release redox-active free iron.Citation11 Heme is a source of total and free iron. Our previous article showed what is known about the contribution of iron to endometriosis development.Citation12 Free iron can contribute to the production of ROS via Fenton chemistry and create increased cellular susceptibility to oxidant-mediated killing.Citation12,Citation13 Thus, hemoglobin overload leads to accumulation of heme and iron, with further ROS generation, and induces apoptosis and cell death, which is beneficial for women with endometriosis.Citation9–Citation12
The molecular mechanisms regulating hemoglobin metabolism
There are essential mechanisms that prevent the toxicity of extracellular hemoglobin.Citation14 Hemoglobin and heme affect the signaling of several important detoxification systems. During physiological and pathological hemolysis, two major uptake systems are represented by (1) the CD163 pathway for uptake of hemoglobin–haptoglobin (Hp) complexes and (2) the CD91 pathway for uptake of hemopexin (Hx)–heme complexes.Citation14 The endocytosis of the ligands leads to Hb degradation in lysosomes of macrophages. CD163 is a macrophage-specific protein and acts as the Hb scavenger receptor.Citation14 CD91 is also known as the low-density lipoprotein receptor-related protein.
The Hp–CD163–HO-1 backup protecting system
Activated macrophages represent the first line of defense against free hemoglobin.Citation14 Hp is an initial key player of the detoxification process. Hemoglobin is tightly bound to Hp, leading to CD163-mediated endocytosis in macrophages. The endometriotic cyst fluid concentration of Hp may be a limiting factor during accelerated hemolysis. The CD163-mediated signaling upregulates several genes, including heme oxygenase (HO)-1, which catalyzes conversion of heme into the metabolites carbon monoxide (CO), biliverdin, and iron.Citation15 The hemoglobin–Hp–CD163–HO-1 system produces anti-inflammatory heme metabolites, suppresses pro-inflammatory cytokines, and protects endometriotic tissues from free hemoglobin-mediated oxidative damage.Citation14 Inflammatory cytokines, interleukin (IL)-6, IL-10, and glucocorticoids stimulate Hp production in macrophages.Citation14,Citation16 HO-1 is induced in response to not only free hemoglobin and heme, but also ROS, biliverdin, and CO.Citation17 It is known that CO protects cells against various types of organ injury. The HO system acts as a potent antioxidant and contributes to the resolution of inflammation, oxidative stress, apoptosis, and cell death. The HO-1 pathway also induces ferritin expression.Citation17 The toxic iron released upon heme degradation is maintained within the iron storage protein ferritin and also is promptly sequestered by ferritin. The endometriotic cell may either die or survive by induction of the HO-1-mediated cytoprotective pathways. A small subset of cells is exceptionally able to survive under excessive oxidative stress conditions while most cells die.
The molecular mechanisms regulating heme metabolism
Here we review specific scavenging systems, including heme export mechanisms and intracellular defenses against heme.
Heme export system
Excess heme is excreted out of the cell across the plasma membrane by heme exporters, FLVCR1a (feline leukemia virus subgroup C cellular receptor 1a) and ABCG2 (ATP-binding cassette, sub-family G (WHITE), member 2).Citation18 FLVCR1a plays a critical role in protecting erythroid cells from the toxic accumulation of heme. ABCG2, the superfamily of ATP-binding cassette exporters, transports heme out of the cell. The density of ABCG2-positive endothelial cells is reduced in ectopic endometrium compared with eutopic endometrium.Citation19
Intracellular defense against heme: the Hx–CD91–HO-1 backup protecting system
The toxic heme is endocytosed by Hx, Hp, LRP1 (low-density lipoprotein receptor-related protein 1, also known as CD91), LRP2, α-1-microglobulin/bikunin precursor, lipoproteins, or hydrophobic intercalation.Citation11 Hx, a heme-binding glycoprotein, is the major transport protein for free heme. Hx–heme complexes bind to the endocytic receptor CD91 and are removed by receptor-mediated endocytosis in macrophages. The heme–Hx–CD91–HO-1 pathway is an essential backup system to detoxify free heme.Citation11 Free heme binding to α-1-microglobulin (A1M) also triggers the formation of a stable A1M trimer and keeps heme at safe levels.Citation11 Heme effectively induces the cytoprotective enzymes, HO-1 and HO-2.Citation11
Regulation of HO-1 induction
In general, HO-1 is induced by several regulatory oxidant-sensitive transcription factors (e.g. nuclear factor-kappaB (NF-κB), activator protein-1 (AP-1), nuclear factor, erythroid 2 (Nrf2), Maf recognition element (MARE), DNA-binding response regulator cAMP responsive element-binding protein, biliverdin reductase, and activating transcription factor 2) and specific kinases (e.g. mitogen-activated protein kinase (MAPK), protein kinase C, protein kinase A, and phosphatidylinositol-3-kinase).Citation20 A transcription factor, Nrf2, is a major regulator of cytoprotection during oxidative stress and regulates genes that contain antioxidant response elements in their promoters.Citation20 The Nrf2–Maf complexes bind to MARE and promote HO-1 gene transcription for executing an anti-oxidative stress response.Citation21 Nrf2 also produces many cytoprotective genes, including ferritin, superoxide dismutase, glutathione peroxidase, and catalase. Furthermore, free heme itself exports a transcription factor Bach1 (BTB and CNC homology 1, basic leucine zipper transcription factor 1) from the nucleus and releases transcriptional repression of the HO-1 gene, leading to overexpression of HO-1 protein.Citation20,Citation21 Heme also activates the MAPK pathway, leading to activation of Nrf2 and subsequent induction of defensive genes.Citation20,Citation21 HO-1 and Nrf2 are important regulators for antioxidants that are highly inducible by a variety of stimuli including Hb, heme, iron, and oxidative stress.
The prooxidant–antioxidant balance in endometriosis
Endometriosis is characterized by recurring episodes of dysmenorrhea, infertility and, rarely, malignant transformation.Citation1 This disorder is a devastating hemorrhagic and hemolytic disease caused by repeated retrograde menstruation (Fig. ). Severe hemolysis occurring during the development of endometriosis results in the production of free Hb, heme, and iron, which cause toxicity and lead to cellular, tissue, and organ injury.Citation22 These compounds act as pro-inflammatory molecules and oxidatively modify lipids and proteins, leading to cell and DNA damage, and subsequently fibrosis development in the peritoneal epithelial cells and endometriotic cells.Citation22 By microarray studies, endometriosis-specific genes are involved in oxidative stress and detoxification systems that fine-tune the balance between pro-inflammatory and anti-inflammatory environments.Citation23 Excess ferrous iron activates the caspase pathways, through enhanced expression of caspase-8 and reduced expression of Bcl-2 (B-cell CLL/lymphoma 2) and Bcl-xL (BCL2-like 1), leading to apoptosis.Citation24 On the other hand, excessive ROS production activates redox-sensitive transcription factors such as NF-κB and AP-1, which in turn promote increased expression of antioxidants, including Hp, CD163, and HO-1.Citation11
Figure 1 Scheme of possible involvement in the development of endometriosis-associated ovarian cancer. Based on the observations discussed we propose that hemoglobin-, heme-, and iron-induced oxidative stress and antioxidants are two significant factors that overlap and presumably exacerbate each other in endometriosis-associated ovarian carcinogenesis. Hemoglobin, heme, and iron associated with oxidative stress can initially induce DNA damage, genomic instability, mutations, and aberrant pro-tumorigenic signaling. High ROS levels are toxic to cells, induce cell death, and potentially block tumorigenesis. To balance oxidative stress, macrophages increase their antioxidant capacity, which strongly suggests that a sublethal level of ROS plays a causal role in cancer progression. We propose the two-step theory for the scheme of possible involvement in the development of endometriosis-associated ovarian cancer. Abbreviations: Hb, hemoglobin; Hp, haptoglobin; HO-1, heme oxygenase-1; CO, carbon monoxide; Nrf2, nuclear factor, erythroid 2; ROS, reactive oxygen species.
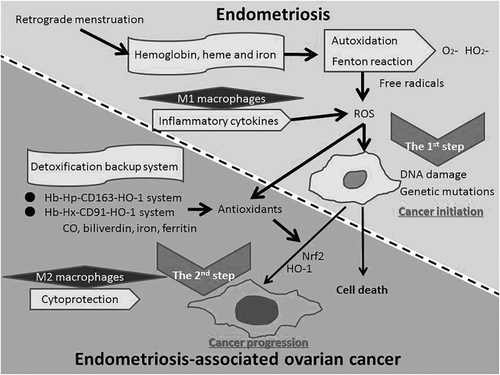
Furthermore, an endometriosis-specific transcription factor, hepatocyte nuclear factor-1β, serves to alleviate damage and promote survival of endometriotic cells by upregulating antioxidant protein expression.Citation3 Notwithstanding elevated quantities of antioxidants in endometriosis, antioxidant activities may be depleted due to substantially higher levels of oxidative stress, enhancing sensitivity to oxidative stress in this disease. Serum oxidative stress parameters were higher, whereas antioxidant defense parameters were relatively lower, in women with endometriosis than in control.Citation25 This imbalance may lead to the development of endometriosis-associated infertility and dysmenorrhea.
Nerve growth factor (NGF) is one of the oxidative stress-sensitive neurotrophic genes.Citation26,Citation27 NGF mRNA and protein were overexpressed in endometriotic lesions relative to eutopic endometrium.Citation26,Citation27 This growth factor produced locally in the peritoneal cavity may be involved in the generation of endometriosis-associated pelvic pain.Citation26,Citation27 Heme participates in the regulation of neuronal-specific neurotrophin genes, through the NGF signaling pathway.Citation28 Therefore, heme and NGF have been proposed as the key factors responsible for the onset and maintenance of pain in endometriosis.Citation27
In endometriosis, HO-1 subserves cytoprotective responses to toxic insults. This enzyme is strongly expressed in ectopic endometrium, especially in red lesions.Citation29 Because free Hb is toxic, endometriotic cells elaborate scavenging factors to keep its intracellular concentration at low levels. Endometriotic cells display higher endogenous oxidative stress compared with an increase in antioxidant levels.Citation30 Therefore, the prooxidant–antioxidant balance values are higher in women with endometriosis compared with controls. These results suggest that Hb, heme, and iron are involved in the pathogenesis or development of endometriosis and that the HO system, although expressed, might be insufficient to detoxify heme in women with endometriosis.Citation29 High levels of ROS are toxic to endometriotic cells and may induce cell death or suppress tumorigenesis.
Haptoglobin
Hp is increased in the peritoneal fluid of women with endometriosis compared to those without disease.Citation31 In response to the inflammatory cytokines IL-1β, IL-6, and TNFα, isolated endometrial stromal cells from women with endometriosis secrete Hp mRNA and protein.Citation32 Endometriotic lesions synthesize and secrete higher levels of Hp.Citation33 However, Hp was identified to be decreased in the plasma levels of women with endometriosis.Citation34 Furthermore, endometriosis protein-I (ENDO-I) gene is a unique form of Hp. The ENDO-I gene displays 99% homology with Hp and is also synthesized and secreted by endometriotic lesions.Citation6
CD163
Scavenger receptor (CD163 and CD206)-positive macrophages were abundant in endometriotic lesions.Citation35 CD206+ macrophages express HO-1, which also promotes the differentiation of macrophages to the M2 phenotype. CD163 and CD206 are involved in both protection against inflammatory molecules and scavenging of Hb with iron transfer into macrophages.Citation36
Hemopexin
This transporter provides anti-oxidative activity against heme or ROS toxicity via HO-1. Women with endometriosis have lower peritoneal fluid levels of Hx.Citation37 Recent data implicate a role of altered heme and iron metabolism in endometriosis patients.Citation37 Therefore, reduced Hx concentrations and heme scavenging in peritoneal fluid might be involved in the pathophysiology of endometriosis.
Transferrin receptor
Higher iron concentrations and transferrin (Tf) saturations were detected in endometriosis.Citation38 Iron uptake is mediated by the binding of Tf to the transferrin receptor (TfR), a putative iron sensor, by endocytosis.Citation39 The TfR-positive cells were increased in infertile women with endometriosis when compared with the control groups. Accordingly, a high TfR expression is a characteristic of endometriosis responding to inflammation. Ferritin expression in peritoneal macrophages was higher in endometriosis patients than in controls.Citation38
Iron regulatory proteins
Iron regulatory proteins 1 and 2 (IRP1 and IRP2) modulate the expression of several mRNAs encoding proteins of iron metabolism.Citation40 They bind to the iron responsive elements in ferritin and TfR1 mRNA and thereby inhibit translation of ferritin and protect TfR1 against degradation, respectively.Citation40 Interestingly, ectopic endometriotic stromal cells exhibited high IRP1 and IRP2 expression.Citation41
Characteristics of macrophage polarization
Endometriosis is a sterile inflammation, which might be driven by damage-associated molecular patterns.Citation42 Macrophage-driven inflammatory networks and their secretory cytokines are essential for endometriosis. It has been proposed that macrophages be classified into M1 macrophages and M2 macrophagesCitation43 (Fig. ). M1 macrophages are characterized by a pro-inflammatory phenotype and are key players of the first line of defense against bacterial infections. After acute inflammation, M2 macrophages within regenerating tissues display regulatory functions in tissue repair, remodeling, wound healing, and promotion of protumoral immune responses.Citation44 M2 macrophages stimulated by Th2 cytokines such as IL-4 or IL-13 can produce anti-inflammatory factors such as IL-10.Citation45 The secretion signature of cytokines exhibits characteristics of both classes of macrophage polarization, i.e. marked M1 macrophages (IL-1β production) and less M2 macrophages (IL-10 secretion).Citation43 The balance of M1 vs. M2 cells may be shifted toward classical M1 macrophages in endometriosis. Therefore, pro-inflammatory processes are associated with M1 cell activation. Other investigators, however, showed that a polarization in favor of M2 macrophages was seen in endometriosis, which displays anti-inflammatory characteristics.Citation36 In summary, conflicting evidence for macrophage polarization in endometriosis has been reported.
Taken together, these findings indicate that endometriosis may be associated with increased oxidative stress. Compared with eutopic endometrium, endometriosis exhibits a differential regulation of hemoglobin, heme, and iron metabolism. Heme/HO-1 system and its related effector molecules in endometriotic cells and M2 macrophages are tightly involved in the regulation of the development of endometriosis, with respect to diverse responses to cellular oxidative stress, inflammation, apoptosis, and cytoprotection. Therefore, excessive oxidative stress is a potent inducer of cell toxicity associated with apoptosis or cell death, while exposure of cells to sublethal oxidative stress results in an adaptive, cytoprotective modulation of various survival signaling pathways.
The prooxidant–antioxidant balance in endometriosis-associated ovarian cancer
It has been proposed that ovarian cancers develop from endometriosis.Citation45 Iron overload has been linked to oxidative DNA damage, genomic instability, and carcinogenesis, as seen in hereditary hemochromatosis, viral hepatitis B and C, exposure to asbestos, as well as endometriosis.Citation10,Citation46 Excess heme is also potently associated with carcinogenesis.Citation47 High heme content in red meat is epidemiologically associated with the risks of cancers such as colon, gastrointestinal, pancreatic, lung, and endometrial cancers, as well as several common diseases, including heart failure and diabetes.Citation47
We focus finally on the impact of the microenvironment in the pathogenesis of endometriosis carcinogenesis (Fig. ). Iron and resultant oxidative stress closely correlate with the progress of endometriosis-associated ovarian cancer.Citation2 In response to oxidative stress, HO-1 and Nrf2 upregulate antioxidant and cellular protective genes.Citation20,Citation21 Cancer cells also express increased levels of antioxidant proteins, which detoxify ROS. Nrf2 acts as an essential master switch of cellular defense against various endogenous and exogenous insults, including oxidative stress.Citation20,Citation21 A growing body of evidence indicates that HO-1 and Nrf2 are upregulated in a variety of human cancers and act as players in tumor progression and aggressiveness.Citation48 HO-1 overexpression may facilitate the remodeling of the tumor microenvironment, making it advantageous for cancer progression.Citation21 Taken together, patients with ovarian cancer have less oxidative stress than those with endometriosis. A fine-tuned regulation of lethal and sublethal oxidative stress responses might modulate either cell death or tumor initiation, respectively, due to the iron-induced DNA damage, mutations, and genomic instability.Citation49 An imbalance in cellular oxidant/antioxidant status, orchestrating reduced oxidants and increased antioxidants, can promote the progression of ovarian cancer.Citation13 Primarily, HO-1 and Nrf2 function to protect nonmalignant cells from oxidative stress through upregulation of detoxifying and antioxidant enzymes. Also, antioxidant defense genes protect malignant cells from oxidative stress. Endometriosis is regarded as a potential model system to address the liaison between Hb-, heme-, and iron-induced chronic oxidative stress and cancer development.
Macrophages exhibit both immunostimulatory and immunoregulatory activities upon exposure to oxidative stress including iron.Citation50 Macrophage-related inflammation releases pro-inflammatory cytokines and also increases antioxidant defense mechanisms.Citation51 As described in the previous section, the backup protection systems include Hp–CD163–HO-1 and Hx–CD91–HO-1 pathways in tumor-associated macrophages.Citation10,Citation14 Hemoglobin initially induces macrophage polarization to the M1 phenotype, which is associated with the release of pro-inflammatory cytokines involved in carcinogenesis.Citation52,Citation53 Subsequently, activation of macrophages can reprogram them to acquire the IL-4-induced M2 phenotype characteristics.Citation54 M2 macrophages present in tissue could be modulated by hemoglobin derivatives generating protumoral signals in this microenvironment.Citation55 The activation state of the polarized M2 subtype of macrophages also secretes immunosuppressive factors and an increased amount of antioxidants.Citation55 In response to the oxidative and inflammatory microenvironment, endometriotic cells and macrophages have evolved multiple mechanisms for protection against cell death and, subsequently, initiation of carcinogenesis. Macrophages change their phenotypes to tumor-associated M2 macrophages in the early stages of carcinogenesis.Citation55
These data allow us to speculate that in cancer evolution there are at least two steps to reach cancer (Fig. ). The first step: the generation of oxidative stress-induced DNA damage evokes enhanced cell apoptosis or survival in endometriotic cells, revealing a functional dichotomy for oxidative stress. The second step: a possible carcinogenic approach may establish a status characterized by the maintenance of antioxidant homeostasis against local chronic oxidative stress, with the prospect of cancer progression in that site.
Discussion
Endometriosis may be caused by seeding of regurgitated endometrial cells. Repeated hemorrhage and hemolysis occur during endometriosis development. Extracellular hemoglobin, heme, and iron are abundantly present in the endometriotic cyst fluid or peritoneal fluid during menstruation. The continuous autoxidation of hemoglobin produces superoxide, indicating the importance of hemoglobin redox reactions.Citation56 Iron can catalytically amplify the production of excess ROS via Fenton chemistry.Citation57 Increased heme or iron exposure is associated with endometriotic cell death, mainly due to its prooxidant activity, which can lead to oxidative DNA damage.
Furthermore, hemoglobin, heme, and iron derivatives help to fine-tune the expression of several genes associated with oxidants and antioxidants. These compounds can lead to high levels of Nrf2 and HO-1 expression.Citation20,Citation21 Nrf2 and HO-1 can counteract oxidative damage and inflammation, which in turn induces malignant transformation of endometriotic cells with persistent DNA damage.Citation50 Indeed, these antioxidants are overexpressed in endometriosis-associated ovarian cancer.Citation34 Thus, cancer progression might be associated with an increased level of the antioxidant defense system capacity.
In conclusion, this review has focused particularly on the double-edged sword of redox imbalance. Specific molecules associated with oxidative stress signaling in endometriosis have implications for the spatial and temporal proceeding of biochemical and immunological processes that contribute to inflammation. Continued imbalances in redox homeostasis, especially an increase in oxidative stress, lead to crucial failures of control defense mechanisms, thus inducing cell death. Regulatory mechanisms provided by antioxidant defense signaling events in a subset of women with endometriosis guarantee cell survival processes, thus promoting the development of pathological conditions such as endometriosis carcinogenesis. An increased amount of antioxidant defense may rather lead to malignant transformation and, consequently, cancer progression.
Disclaimer statements
Contributors None.
Funding The present review was supported by grant-in-aid for Scientific Research from the Ministry of Education, Science, and Culture of Japan to the Department of Obstetrics and Gynecology, Nara Medical University (to H.K.).
Conflicts of interest No potential conflicts of interest relevant to this article were reported.
Ethics approval None.
References
- Kobayashi H, Sumimoto K, Moniwa N, Imai M, Takakura K, Kuromaki T, et al. Risk of developing ovarian cancer among women with ovarian endometrioma: a cohort study in Shizuoka, Japan. Int J Gynecol Cancer 2007;17:37–43. doi: 10.1111/j.1525-1438.2006.00754.x
- Yamaguchi K, Mandai M, Toyokuni S, Hamanishi J, Higuchi T, Takakura K, et al. Contents of endometriotic cysts, especially the high concentration of free iron, are a possible cause of carcinogenesis in the cysts through the iron-induced persistent oxidative stress. Clin Cancer Res 2008;14:32–40. doi: 10.1158/1078-0432.CCR-07-1614
- Akasaka J, Uekuri C, Shigetomi H, Koike M, Kobayashi H. Hepatocyte nuclear factor (HNF)-1β and its physiological importance in endometriosis. Biomed Rep 2013;1:13–7.
- Bokor A, Debrock S, Drijkoningen M, Goossens W, Fülöp V, D'Hooghe T. Quantity and quality of retrograde menstruation: a case–control study. Reprod Biol Endocrinol 2009;7:123. doi: 10.1186/1477-7827-7-123
- Nagy E, Eaton JW, Jeney V, Soares MP, Varga Z, Galajda Z, et al. Red cells, hemoglobin, heme, iron, and atherogenesis. Arterioscler Thromb Vasc Biol 2010;30:1347–53. doi: 10.1161/ATVBAHA.110.206433
- Sharpe-Timms KL, Ricke EA, Piva M, Horowitz GM. Differential expression and localization of de-novo synthesized endometriotic haptoglobin in endometrium and endometriotic lesions. Hum Reprod 2000;15:2180–5. doi: 10.1093/humrep/15.10.2180
- Kobayashi H, Imanaka S, Nakamura H, Tsuji A. Understanding the role of epigenomic, genomic and genetic alterations in the development of endometriosis (review). Mol Med Rep 2014;9:1483–505.
- Yoshimoto C, Iwabuchi T, Shigetomi H, Kobayashi H. Cyst fluid iron-related compounds as useful markers to distinguish malignant transformation from benign endometriotic cysts. Cancer Biomark 2015;15:493–9. doi: 10.3233/CBM-150484
- Shigetomi H, Higashiura Y, Kajihara H, Kobayashi H. A potential link of oxidative stress and cell cycle regulation for development of endometriosis. Gynecol Endocrinol 2012;28:897–902. doi: 10.3109/09513590.2012.683071
- Fonseca-Nunes A, Jakszyn P, Agudo A. Iron and cancer risk – a systematic review and meta-analysis of the epidemiological evidence. Cancer Epidemiol Biomarkers Prev 2014;23:12–31. doi: 10.1158/1055-9965.EPI-13-0733
- Thomsen JH, Etzerodt A, Svendsen P, Moestrup SK. The haptoglobin–CD163–heme oxygenase-1 pathway for hemoglobin scavenging. Oxid Med Cell Longev 2013;2013:523652. doi: 10.1155/2013/523652
- Lo SS, Lin SC, Wu CW, Chen JH, Yeh WI, Chung MY, et al. Heme oxygenase-1 gene promoter polymorphism is associated with risk of gastric adenocarcinoma and lymphovascular tumor invasion. Ann Surg Oncol 2007;14:2250–6. doi: 10.1245/s10434-006-9290-7
- Glasauer A, Chandel NS. Targeting antioxidants for cancer therapy. Biochem Pharmacol 2014;92:90–101. doi: 10.1016/j.bcp.2014.07.017
- Etzerodt A, Moestrup SK. CD163 and inflammation: biological, diagnostic, and therapeutic aspects. Antioxid Redox Signal 2013;18:2352–63. doi: 10.1089/ars.2012.4834
- Philippidis P, Mason JC, Evans BJ, Nadra I, Taylor KM, Haskard DO, et al. Hemoglobin scavenger receptor CD163 mediates interleukin-10 release and heme oxygenase-1 synthesis: antiinflammatory monocyte-macrophage responses in vitro, in resolving skin blisters in vivo, and after cardiopulmonary bypass surgery. Circ Res 2004;94:119–26. doi: 10.1161/01.RES.0000109414.78907.F9
- Schaer DJ, Alayash AI, Buehler PW. Gating the radical hemoglobin to macrophages: the anti-inflammatory role of CD163, a scavenger receptor. Antioxid Redox Signal 2007;9:991–9. doi: 10.1089/ars.2007.1576
- Lanceta L, Li C, Choi AM, Eaton JW. Haem oxygenase-1 overexpression alters intracellular iron distribution. Biochem J 2013;449:189–94. doi: 10.1042/BJ20120936
- Chiabrando D, Vinchi F, Fiorito V, Mercurio S, Tolosano E. Heme in pathophysiology: a matter of scavenging, metabolism and trafficking across cell membranes. Front Pharmacol 2014;5:61. doi: 10.3389/fphar.2014.00061
- Matsuzaki S, Darcha C. Adenosine triphosphate-binding cassette transporter G2 expression in endometriosis and in endometrium from patients with and without endometriosis. Fertil Steril 2012;98:1512–20.e3. doi: 10.1016/j.fertnstert.2012.07.1133
- Lee JM, Li J, Johnson DA, Stein TD, Kraft AD, Calkins MJ, et al. Nrf2, a multi-organ protector? FASEB J 2005;19:1061–6. doi: 10.1096/fj.04-2591hyp
- Na HK, Surh YJ. Oncogenic potential of Nrf2 and its principal target protein heme oxygenase-1. Free Radic Biol Med 2014;67:353–65. doi: 10.1016/j.freeradbiomed.2013.10.819
- Kobayashi H, Yamada Y, Kanayama S, Furukawa N, Noguchi T, Haruta S, et al. The role of iron in the pathogenesis of endometriosis. Gynecol Endocrinol 2009;25:39–52. doi: 10.1080/09513590802366204
- Kobayashi H, Kajiwara H, Kanayama S, Yamada Y, Furukawa N, Noguchi T, et al. Molecular pathogenesis of endometriosis-associated clear cell carcinoma of the ovary (review). Oncol Rep 2009;22:233–40. doi: 10.3892/or_00000417
- Hsieh YP, Huang CH, Lee CY, Lin CY, Chang CC. Silencing of hepcidin enforces the apoptosis in iron-induced human cardiomyocytes. J Occup Med Toxicol 2014;9:11. doi: 10.1186/1745-6673-9-11
- Turgut A, Özler A, Görük NY, Tunc SY, Evliyaoglu O, Gül T. Copper, ceruloplasmin and oxidative stress in patients with advanced-stage endometriosis. Eur Rev Med Pharmacol Sci 2013;17:1472–8.
- Kajitani T, Maruyama T, Asada H, Uchida H, Oda H, Uchida S, et al. Possible involvement of nerve growth factor in dysmenorrhea and dyspareunia associated with endometriosis. Endocr J 2013;60:1155–64. doi: 10.1507/endocrj.EJ13-0027
- Kobayashi H, Yamada Y, Morioka S, Niiro E, Shigemitsu A, Ito F. Mechanism of pain generation for endometriosis-associated pelvic pain. Arch Gynecol Obstet 2014;289:13–21. doi: 10.1007/s00404-013-3049-8
- Sengupta A, Hon T, Zhang L. Heme deficiency suppresses the expression of key neuronal genes and causes neuronal cell death. Brain Res Mol Brain Res 2005;137:23–30. doi: 10.1016/j.molbrainres.2005.02.007
- Van Langendonckt A, Casanas-Roux F, Dolmans MM, Donnez J. Potential involvement of hemoglobin and heme in the pathogenesis of peritoneal endometriosis. Fertil Steril 2002;77:561–70. doi: 10.1016/S0015-0282(01)03211-3
- Prieto L, Quesada JF, Cambero O, Pacheco A, Pellicer A, Codoceo R, et al. Analysis of follicular fluid and serum markers of oxidative stress in women with infertility related to endometriosis. Fertil Steril 2012;98:126–30. doi: 10.1016/j.fertnstert.2012.03.052
- Ferrero S, Gillott DJ, Remorgida V, Anserini P, Price K, Ragni N, et al. Haptoglobin beta chain isoforms in the plasma and peritoneal fluid of women with endometriosis. Fertil Steril 2005;83:1536–43. doi: 10.1016/j.fertnstert.2004.11.064
- Sharpe-Timms KL, Nabli H, Zimmer RL, Birt JA, Davis JW. Inflammatory cytokines differentially up-regulate human endometrial haptoglobin production in women with endometriosis. Hum Reprod 2010;25:1241–50. doi: 10.1093/humrep/deq032
- Sharpe-Timms KL, Zimmer RL, Ricke EA, Piva M, Horowitz GM. Endometriotic haptoglobin binds to peritoneal macrophages and alters their function in women with endometriosis. Fertil Steril 2002;78:810–9. doi: 10.1016/S0015-0282(02)03317-4
- Hwang JH, Lee KS, Joo JK, Wang T, Son JB, Park JH, et al. Identification of biomarkers for endometriosis in plasma from patients with endometriosis using a proteomics approach. Mol Med Rep 2014;10:725–30.
- Cominelli A, Gaide Chevronnay HP, Lemoine P, Courtoy PJ, Marbaix E, Henriet P. Matrix metalloproteinase-27 is expressed in CD163+/CD206+ M2 macrophages in the cycling human endometrium and in superficial endometriotic lesions. Mol Hum Reprod 2014;20:767–75. doi: 10.1093/molehr/gau034
- Bacci M, Capobianco A, Monno A, Cottone L, Di Puppo F, Camisa B, et al. Macrophages are alternatively activated in patients with endometriosis and required for growth and vascularization of lesions in a mouse model of disease. Am J Pathol 2009;175:547–56. doi: 10.2353/ajpath.2009.081011
- Wölfler MM, Meinhold-Heerlein IM, Henkel C, Rath W, Neulen J, Maass N, et al. Reduced hemopexin levels in peritoneal fluid of patients with endometriosis. Fertil Steril 2013;100:777–81. doi: 10.1016/j.fertnstert.2013.05.010
- Lousse JC, Defrère S, Van Langendonckt A, Gras J, González-Ramos R, Colette S, et al. Iron storage is significantly increased in peritoneal macrophages of endometriosis patients and correlates with iron overload in peritoneal fluid. Fertil Steril 2009;91:1668–75. doi: 10.1016/j.fertnstert.2008.02.103
- Worthen CA, Enns CA. The role of hepatic transferrin receptor 2 in the regulation of iron homeostasis in the body. Front Pharmacol 2014;5:34. doi: 10.3389/fphar.2014.00034
- Wilkinson N, Pantopoulos K. The IRP/IRE system in vivo: insights from mouse models. Front Pharmacol 2014;5:176. doi: 10.3389/fphar.2014.00176
- Kobayashi H, Yamashita Y, Iwase A, Yoshikawa Y, Yasui H, Kawai Y, et al. The ferroimmunomodulatory role of ectopic endometriotic stromal cells in ovarian endometriosis. Fertil Steril 2012;98:415–22. e1–12. doi: 10.1016/j.fertnstert.2012.04.047
- Kobayashi H, Higashiura Y, Shigetomi H, Kajihara H. Pathogenesis of endometriosis: the role of initial infection and subsequent sterile inflammation. Mol Med Rep 2014;9:9–15.
- Beste MT, Pfäffle-Doyle N, Prentice EA, Morris SN, Lauffenburger DA, Isaacson KB, et al. Molecular network analysis of endometriosis reveals a role for c-Jun-regulated macrophage activation. Sci Transl Med 2014;6(222):222ra16. doi: 10.1126/scitranslmed.3007988
- Xu L, Yang F, Lin R, Han C, Liu J, Ding Z. Induction of m2 polarization in primary culture liver macrophages from rats with acute pancreatitis. PLoS One 2014;9(9):e108014. doi: 10.1371/journal.pone.0108014
- Lim D, Oliva E. Precursors and pathogenesis of ovarian carcinoma. Pathology 2013;45:229–42. doi: 10.1097/PAT.0b013e32835f2264
- Toyokuni S. Iron as a target of chemoprevention for longevity in humans. Free Radic Res 2011;45:906–17. doi: 10.3109/10715762.2011.564170
- Hooda J, Shah A, Zhang L. Heme, an essential nutrient from dietary proteins, critically impacts diverse physiological and pathological processes. Nutrients 2014;6:1080–102. doi: 10.3390/nu6031080
- Jozkowicz A, Was H, Dulak J. Heme oxygenase-1 in tumors: is it a false friend? Antioxid Redox Signal 2007;9:2099–117. doi: 10.1089/ars.2007.1659
- Torti SV, Torti FM. Iron and cancer: more ore to be mined. Nat Rev Cancer 2013;13:342–55. doi: 10.1038/nrc3495
- Xu X, Li H, Hou X, Li D, He S, Wan C, et al. Punicalagin induces Nrf2/HO-1 expression via upregulation of PI3K/AKT pathway and inhibits LPS-induced oxidative stress in RAW264.7 macrophages. Mediators Inflamm 2015;2015:380218.
- Gostner JM, Becker K, Fuchs D, Sucher R. Redox regulation of the immune response. Redox Rep 2013;18:88–94. doi: 10.1179/1351000213Y.0000000044
- Gleissner CA. Macrophage phenotype modulation by CXCL4 in atherosclerosis. Front Physiol 2012;3:1. doi: 10.3389/fphys.2012.00001
- Young VJ, Brown JK, Saunders PT, Horne AW. The role of the peritoneum in the pathogenesis of endometriosis. Hum Reprod Update 2013;19:558–69. doi: 10.1093/humupd/dmt024
- Itoh F, Komohara Y, Takaishi K, Honda R, Tashiro H, Kyo S, et al. Possible involvement of signal transducer and activator of transcription-3 in cell-cell interactions of peritoneal macrophages and endometrial stromal cells in human endometriosis. Fertil Steril 2013;99:1705–13. doi: 10.1016/j.fertnstert.2013.01.133
- Thanee M, Loilome W, Techasen A, Namwat N, Boonmars T, Pairojkul C, et al. Quantitative changes in tumor-associated M2 macrophages characterize cholangiocarcinoma and their association with metastasis. Asian Pac J Cancer Prev 2015;16:3043–50. doi: 10.7314/APJCP.2015.16.7.3043
- Winterbourn CC. Free-radical production and oxidative reactions of hemoglobin. Environ Health Perspect 1985;64:321–30. doi: 10.1289/ehp.8564321
- Patel M, Ramavataram DV. Non transferrin bound iron: nature, manifestations and analytical approaches for estimation. Indian J Clin Biochem 2012;27:322–32. doi: 10.1007/s12291-012-0250-7