Abstract
Atherosclerosis is a leading cause of coronary heart disease and stroke. Since 1981, more than 980,000 cases of AIDS have been reported in the United States. According to the Centers for Disease Control, more than 1 million Americans may be infected with HIV. By killing or damaging CD4+ T cells of the body’s immune system, HIV progressively destroys the body’s ability to fight infections. People diagnosed with AIDS often suffer from life-threatening diseases caused by opportunistic infections such as tuberculosis. HIV-infected individuals have increased risks for atherosclerosis. This review summarizes the effects of oxidized low density lipoproteins in impairing macrophage functions in individuals with atherosclerosis (with and without HIV infection) thereby enhancing the susceptibility to Mycobacterium tuberculosis infection.
Keywords:
Introduction
Oxidized low-density lipoprotein (ox-LDL) has been shown in several studies to be an independent marker of the progression of atherosclerosis.Citation1–Citation4 The pathophysiology relates to macrophage ingestion of excess ox-LDL and the formation of foam cells, the acknowledged trigger of atherosclerosis.Citation5 It has been shown that both high-density lipoprotein (HDL) and low-density lipoprotein (LDL) cholesterol contain the antioxidant enzyme glutathione peroxidase embedded in the lipoprotein, and a continuous supply of glutathione (GSH) is needed to prevent the oxidation of HDL and LDL cholesterol.Citation6 The observation that excessive ox-LDL ingestion by macrophage interferes with normal macrophage function may have implications in other disease conditions in which low GSH plays a role, such as HIV infection. Individuals with HIV infection are at increased risk for the progression of atherosclerosisCitation7 as well as infectious diseases such as tuberculosis (TB).Citation8 This review postulates that the compromise in macrophage function that occurs with HIV may increase the risk of both atherosclerosis as well as susceptibility to Mycobacterium tuberculosis (M. tb) infection. The increase in ox-LDL ingestion and resulting macrophage dysfunction may contribute to the increased risk for M. tb infection in both HIV-positive and HIV-negative individuals.
Atherosclerosis
Atherosclerosis is a leading cause of coronary heart disease and stroke, which were responsible for more than 589,000 deaths in 2005 – almost 25% of all deaths in the United States. Each year more than 1.2 million will suffer a coronary attack and almost 800,000 people will suffer a stroke. Atherosclerosis is a process in which deposits of fatty substances, cholesterol, cellular waste products, calcium and other substances build up in the inner lining of an artery. This build-up is called plaque.Citation7 It usually affects large- and medium-sized arteries. Some hardening of arteries often occurs when people grow older. Plaques can grow large enough to significantly reduce the blood’s flow through an artery. But most of the damage occurs when they become fragile and rupture. Plaques that rupture cause blood clots to form that can block blood flow or break off and travel to another part of the body. If either happens and blocks a blood vessel that feeds the heart, it causes a heart attack. And if blood supply to the arms or legs is reduced, it can cause difficulty walking and eventually lead to gangrene. Atherosclerosis can result in myocardial infarction, and bits of plaque can lodge in arteries in the brain, causing a stroke.Citation7
Glutathione
GSH is a tripeptide that, in its reduced form, protects cells against oxidizing agents, free radicals and reactive oxygen intermediates (ROI). In addition to its antioxidant role, GSH plays a vital role in maintenance of cell viability and regulating immune cell functions.Citation9 Synthesis of GSH occurs in 2 steps. The initial step (the rate limiting step) is the formation of a dipeptide, γ-glutamyl cysteine, a reaction that is catalyzed by γ-glutamylcysteine synthetase. Intracellular levels of L-cysteine are substantially lower than levels of L-glutamate and glycine. Therefore, GSH synthesis is limited by the availability of cysteine.Citation10 The second step involved in the synthesis of GSH is the formation of γ-glutamyl cysteine glycine, catalyzed by the enzyme GSH synthetase.Citation10 Intracellular GSH levels can be increased or decreased by treatment with N-acetyl cysteine (NAC) or buthionine sulfoximine (BSO), respectively. The most efficient way to increase the levels of cysteine in cells grown in vitro is to supply the culture medium with NAC, which is easily taken up by the cells and is nontoxic. Intracellularly, NAC is de-acetylated and cysteine is utilized for GSH synthesis. BSO specifically inhibits the activity of the γ-glutamyl-cysteinyl synthetase enzyme, which catalyzes the first step reaction in the synthesis of GSH,Citation9,Citation10 leading to inhibition in the synthesis of GSH.
Oxidized LDL
Oxidation of LDL (ox-LDL) cholesterol has been shown to convert LDL cholesterol to a form that is recognized by scavenger receptors on macrophages and to contribute to foam cell formation.Citation6 It has been known for some time that vitamin E and β-carotene are found in the LDL complex and that decreases in vitamin E and β-carotene are early events reflecting the initial stages of lipid peroxidation formation. Citation11 It has only recently been shown that glutathione peroxidase (GPx) is also found to occur naturally in the LDL lipoprotein. Substituting the unique substrate for GPx, reduced GSH, with liposomal GSH can slow the formation of ox-LDL in vitro in human blood and slow atherosclerosis in vivo in the ApoE(−/−) mouse model of atherosclerosis.Citation12
Ox-LDL and atherosclerosis
Ox-LDL has been shown in several studies to be an independent marker of the progression of atherosclerosis.Citation1–Citation4 The pathophysiology relates to macrophage ingestion of excess ox-LDL and the formation of foam cells, the acknowledged trigger of atherosclerosis.Citation5 The mechanism of ox-LDL and the requirement for GSH to prevent oxidation of LDL cholesterol has been described in Rosenblat et alCitation12 using liposomal GSH as the source of GSH in both human blood, in vitro studies and in the mouse model of atherosclerosis. This study demonstrated that both LDL and HDL contain the antioxidant enzyme GSH peroxidase embedded in the lipoprotein.Citation12 As GSH is the single substrate for GSH peroxidase, a continuous supply of GSH in the reduced state is needed to prevent the oxidation of LDL and HDL cholesterol. This study also demonstrated that preventing the oxidation of LDL and increasing the intracellular GSH levels is able to maintain normal macrophage function. It is likely that the presence of excess amounts of ox-LDL will be an indicator of general macrophage dysfunctions as it has been shown to stimulate the release of macrophage inflammatory protein 1-alpha (MIP-1alpha)Citation5 as well as other immune responses. Ox-LDL is known to be ingested by macrophages and without adequate antioxidant function is prone to form foam cells (), a transition which is also a contributing factor to the formation of atherosclerosisCitation14 and atherosclerotic lesions.Citation13–Citation15
Figure 1 Receptor mediated uptake of oxidized low density lipoproteins by macrophages resulting in foam cell formation.
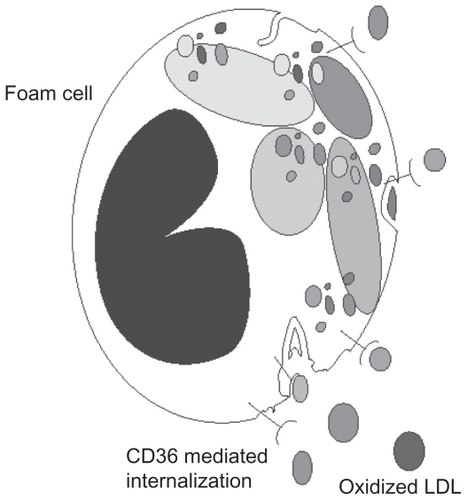
Macrophages and atherosclerosis
Macrophages are derived from the circulating pool of monocytes. Citation16 Monocytes are produced in the bone marrow and, in the absence of specific survival signals, are programmed to undergo apoptosis in 24–48 hours.Citation16,Citation17 When monocytes are recruited into local environments they differentiate into macrophages. In arteries, macrophage scavenging function may be diverted to the formation of plaque; in the lung they become alveolar macrophages. In atherosclerosis, macrophage oxidative impairment occurs in the GSH-related antioxidant system,Citation5 probably related to the unregulated ingestion of ox-LDL via the CD36 receptor.Citation18,Citation19
Role of GSH in macrophage function
In the natural state, GSH levels are maintained by both the synthesis of GSH and by GSH reductase, an enzyme that catalyzes the reduction of glutathione disulfide (GSSG) to GSH, with NADPH as the reducing cofactor. Increasing the expression of GSH reductase, the enzyme which returns GSH to the biochemically reduced (functional) state, decreases atherosclerotic lesions.Citation20 It has been shown that lymphocyte proliferation after mitogenic lectin exposure is decreasedCitation21 with low intracellular GSH.Citation21,Citation22
The level of reduced GSH in the extracellular lung fluid (ELF) or alveolar lining fluid has been estimated to be as much as 140-fold higher than the level of reduced GSH found in the plasma of the same individuals.Citation23 The antioxidant GSH is essential for the detoxification of endogenous and exogenous oxidant radicals and protection of cells residing in the airway and alveolusCitation24,Citation25 and during oxidative stress such as cigarette smoking.Citation26 In disease states, the level of ELF GSH may decrease dramatically to the point that acute respiratory distress syndrome has only 0.05% of the amount found in the normal ELF.Citation27
In the lung, intracellular antioxidants are expressed at a relatively low level in the lung epithelial tissue and are not induced by oxidative stress. The major antioxidants in the lung are extracellular and the GSH system is the major antioxidant mechanism used in the airways.Citation28 Alveolar macrophage cells rely on the ELF level of GSH to provide the biochemical support to maintain intracellular GSH levelsCitation29 and protect their membranes during the respiratory burst.Citation30
When macrophage GSH availability is limited, cellular functions such as phagocytosis and microbial clearance are compromised.Citation24 The mouse model of decreased alveolar macrophage GSH resulted in a decrease in the rate of phagocytosis of individual macrophages as well as a decrease in the overall percentage of macrophages positive for phagocytosis. Citation24 Similar findings have been reported in the alveolar function of guinea pigs exposed to ethanol in utero resulting in low macrophage GSH. The addition of precursors of GSH to the diet of the ethanol-exposed animals increased the GSH levels in the ELF and alveolar macrophages and maintained both the rate of internalization and the number of macrophages positive for internalization.Citation24 It was also shown that decreased GSH availability in the ELF and alveolar macrophages of ethanol-fed mice resulted in an increase of apoptosis of macrophages that could be reversed by maintaining GSH availability.Citation24
GSH and arterial macrophage functions in atherosclerosis
GSH deficiency in the macrophage has been shown to be associated with compromise of mitochondrial function. GSH plays an integral role in protecting the mitochondria from ROI damage and it is the level of mitochondrial GSH that determines when cellular toxicity commences.Citation18 GSH deficiency in mitochondria contributes to macrophage dysfunction in atherosclerosis. Data from a study of the upregulation of GSH reductase in macrophages provide direct evidence that the GSH-dependent anti-oxidant system in macrophages plays a critical role in atherogenesis.Citation20 The study shows that overexpression of GSH in either mitochondria or the cytosol of macrophages decreases the severity of atherosclerosis in this animal model.Citation20
Ox-LDL-related peroxyl radical formation is implicated in both mitochondrial dysfunction and macrophage lysis.Citation19 Ox-LDL toxicity is related to compromise of mitochondrial function including an increase in mitochondrial Ca+2, opening of mitochondrial permeability transition pores and depolarization of mitochondrial membrane potential.
It has been observed that maintaining adequate GSH levels in macrophage will prevent the toxicity of ox-LDL in an in vitro macrophage model.Citation31–Citation33 These findings help explain why a decreased amount of reduced GSH in blood has been shown to be an independent marker of progression of atherosclerosis in 2 studies.Citation13,Citation14
HIV
Worldwide, it is estimated that approximately 40 million people are infected with HIV, two-thirds of whom live in sub-Saharan Africa. Three regions, Africa, Asia, and Latin America, have the highest rates of new infections. AIDS is the fourth leading cause of death worldwide.Citation34 HIV is the cause of AIDS. Both HIV-1 and HIV-2 cause AIDS, but HIV-1 is found worldwide, whereas HIV-2 is found primarily in West Africa. HIV preferentially infects and kills CD4+ T lymphocytes, resulting in the loss of cell-mediated immunity and a high probability that the host will develop opportunistic infections. HIV-infected individuals have an increased risk for atherosclerosis and susceptibility to M. tb infection.Citation35
HIV and atherosclerosis
As people with HIV live longer due to effective combination antiretroviral therapy (ART), cardiovascular disease has become an increasingly important cause of morbidity and mortality. But it remains controversial whether HIV infection contributes to accelerated atherosclerosis independent of traditional cardiovascular risk factors. In a recent cross-sectional study, more than 400 HIV-positive participants without pre-existing cardiovascular disease in the FRAM (Fat Redistribution and Metabolic Change in HIV Infection) study were compared with HIV-negative participants in the MESA (Multi-Ethnic Study of Atherosclerosis) cohort.Citation7 The preclinical atherosclerosis was assessed by measuring carotid intima-media thickness (IMT), or thickness of the walls of the arteries in the neck that supply the brain. IMT was evaluated at 2 sites in the artery, known as the internal/ bulb. Even after adjustment for traditional cardiovascular disease risk factors, HIV infection was accompanied by more extensive atherosclerosis as measured by IMT.Citation7 The association of HIV infection with IMT was similar to that of traditional cardiovascular disease risk factors, such as smoking. The impact of HIV infection on preclinical atherosclerosis overrides any small differences related to antiretroviral therapy or specific drug classes. The effect of HIV is so big that no drug or class of drugs stands out as being an effective contributor.Citation7
GSH and HIV-infection
Findings from our research laboratory as well as by other groups confirm that intracellular levels of GSH are decreased in patients with AIDS,Citation35 in whom the risk of TB is many times that of healthy individuals.Citation35 The factors responsible for low GSH in HIV infection are poorly understood. Infection with HIV is believed to trigger a range of metabolic changes in addition to the progressive deficits in cellular immunity and increased susceptibility to opportunistic infections that are its clinical hallmarks, and the progression to AIDS.Citation36–Citation45 The decreased GSH content in immune cells of HIV-positive individuals was at least in part attributed to the decrease in plasma cysteine and increased plasma glutamate (an inhibitor of cysteine permeation via the Xc-transport system), as observed during early infection (). The decreased intracellular GSH and plasma cysteine observed in HIV patients may also be due to chronic oxidative stress and this may lead to the progression of the disease. The decreased availability of cysteine can be overcome to some extent by the cysteine precursor NAC.Citation42 Herzenberg and colleagues found that NAC treatment improves the clinical situation and delays the disease progression.Citation43 This study showed that long-term NAC administration to AIDS patients improves their hematological profile, GSH content and life expectancy.Citation43
Figure 2 Possible causes for decreased intracellular levels of glutathione in individuals with HIV and/or M. tb infection.
Ox-LDL and HIV
Increased incidence of cardiovascular events in HIV patients has sparked interest in elucidating the molecular mechanism behind endothelial dysfunction resulting in atherosclerotic pathology. While there is no evidence within the literature of a molecular mechanism responsible for increased cardiovascular risk within such HIV+ cohorts, animal models expressing transgenic HIV-1 offer a promising insight. One such study has shown significantly increased aortic arch lectin- like oxidized-low-density-lipoprotein receptor-1 (LOX-1) gene expression in HIV-1 transgenic rats compared to control rats.Citation46 LOX-1 is an endothelial receptor for ox-LDL and is hypothesized to be an early marker of endothelial dysfunction. It has been proposed that endothelial dysfunction is the first step in progression towards atherosclerotic plaque formation. This same group has also demonstrated significantly increased expression of inflammatory endothelial adhesion molecules in the same HIV-1 model. Both inducible vascular cell adhesion molecule- 1 (VCAM-1) and constitutively expressed intercellular adhesion molecule- 1 (ICAM-1) were both significantly elevated in HIV-1 transgenic rats compared to control rats, strongly suggesting for the first time that the HIV infection, independent of antiretroviral therapy- associated dyslipidemia, is capable of pathologic endothelial dysfunction that may prelude atherosclerotic plaque formation in major blood vessels.Citation46
Tuberculosis
TB is the most prevalent infectious disease in the world. Citation47–Citation54 In recent years there has been a significant increase in the incidence of TB due to the emergence of multi-drug resistant strains of M. tb and the increased numbers of highly susceptible immuno-compromised individuals arising from the AIDS pandemic.Citation8 It is also believed that in developing countries, as many as 40%–80% of individuals with AIDS are at risk of developing TB.Citation8,Citation35 The innate and adaptive immune systems contribute to host defenses against M. tb infection.Citation48–Citation54 Control of M. tb infection occurs both at the macroscopic and cellular levels. At the macroscopic level, the physical containment of viable mycobacteria within fibrotic granulomas contributes to prevention of overt disease. Citation49–Citation50 Control of mycobacterial replication also occurs within macrophages. Resident tissue macrophages provide the first-line defense against M. tb infection. Macrophages and natural killer (NK) cells play an important role in innate defense against M. tb infection.Citation51–Citation53
Tuberculosis and GSH
Our group reported that GSH plays a key role in limiting intracellular growth of Mycobacterium bovis BCG in nitric oxide (NO)-deficient macrophages, such as macrophages derived from inducible nitric oxide synthase (NOS2)-knock out mice and human peripheral blood monocyte-derived macrophages (HMDM). Thus, GSH has direct antimycobacterial activity distinct from its role as an NO carrier, functioning as an effector molecule in innate defense against M. tb infection.Citation47,Citation51,Citation52 These results unfold a novel and potentially important innate defense mechanism adopted by human macrophages to control M. tb infection.Citation51,Citation52 Consistent with these observations, we have also found that GSH mediates growth control of virulent M. tb in human blood cultures.Citation35,Citation53 These results indicate that the inability of immune cells to contain M. tb growth may be a consequence of the inability of the immune cells to maintain adequate GSH levels during in vitro infection.Citation35 Furthermore, our recent studies indicate that GSH in combination with cytokines such as interleukin (IL-2) and IL-12, activate NK cells to control M. tb infection. Citation53 Most importantly, our recent studies demonstrate that GSH levels are significantly reduced in peripheral blood mononuclear cells and red blood cells isolated from TB patients and this decrease correlates with increased pro-inflammatory cytokine production and enhanced growth of M. tb.Citation54 Our group was the first to report that GSH levels are decreased in individuals with pulmonary TB and to correlate GSH levels with protective immunity.Citation54 Furthermore, our study provides a direct relationship between decreased GSH levels, pro-inflammatory cytokine production and enhanced growth of M. tb in TB patients.Citation54
ROI, GSH and TB
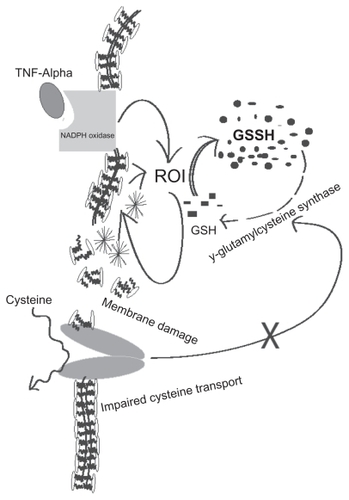
ROI are generated from NADPH through a catalytic reaction of membrane bound enzyme NADPH oxidase, which is activated by assembling cytosolic regulatory components. The most important characteristic of ROI, either in vivo or in vitro, is peroxidation of lipids resulting in tissue damage and death of affected cells. Lipid peroxidation is a chain reaction providing a continuous supply of free radicals that initiate further peroxidation.Citation55 Free radical-induced lipid peroxidation causes marked alterations in the structural integrity and functions of membrane. The lipid peroxides formed at the primary site such as lungs could be transferred through the circulation to other organs and tissues.Citation56–Citation58 Jack et alCitation59 reported that several circulating markers of free radical activity were increased in pulmonary TB patients and some of these markers remain elevated even after completion of antimicrobial chemotherapy, indicating ongoing oxidative stress, which may contribute to decreased GSH levels. It is our prediction that in active TB, increased levels of ROI cause impaired cystine transport leading to decreased levels of GSH ().
TNF-α, ROI and GSH
It has been shown that TNF-α stimulates ROI production. TNF-α could impair GSH-redox status by production of ROI, and impairment of the GSH-reductase system, thereby leading to decreased regeneration of reduced GSH from oxidized GSSG (). Moreover, enhanced ROI is likely to increase TNF-α in various cells, and depletion of reduced GSH will increase the inflammatory response to the cytokine. Nuclear factor-kappa B (NF-κB) is a DNA binding protein and a ROI-sensitive transcription factor for several cytokines, including TNF-α.Citation57 While NF-kB activation (leading to TNF-α production) is induced and enhanced by ROI, it can also be blocked by anti-oxidants such as vitamin E and GSH-enhancing agents such as NAC.Citation57 Excessive production of TNF-α and increased tissue sensitivity to this cytokine has been implicated in the immunopathology of TB, such as caseous necrotizing reactions. For example, excess TNF-α (relative to its receptors) in human bronchoalveolar lavage fluid was associated with tissue necrosis and cavity formation.Citation59–Citation61 Systemic spillover of TNF-α may account for unwanted inflammatory effects like fever and wasting, which manifests clinically as cachexia, consistent with the original designation as cachetin.Citation65 In HIV-infected persons, the effects of TNF-α may be particularly deleterious, as this cytokine has been implicated as a stimulus for HIV expression and for activation-induced T cell apoptosis. For these reasons, several studies have examined anti-TNF treatments in persons with AIDS and in HIV+ TB.Citation62–Citation65 Thus enhanced TNF-α production may represent a pathogenic loop, leading to enhanced inflammation and ROI production, leading to reduced GSH levels.
Atherosclerosis and tuberculosis
Atherosclerosis is an inflammatory disease involving the accumulation of macrophages in the intima. Wnt5a is a noncanonical member of the Wnt family of secreted glycoproteins. Recently, human macrophages have been shown to express Wnt5a upon stimulation with bacterial pathogens in vitro and in granulomatous lesions in the lung of M. tb-infected patients. Wnt5a expression has also been linked to Toll-like receptor-4 (TLR-4), an innate immune receptor implicated in atherosclerosis. These observations, along with the fact that Wnt5a is involved in cell migration and proliferation, led Christman et al to postulate that Wnt5a plays a role in atherosclerosis.Citation66 To investigate this hypothesis, Christman et al characterized Wnt5a expression in murine and human atherosclerotic lesions. Tissue sections derived from the aortic sinus to the aortic arch of apolipoprotein E-deficient mice and sections derived from the carotid arteries of patients undergoing endarterectomy were subjected to immunohistochemical analysis. All samples were found to be positive for Wnt5a with predominant staining in the areas of macrophage accumulation within the intima. In parallel, the investigators probed for the presence of TLR-4 and found coincident TLR-4 and Wnt5a expression.Citation66 For both the Wnt5a and TLR-4 staining, consecutive tissue sections treated with an isotype- and species-matched Ig served as a negative control and exhibited little, if any, reactivity. Quantitative RT-PCR revealed that Wnt5a mRNA expression in RAW264.7 murine macrophages can be induced by stimulation with LPS, a known ligand for TLR-4. Combined, these findings demonstrate for the first time Wnt5a expression in human and murine atherosclerotic lesions and suggest that cross-talk between TLR-4 and Wnt5a is operative in atherosclerosis.Citation66
Conclusion
Macrophage functions are compromised by decreased intracellular levels of GSH when an excess of ox-LDL is present. This has been shown to increase the development and progression of atherosclerosis. Reduction of intracellular GSH is a feature of HIV, which is also associated with an increased risk of atherosclerosis. The compromise of innate immune function that occurs with low GSH in the macrophage may also increase susceptibility to infection with M. tb, for which individuals with HIV have an increased risk. It is possible that the compromise of macrophage function related to the ingestion of ox-LDL and the decreased macrophage levels of GSH increase the risk of contracting macrophage-related infectious disease such as M. tb infection in both HIV-positive and HIV-negative individuals. These observations suggest that further investigation into the relationship between low GSH, increased ox-LDL and susceptibility to infection with M. tb is warranted.
Acknowledgment
Corresponding author’s work is supported by American Heart Association (Scientist Development Grant), Potts Memorial Foundation, UMDNJ Foundation grant, Your Energy Systems and start-up funds from Western University of Health Sciences.
Disclosure
The authors declare no conflicts of interest.
References
- GomezMValleVArosFSanzGSalaJFiolMOxidized LDL, lipoprotein (a) and other emergent risk factors in acute myocardial infarction (FORTIAM study)Rev Esp Cardiol200962437338219401122
- NarukoTUedaMEharaSItohAHazeKShiraiNPersistent high levels of plasma oxidized low-densty lipoprotein after acute myocardial infarction predict stent restenosisArterioscler Thromb Vasc Biol200626487788316469945
- MeisingerCBaumertJKhuseyinovaNLoewelHKoenigWPlasma oxidized low-density lipoprotein, a strong predictor for acute coronary heart disease events in apparently healthy, middle-aged men from the general populationCirculation2005112565165716043640
- YamashitaHEharaSYoshiyamaMNarukoTHazeKShiraiNElevated plasma levels of oxidized low-density lipoprotein relate to the presence of angiographically detected complex and thrombotic coronary artery lesion morphology in patients with unstable anginaCirc J200771568168717456991
- PentikainenMOOorniKAla-KorpelaMKovanenPTModified LDL – trigger of atherosclerosis and inflammation in the arterial intimaJ Internal Med2000247335937010762453
- SteinbrecherUPWitztumJLParthasarathySSteinbergDDecrease in reactive amino groups during oxidation or endothelial cell modification of LDL. Correlation with changes in receptor-mediated catabolismArterioscler Thromb Vasc Biol198772135143
- GrunfeldCDelaneyJAWankeCPreclinical atherosclerosis due to HIV infection: carotid intima-medial thickness measurements from the FRAM studyAIDS200923141841184919455012
- HavlirDVGetahunHSanneINunnPOpportunities and challenges for HIV care in overlapping HIV and TB epidemicsJAMA2008300442343018647985
- DenekeSMFanburgBLRegulation of cellular glutathioneAm J Physiol19892574 Pt 1L163L1732572174
- GriffithOWBiologic and pharmacologic regulation of mammalian glutathione synthesisFree Radic Biol Med19999–1092293510569625
- WitztumJLSteinbergDRole of oxidized low density lipoprotein in atherogenesisJ Clin Invest1991886178517921752940
- RosenblatMVolkovaNColemanRAviramMAnti-oxidant and anti-atherogenic properties of liposomal glutathione: studies in vitro, and in the atherosclerotic apolipoprotein E-deficient miceAtherosclerosis20071952e61e6817588583
- FimognariFLScarlataSConteMEIncalziRAMechanisms of atherothrombosis in chronic obstructive pulmonary diseaseInt J Chron Obstruct Pulmon Dis200831899618488431
- CollesSMMaxsonJMCarlsonSGChisolmGMOxidized LDL-induced injury and apoptosis in atherosclerosis. Potential roles for oxysterolsTrends Cardiovasc Med2001113–413113811686002
- BerlinerJATerritoMCSevanianARaminSKimJABamshadBMinimally modified low density lipoprotein stimulates monocyte endothelial interactionsJ Clin Invest1990854126012662318980
- ThomasEDRambergRESaleGESparkesRSGoldeDWDirect evidence for a bone marrow origin of the alveolar macrophage in manScience1976192424310161018775638
- ClineMJLehrerRITerritoMCGoldeDWUCLA Conference. Monocytes and macrophages: functions and diseasesAnn Intern Med19788817888339803
- PingXDHarrisFLBrownLAGauthierTWIn vivo dysfunction of the term alveolar macrophage after in utero ethanol exposureAlcohol Clin Exp Res200731230831617250624
- DengTZhangLGeYLuMZhengXRedistribution of intracellular calcium and its effect on apoptosis in macrophages: Induction by oxidized LDLBiomed Pharmacother200863426727418602792
- QiaoMKisgatiMCholewaJMZhuWSmartEJSulistioMSIncreased expression of glutathione reductase in macrophages decreases atherosclerotic lesion formation in low-density lipoprotein receptor-deficient miceArterioscler Thromb Vasc Biol20072761375138217363688
- HamilosDLZelarneyPMascaliJJLymphocyte proliferation in glutathione-depleted lymphocytes: direct relationship between glutathione availability and the proliferative responseImmunopharmacology19891832232352575086
- MessinaJPLawrenceDACell cycle progression of glutathione-depleted human peripheral blood mononuclear cells is inhibited at S phaseJ Immunol19891436197419812789253
- CantinAMNorthSLHubbardRCCrystalRGNormal alveolar epithelial lining fluid contains high levels of glutathioneJ Appl Physiol19876311521573040659
- BrownLASPingX-DHarrisFLGauthierTWGlutathione availability modulates alveolar macrophage function in the chronic ethanol-fed ratAm J Physiol Lung Cell Mol Physiol20072924L824L83217122355
- StockerRKeaneyJFJrRole of oxidative modifications in atherosclerosisPhysiol Rev20048441381147815383655
- HanselTTBarnesPJRecent Advances in the Pathophysiology of COPDBasel Boston Birkhäuser Verlag2004
- RahmanIMacNeeWOxidative stress and regulation of glutathione in lung inflammationEur Respir J200016353455411028671
- BarnesPJOxidative stress in COPDHanselTTBarnesPJRecent Advances in the Pathophysiology of COPDBasel; BostonBirkhäuser Verlag2004
- LoebGASkeltonDCCoatesTDFormanHJRole of selenium-dependent glutathione peroxidase in antioxidant defenses in rat alveolar macrophagesExp Lung Res198814 Suppl9219363208729
- PietarinenPRaivioKDevlinRBCrapoJDChangLYKinnulaVLCatalase and glutathione reductase protection of human alveolar macrophages during oxidant exposure in vitroAm J Respir Cell Mol Biol19951344344417546773
- ZhangWSalomonRGOxidized phospholipids, isolevuglandins, and atherosclerosisMol Nutr Food Res200549111050106216270278
- WintergerstESJelkJRahnerCAsmisRApoptosis induced by oxidized low density lipoprotein in human monocyte-derived macrophages involves CD36 and activation of caspase-3Eur J Biochem2000267196050605910998066
- WangYQiaoMMieyalJJAsmisLMAsmisRMolecular mechanism of glutathione-mediated protection from oxidized low-density lipoprotein-induced cell injury in human macrophages: role of glutathione reductase and glutaredoxinFree Radic Biol Med200641577578516895798
- PedersenNCHoEBrownMLYamamotoJKIsolation of a T-lymphotropic virus from domestic cats with an immunodeficiency-like syndromeScience19872357907933643650
- VenketaramanVRodgersTLinnaresRTuberculosis immunity in healthy and HIV-infected subjectsAIDS Res Ther200631516504020
- BuhlRJaffeHAHolroydKJSystemic glutathione deficiency in symptom-free HIV-seropositive individualsLancet19892129412982574255
- de QuayBMalinverniRLauterburgBHGlutathione depletion in HIV-infected patients: role of cysteine deficiency and effect of oral N-acetylcysteineAIDS199268158191418777
- EckHPGmunderHHartmannMPetzoldtDDanielVDrogeWLow concentrations of acid-soluble thiol (cysteine) in the blood plasma of HIV-1-infected patientsBiol Chem Hoppe Seyler19893701011082784973
- HelblingBvon OverbeckJLauterburgBHDecreased release of glutathione into the systemic circulation of patients with HIV infectionEur J Clin Invest19962638448682153
- PantaleoGGraziosiCFauciASNew concepts in the immunopathogenesis of human immuno deficiency infectionN Engl J Med19933283273358093551
- LevyJAHIV pathogenesis and long term survivalAIDS19937140114108280406
- DrogeWHolmERole of cysteine and glutathione in HIV infection and other diseases associated with muscle wasting and immunological dysfunctionFASEB J199711107710899367343
- HerzenbergLAde RosaSCDubsJGGlutathione deficiency is associated with impaired survival in HIV diseaseProc Natl Acad Sci U S A199794196719729050888
- StaalFJGlutathione and HIV infection: reduced reduced or increased oxidized?Eur J Clin Invest199821941969568464
- TsaoTCHongJLiLFHsiehMJLiaoSKChangKSImbalances between tumor necrosis factor-alpha and its soluble receptor forms, and interleukin-1beta and interleukin-1 receptor antagonist in BAL fluid of cavitary pulmonary tuberculosisChest200011710310910631206
- HagAMKristoffersenUSPedersenSFRegional gene expression of LOX-1, VCAM-1, and ICAM-1 in aorta of HIV-1 transgenic ratsPloS One20094e817019997643
- DayaramYKTalaueMTConnellNDVenketaramanVCharacterization of a glutathione metabolic mutant of Mycobacterium tuberculosis and its resistance to glutathione and nitrosoglutathioneJ Bacteriol200618841364137216452418
- BrillKJLiQLarkinRHuman natural killer cells mediate killing of intracellular Mycobacterium tuberculosis H37Rv via granule-independent mechanismsInfect Immun20016931755176511179353
- ChanEDChanJSchlugerNWWhat is the role of nitric oxide in murine and human host defense against tuberculosis? Current knowledgeAm J Respir Cell Mol Biol200125560661211713103
- FlynnJLChanJTrieboldKJDaltonDKStewartTABloomBRAn essential role for interferon-α in resistance to Mycobacterium tuberculosis infectionJ Exp Med1993178224922527504064
- VenketaramanVDayaramYKAminAGRole of glutathione in macrophage control of mycobacteriaInfect Immun20037141864187112654802
- VenketaramanVDayaramYKTalaueMTConnellNDGlutathione and nitrosoglutathione in macrophage defense against M. tuberculosisInfect Immun20057331886189415731094
- MillmanACSalmanMDayaramYKConnellNDVenketaramanVNatural killer cells, glutathione, cytokines and innate immunity against Mycobacterium tuberculosisJ Interferon Cytokine Res20082811318370866
- VenketaramanVMillmanACSalmanMGlutathione levels and immune responses in tuberculosis patientsMicrob Pathog20084425526117959342
- GutteridgeJMLipid peroxidation and antioxidants as biomarkers of tissue damageClin Chem19954112 Pt 2181918287497639
- Schulze-OsthoffKLosMBaeuerlePARedox signalling by transcription factors NF-kappa B and AP-1 in lymphocytesBiochem Pharmacol1995507357417575632
- DubeySSSinhaKKGuptaJPVitamin C status, glutathione and histamine in gastric carcinoma, tuberculous enteritis and non-specific ulcerative colitisIndian J Physiol Pharmacol19852921111144093192
- KwiatkowskaSPiaseckaGZiebaMPiotrowskiWNowakDIncreased serum concentrations of conjugated diens and malondialdehyde in patients with pulmonary tuberculosisRespir Med199993427227610464892
- JackCIJacksonMJHindCRCirculating markers of free radical activity in patients with pulmonary tuberculosisTuber Lung Dis19947521321378032046
- TsaoTCHongJLiLFHsiehMJLiaoSKChangKSImbalances between tumor necrosis factor-alpha and its soluble receptor forms, and interleukin-1beta and interleukin-1 receptor antagonist in BAL fluid of cavitary pulmonary tuberculosisChest2000117110310910631206
- MakonkawkeyoonSLimson PobreRNMoreiraALSchaufVKaplanGThalidomide inhibits the replication of human immunodeficiency virus type 1Proc Natl Acad Sci U S A199390597459788327469
- WallisRSNsubugaPOkweraAPentoxifylline in human immunodeficiency virus-seropositive tuberculosis: a randomized, controlled trialJ Infect Dis199617447277338843209
- Mayanja-KizzaHJones-LopezEOkweraAUganda-Case Western Research Collaboration. Immunoadjuvant prednisolone therapy for HIV-associated tuberculosis: a phase 2 clinical trial in UgandaJ Infect Dis2005191685686515717259
- WallisRSKalayjianRJacobsonJMA study of the immunology, virology, and safety of prednisone in HIV-1-infected subjects with CD4 cell counts of 200 to 700 mm(−3)J Acquir Immune Defic Syndr200332328128612626887
- van CrevelROttenhoffTHvan der MeerJWInnate immunity to Mycobacterium tuberculosisClin Microbiol Rev200215229430911932234
- ChristmanMA2ndGoetzDJDickersonEWnt5a is expressed in murine and human atherosclerotic lesionsJ Physiol Heart Circ Physiol20082946H2864H2870