
Abstract
The brain areas that endow insects with the ability to see consist of remarkably complex neural circuits. Reiterated arrays of many diverse neuron subtypes are assembled into modular yet coherent functional retinotopic maps. Tremendous progress in developing genetic tools and cellular markers over the past years advanced our understanding of the mechanisms that control the stepwise production and differentiation of neurons in the visual system of Drosophila melanogaster. The postembryonic optic lobe utilizes at least two modes of neurogenesis that are distinct from other parts of the fly central nervous system. In the first optic ganglion, the lamina, neuroepithelial cells give rise to precursor cells, whose proliferation and differentiation depend on anterograde signals from photoreceptor axons. In the second optic ganglion, the medulla, the coordinated activity of four signaling pathways orchestrates the gradual conversion of neuroepithelial cells into neuroblasts, while a specific cascade of temporal identity transcription factors controls subtype diversification of their progeny.
INTRODUCTION
Insects including flies have beautifully structured small brains. Although they are sometimes described as simple models for studies of brain function and development based on cell numbers, they are exquisitely complex in their own right. One brain region, for which this may apply more than any other, is the insect visual system, consisting of the compound eye and the underlying optic lobe. Ramón y Cajal and Domingo Sanchez y Sanchez began to document the intricacies of visual connectivity in the dipteran optic lobe with drawings of Golgi impregnated neurons almost a hundred years ago (CitationRamon y Cajal & Sanchez y Sanchez, 1915). However, already Henri Viallanes’ early lithographic illustrations of fly compound eyes and optic lobes during metamorphosis (CitationViallanes, 1883) highlighted a closely related enduring question: How can such a complex part of the brain form during development? Clearly, before any wiring of neuronal connections can be initiated, appropriate numbers of diverse neuron subtypes have to be produced. In this review, we focus on this first essential step in visual circuit assembly and outline our current understanding of the fundamental strategies, by which neurons are generated and specified in the visual system of the genetically tractable fruit fly Drosophila melanogaster.
Numbers and Cellular Diversity in the Drosophila Visual System
Whereas the adult central brain of Drosophila is estimated to contain 11,000 neurons in each hemisphere (CitationIto et al., 2013; CitationYu et al., 2013), their optic lobes each consist of ∼60,000 neurons (CitationHofbauer & Campos-Ortega, 1990; CitationRaghu et al., 2011). The eye constitutes a highly regular, almost crystalline array of ∼780 facets or ommatidia in female and ∼730 in male flies (CitationWolff & Ready, 1993), each containing the cell bodies of 8 photoreceptor subtypes (R cells, R1–R8). Their axons extend into the optic lobe, which is subdivided into four ganglia, the lamina, medulla, lobula plate, and lobula. All neuropils share a retinotopic organization into reiterated columnar microcircuits. The medulla, lobula plate, and lobula additionally are structured into at least ten, four, and six perpendicular interconnected synaptic layers, respectively (). While columns are equivalent to an array of channels that receive sensory input from discrete locations across the visual field, layers are devoted to extract information about specific visual features, such as color, motion, or contrast from each of these points in space (CitationClandinin & Feldheim, 2009).
Figure 1. Architecture of the adult Drosophila visual system. (A) The optic lobe consists of the lamina, medulla, lobula plate, and lobula. The illustration shows a subset of neuron subtypes and their retinotopic projections. R1–R6 cells extend axons from the retina into the lamina, where they connect with lamina neurons L1–L3 in cartridges. Axons of R8 and R7 cells and lamina neurons L1–L5 terminate in one or more of ten medulla neuropil layers (M1–M10). Transmedullary neurons (Tm and TmY) project from the medulla to subsets of six lobula (Lo) and four lobula plate (Lop) neuropil layers. Distal medulla (Dm) neurons innervate several columns in upper medulla layers. Medulla intrinsic (Mi) neurons connect distal and proximal layers. C neurons extend branches into the medulla and lamina. T neurons connect the lobula or lobula plate and medulla, or lobula and lobula plate. (B) R1–R6 axons innervate lamina cartridges following the neural superposition principle.
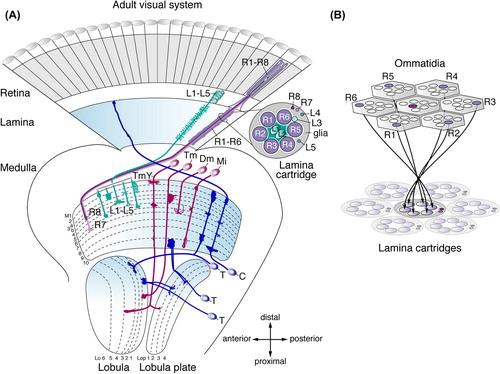
The adult lamina consists of ∼4000 neurons. R1–R6 axons innervate the lamina following the neural superposition principle (CitationBraitenberg, 1967; CitationKirschfeld, 1967; CitationMeinertzhagen & Hanson, 1993; CitationClandinin & Zipursky, 2000; CitationSanes & Zipursky, 2010; CitationHadjieconomou et al., 2011). Axons of six different R1–R6 photoreceptors in six neighboring ommatidia that receive input from the same point of space in the visual field converge into the same columnar modules, called lamina cartridges, where they form synaptic connections with three of five lamina neuron subtypes (L1–L5) (). This ensures the coherent representation of the visual field, while increasing the sensitivity to light stimuli. The topographic organization is subsequently maintained in corresponding underlying columnar modules in the medulla, lobula plate, and lobula.
R8 and R7 axons, as well as the axons of lamina neurons L1–L5 extend into the neuropil of the largest ganglion, the medulla, which consists of ∼40,000 neurons (CitationHofbauer & Campos-Ortega, 1990). R8 axons terminate in the proximal half of the medulla neuropil layer M3 (M3b; CitationTing et al., 2014), whereas R7 axons target to layer M6. In each column, the axon terminals of L1–L5 lamina neurons arborize in the distal layers M1–M5. The first comprehensive analysis in the Drosophila visual system using Golgi impregnations (CitationFischbach & Dittrich, 1989) reported ∼70 medulla neuron subtypes that have cell bodies in the surrounding cortex and form morphologically distinct arborizations within specific sets of layers in one or several columns in this neuropil. Transmedullary (Tm) and transmedullary Y (TmY) neurons are projection neurons that relay information from the medulla to the lobula or to the lobula and lobula plate, respectively. Interneurons include medulla intrinsic (Mi) neurons that connect distal and proximal medulla neuropil layers, and amacrine cells that innervate several columns in distal or proximal layers (Dm and Pm neurons). Moreover, ascending neuron subtypes connect the medulla and lamina (CitationTuthill et al., 2013). Finally, synaptic columns are innervated by numerous tangential medulla neuron subtypes with wide-field projection patterns in defined layers (CitationFischbach & Dittrich, 1989). The total number of medulla neuron subtypes is set to increase as systematic genetic single cell labeling and electron microscopy approaches continue to report new subtypes (CitationMorante & Desplan, 2008; CitationHasegawa et al., 2011; CitationRaghu & Borst, 2011; CitationRaghu et al., 2011; CitationRaghu et al., 2013; CitationTakemura et al., 2013). Recent dense serial electron microscopy reconstructions identified ∼40 distinct medulla neuron subtypes with columnar branching patterns (CitationTakemura et al., 2013). These include synperiodic neurons present once in every column (1 neuron/1 column) or ultraperiodic neurons found in every column, but with more than one representative (> 1 neuron/1 column). Additionally, there are infraperiodic subtypes, which are not present in each column (1 neuron/> 1 column) (CitationTakemura et al., 2013).
The lobula plate and lobula consist of ∼15,000 neurons (CitationHofbauer & Campos-Ortega, 1990). So far, ∼28 neuron subtypes with cell bodies adjacent to the lobula plate neuropil have been mapped (CitationFischbach & Dittrich, 1989). These include neurons with columnar arborizations interconnecting distinct medulla, lobula, and lobula plate layers, as well as ascending neuron subtypes that ramify in the medulla and lamina. The perhaps best-studied neuron subtypes in the lobula plate include the motion-sensitive giant lobula plate tangential cells (LPTC), whose dendritic branches receive visual input from hundreds of columns (CitationJoesch et al., 2008). Finally, at least 24 different subtypes of visual projection neurons have been associated with the lobula: these either connect the lobula or the lobula, lobula plate and medulla with the optic glomeruli or other higher visual processing centers within the central brain (CitationOtsuna & Ito, 2006; CitationStrausfeld et al., 2007).
The initial step toward connecting R-cell axons and higher order neurons within a coherent functional retinotopic map, thus, begins with a formidable challenge of producing sufficient numbers and cellular diversity during neurogenesis. First, significantly more neurons are generated in the optic lobe compared to the central brain. Second, these neurons differentiate into many specific subtypes at a scale of diversity that is unexpectedly comparable to that found in the vertebrate retina (CitationSanes & Zipursky, 2010). Third, because there is not always a one to one ratio of neurons per column, precise numbers of each subtype need to be produced for each column or groups of columns in each ganglion, and ratios of numbers must be coordinated to match between the eye and across ganglia. Consistently, over the recent years it has emerged that neurogenesis in the optic lobe follows developmental rules, which are to some extent distinct from those in other parts of the central nervous system (CNS).
For Comparison: Neurogenesis in the Drosophila Central Brain and Ventral Nerve Cord
Neuroblasts (NBs), the neural stem cells in the fly central brain and ventral nerve cord (VNC), are generated during embryonic development. Embryonic NBs arise from the ventrolateral neuroectoderm (reviewed in: CitationHomem & Knoblich, 2012). They are singled out by Notch-dependent lateral inhibition and activation of proneural gene expression, and subsequently delaminate (CitationCampos-Ortega, 1993). Most commonly, NBs follow a type I mode of neurogenesis: they undergo repeated asymmetric divisions to self-renew and to generate a ganglion mother cell (GMC). This frequently smaller daughter cell then divides once symmetrically to produce two offspring, which can be either neurons or neurons and glia. These form the embryonic and larval functional CNS and about 10% of the adult CNS (CitationTruman et al., 2004). During late embryogenesis, a significant proportion of NBs in abdominal VNC segments undergoes apoptosis (CitationWhite et al., 1994), while four NBs in the mushroom bodies and one NB lateral to the antennal lobe continue to proliferate (CitationIto & Hotta, 1992). Most NBs in the central brain and VNC, however, transiently enter a quiescent state characterized by the arrest in the G1 phase of the cell cycle (reviewed in: CitationBrand & Livesey, 2011). From the second half of the first instar larval stage onward these NBs resume proliferation generating 90% of neurons in the adult CNS in a second phase of neurogenesis (CitationTruman et al., 2004). Finally, cell divisions cease during early pupal development.
The majority of ∼114 lineages identified in the adult central brain are generated by type I NBs (CitationIto et al., 2013; CitationYu et al., 2013). However, in medial regions of the central brain, eight postembryonic NBs follow a different division pattern: these type II NBs give rise to transit-amplifying intermediate neural precursors, which in self-renewing asymmetric divisions produce GMCs and ultimately neurons and glia (CitationBello et al., 2008; CitationBoone & Doe, 2008; CitationBowman et al., 2008; CitationBayraktar & Doe, 2013). In the central brain and the VNC, neurons are thus generated during two phases of neurogenesis by NBs that originated from the embryonic neuroectoderm. By contrast in the optic lobe, neurons are derived from two active neuroepithelia during postembryonic development (see below).
Early Optic Lobe Development: The Beginning
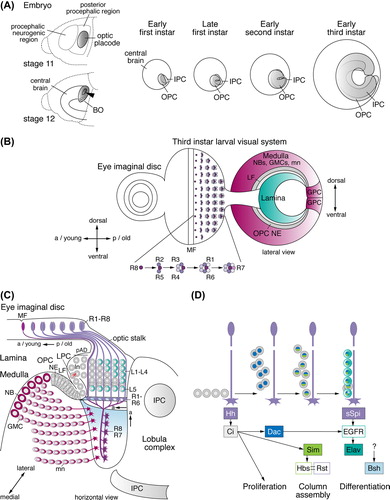
The development of the optic lobe is initiated during embryogenesis. From stages 8 to late 11, ∼100 NBs delaminate from the procephalic neurogenic region in the head ectoderm to generate the central brain (CitationUrbach & Technau, 2003, Citation2004). At stage 11, an adjacent coherent group of cells, which has undergone four rounds of cell divisions in the posterior procephalic region, differentiates into the optic placode (CitationGreen et al., 1993). These columnar shaped cells invaginate during stages 12 and 13. They initially form a pouch and then a flat vesicle of approximately 35 to 80 cells, which attaches to the lateral surface of the brain (CitationWhite & Kankel, 1978; CitationHofbauer & Campos-Ortega, 1990; CitationGreen et al., 1993). Invagination requires the activity of the SIX homeobox transcription factor Sine oculis (CitationCheyette et al., 1994). Optic placode cells remain quiescent during subsequent stages of embryonic development, but begin to proliferate after larval hatching. They become subdivided into two primordia, called the outer and inner proliferation centers (OPC and IPC) (CitationWhite & Kankel, 1978) or alternatively outer and inner optic anlagen (CitationHofbauer & Campos-Ortega, 1990). The superficially located OPC is dedicated to generating neurons in the lamina and medulla, while the centrally located IPC gives rise to lobula plate and lobula neurons (CitationMeinertzhagen & Hanson, 1993).
As both primordia grow, the superficially located OPC and the more centrally located IPC become horse-shoe shaped by the end of the first instar larval stage and the beginning of the second instar larval stage, respectively (CitationNassif et al., 2003) (). By the end of the second instar larval stage, the OPC expands to approximately 700 and the IPC to 400 cells, and the initially adjacent primordia become separated by newly produced postmitotic neurons (CitationHofbauer & Campos-Ortega, 1990; CitationNassif et al., 2003). The arrival of new molecular and genetic markers in the recent years made it possible to extend these early insightful studies and unambiguously identify neuroepithelial (NE) cells, NBs and other neural progenitor types. This revealed that the initially formed cells in the OPC and IPC are of NE character and divide symmetrically to expand the pool of cells that subsequently give rise to different types of neural progenitors (CitationEgger et al., 2007). In the embryonic neuroectoderm, individual cells develop into NBs, while the remaining cells are specified as epidermal cells (CitationCampos-Ortega, 1993). By contrast, the OPC and IPC constitute neuroepithelia, in which all cells eventually convert into neural progenitors (CitationEgger et al., 2007). Likely as a consequence of a duplication event in ancestral arthropods (CitationStrausfeld, 2005), the OPC neuroepithelium is subdivided: the lateral domain produces lamina neurons, while the medial domain facing the central brain generates medulla neurons (CitationEgger et al., 2007; CitationYasugi et al., 2008). These domains produce their offspring by two distinct modes of neurogenesis.
Figure 2. Early optic lobe development and lamina neurogenesis. (A) Embryonic stages 11 and 12: the optic placode forms in the posterior procephalic region of the head ectoderm, invaginates (arrowhead), and attaches to the central brain. Ventral cells develop into Bolwig's organ (BO), containing the larval photoreceptor cells. At the early first instar larval stage, the outer and inner proliferation centers (OPC and IPC) are visible. They expand by symmetric divisions and become crescent-shaped during the late first and early second instar larval stages, respectively. The OPC and IPC separate, as they begin to produce offspring from the early third instar larval stage onward. (B) In the third instar larval eye imaginal disc, rows of R-cell clusters progressively arise posteriorly (p) of the morphogenetic furrow (MF). R8 cells sequentially recruit R2/R5, R3/R4, R1/R6, and R7 cells into each cluster. A lateral view of the optic lobe shows the crescents of the lamina, the OPC neuroepithelium (NE) and medulla NBs, GMCs, and medulla neurons (mn). Lamina neurogenesis occurs in the wake of the lamina furrow (LF). Glial precursor cell (GPC) areas are located at the dorsal and ventral tips of the OPC crescent. (C) A horizontal view of the third instar larval eye imaginal disc and optic lobe shows the stepwise formation of lamina neurons (ln) and medulla neurons. Medially, the OPC NE gives rise to medulla NBs, GMCs and medulla neurons. Laterally, the OPC NE generates lamina precursor cells (LPCs). The position of the NE to LPC conversion at the bottom of the lamina furrow is approximate in this schematic presentation. LPCs give rise to undifferentiated lamina neurons in the pre-assembly domain (pAD). Lamina neurons assemble into columns in close association with R-cell bundles and differentiate into neuronal subtypes L1–L5. The IPC produces neurons of the lobula complex. (D) R-cell axons provide the anterograde signals Hedgehog (Hh) and Spitz (sSpi) to control LPC proliferation, lamina column assembly and lamina neuron differentiation. a, anterior; Bsh, Brain-specific homeobox; Ci, Cubitus interruptus; Dac, Dachshund; EGFR, epidermal growth factor receptor; Elav, Embryonic lethal abnormal vision; Hbs, Hibris; Sim, Single-minded; Rst, Roughest.
Generating Lamina Neurons
Eye Development in Brief
As early as 1943, Maxwell E. Power had uncovered that eye and optic lobe development are intricately connected, because mutants with smaller or no eyes exhibited optic lobes with a smaller or no lamina (CitationPower, 1943). Eye development is initiated during embryogenesis with a cluster of 6–23 cells in the embryonic ectoderm that invaginates and develops into the eye-antennal imaginal disc (CitationWolff & Ready, 1993). During early larval development these epithelial founder cells proliferate extensively, while specific interactions of signaling pathways subdivide the disc into an anterior part that gives rise to the antenna and a posterior part dedicated to generating the eye and head capsule. A transcription factor cascade including members of the retinal determination gene network (RDGN) directs the specification of the eye field (for details, see: CitationKumar, 2010). From the mid third instar larval stage to 24 hours after puparium formation (APF), a differentiation wave progresses from the most posterior part of the eye disc in anterior direction across the entire eye territory. The wave front is marked by an indentation in the eye disc epithelium, called the morphogenetic furrow. In its wake, R cells sequentially differentiate cell by cell, cluster by cluster and row by row in a 2 hour rhythm (CitationTreisman, 2013). Photoreceptors are not clonally related but recruited by precisely orchestrated cell–cell interactions. In each assembling ommatidial cluster, R8 is specified first, followed by R2 and R5, R3 and R4, R1 and R6, and finally R7 (CitationTreisman, 2013) (). R cells from each ommatidial cluster are thought to extend axons through the optic stalk into the optic lobe in a single bundle, as soon as they have been specified and in the same temporal order. R1–R6 growth cones terminate in the lamina plexus, while R8 and R7 axons extend through the lamina into the medulla (). Consistent with their birth order and relative positions in the eye field, R-cell fibers project to precise locations within the optic lobe to establish a retinotopic map along the anterior-posterior and dorso-ventral axes (CitationClandinin & Feldheim, 2009).
R-cell Dependent Neurogenesis in the Lamina
With the arrival of R-cell axons in the optic lobe during the mid third instar larval stage at around 72 hours after larval hatching, the OPC neuroepithelium undergoes a significant morphological change with the appearance of a deep groove—called the lamina furrow at its lateral edge (CitationHofbauer & Campos-Ortega, 1990; CitationSelleck & Steller, 1991; CitationMeinertzhagen & Hanson, 1993). Lateral OPC NE cells gradually give rise to lamina precursor cells (LPCs) located posteriorly of the lamina furrow. These represent a distinct progenitor cell type, which does not express NB markers (CitationEgger et al., 2007). Each LPC undergoes a terminal symmetric division to generate undifferentiated lamina neurons (CitationHuang & Kunes, 1996). New R-cell axon bundles successively extend posteriorly of these immature lamina neurons located in a region defined as pre-assembly domain (CitationUmetsu et al., 2006). Seven neurons are incorporated into lamina columns in close association with each older R-cell axon fascicle. Five cells differentiate into lamina neuron subtypes L1–L5 (CitationHuang & Kunes, 1996) ().
During the past years a series of studies considerably advanced our understanding of the molecular mechanisms that control lamina neurogenesis. The youngest R-cell axon bundles extend into the immediate vicinity of the lower posterior shank of the lamina furrow, where LPCs in the G1 phase of the cell cycle are located (CitationSelleck et al., 1992). R-cell axons release the anterograde signal Hedgehog (Hh) leading to Cubitus interruptus activation in LPCs (). This triggers their progression from G1 to S phase, enabling them to undergo a final division and to generate lamina neurons (CitationSelleck & Steller, 1991; CitationSelleck et al., 1992; CitationHuang & Kunes, 1996; CitationChu et al., 2006). In parallel, Hh signaling induces expression of the basic helix-loop-helix Per-Arnt-Sim (bHLH-Pas) protein Single-minded (CitationUmetsu et al., 2006). This promotes the expression of the immunoglobulin superfamily protein Hibris in immature lamina neurons. Interactions with Irregular Chiasm C/Roughest (IrreC/Rst), a second Irre cell recognition module (IRM) protein (CitationFischbach et al., 2009) expressed by R-cell axons, enable the recruitment of lamina neuron sets into columns (CitationSugie et al., 2010). Finally, Hh promotes the expression of the RDGN transcription factor Dachshund (Dac), which is essential for epidermal growth factor receptor (EGFR) upregulation in lamina neurons (CitationHuang & Kunes, 1998; CitationHuang et al., 1998; CitationChotard et al., 2005). This endows neurons with the competence to respond to a second anterograde R-cell derived signal, the EGF-like ligand Spitz (Spi), which induces their maturation into neuron subtypes L1–L5 expressing the pan-neuronal differentiation marker Embryonic lethal abnormal vision (Elav) (CitationRobinow & White, 1991; CitationHuang et al., 1998; CitationYogev et al., 2010). Furthermore, they begin to express subtype-specific determinants such as the Brain-specific homeobox (Bsh) transcriptional regulator, which is required for the specification of lamina neuron subtypes L4 and L5 (CitationHuang et al., 1998; CitationHasegawa et al., 2013). Thus, along the anterior-posterior axis, the temporal differentiation pattern of R cells in the eye is transformed into a temporal pattern of innervation and, by the use of two anterograde signals, into a wave of proliferation, column assembly, and differentiation of lamina neurons. This ensures that the numbers of ommatidia and underlying lamina cartridges match (CitationHuang & Kunes, 1996).
Generating Medulla Neurons
Sequential Neuroepithelial to Neuroblast Conversion in the Medulla
The “Proneural Wave”
From the early third instar larval stage onward, cells at the medial edge of the OPC NE crescent begin to lose their columnar shape and adherens junctions and convert into NBs. Whereas NE cells increase in number by symmetric divisions, medulla NBs follow the proliferation pattern of type I NBs. They divide asymmetrically to self-renew, while also budding off GMCs. These in turn give rise to tightly packed pearl strings of postmitotic neurons and also glia in one symmetric division (CitationEgger et al., 2007; CitationLi et al., 2013b; see also below).
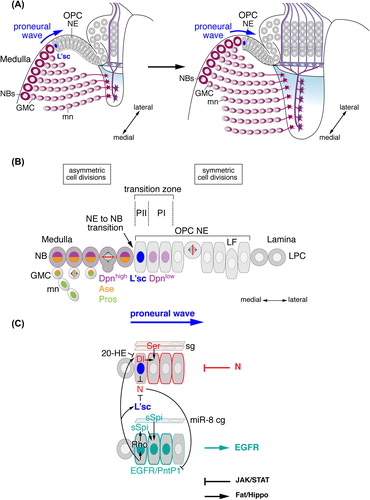
NE cells are sequentially converted into medulla NBs in a wave of neurogenesis that progresses from the medial to the lateral OPC (). Proneural genes of the Achaete-Scute complex encode four basic helix-loop-helix (bHLH) transcription factors: Achaete (Ac), Scute (Sc), Lethal of scute (L'sc), and Asense (Ase) (CitationCabrera et al., 1987; CitationBrand et al., 1993; CitationJarman et al., 1993; reviewed in: CitationGarcia-Bellido & de Celis, 2009). In the OPC, Ac is not expressed, while sc mRNA is present in all NE cells and NBs, and Ase protein in NBs and GMCs (CitationEgger et al., 2007; CitationYasugi et al., 2008). The wide expression of scute supports the notion that the OPC constitutes a neuroepithelium (CitationEgger et al., 2007). By contrast, along the entire OPC crescent, L'sc is transiently expressed in a narrow band of one to two NE cells that will transform into medulla NBs. Because L'sc expression is displaced laterally in a highly synchronized manner, as NBs are generated, this progressive differentiation pattern in the OPC has been named “proneural wave” (CitationYasugi et al., 2008). l'sc mediates the timely onset of NB formation because its loss causes a delay in NB formation (CitationYasugi et al., 2008). Moreover, gain-of-function approaches indicate that this proneural factor is sufficient to induce the NE to NB transition (CitationYasugi et al., 2008; CitationEgger et al., 2010).
Figure 3. Medulla neurogenesis depends on the sequential conversion of neuroepithelial (NE) cells to NBs. (A) Outer proliferation center (OPC) NE cells gradually convert into medulla NBs in a medial to lateral orientation. The advancement of the proneural wave is defined by Lethal of scute (L'sc) expression. (B) The gradual conversion of NE cells to NBs involves a switch from symmetric to asymmetric cell divisions. The transition zone consists of progenitors PII expressing L'sc and PI expressing low levels of Deadpan (Dpn). NBs express Dpn and Asense (Ase). GMCs express Ase and Prospero (Pros). Newly formed medulla neurons (mn) maintain Pros. (C) The progression of the proneural wave from medial to lateral is negatively regulated by the Notch (N) pathway and positively regulated by the epidermal growth factor receptor (EGFR) pathway. N signaling is activated in PI NE cells by Delta (Dl) in PII and by Serrate (Ser) in surface glia (sg). Dl expression is repressed by Ecdysone (20-HE, 20-Hydroxyecdysone) signaling. EGFR signaling activates Pointed P1 (PntP1) and Rhomboid (Rho), which promote the secretion of Spitz (sSpi) from PII. sSpi is also provided by a cortex glia subtype (cg) expressing the microRNA miR-8. N and EGFR signaling regulate each other. The JAK/STAT and Fat/Hippo signaling pathways provide additional negative and positive inputs, respectively. LF, lamina furrow; LPC, lamina precursor cells.
Coordinated Actions of Notch, EGFR, JAK/STAT, and Hippo/Fat Signaling Pathways
A series of recent studies provided evidence that the gradual conversion of NE cells into NBs in the medial OPC is tightly regulated by the coordinated activity of four signaling pathways (). Dynamic regulation of Notch signaling lies at the heart of the molecular machinery (CitationEgger et al., 2010; CitationNgo et al., 2010; CitationReddy et al., 2010; CitationYasugi et al., 2010; CitationOrihara-Ono et al., 2011; CitationWang et al., 2011b; CitationWeng et al., 2012). The Notch receptor is strongly expressed in NE cells, with the exception of L'sc-positive NE cells adjacent to medulla NBs (also defined as progenitors PII), in which levels are reduced. By contrast, Delta, one of the key activating ligands of Notch signaling in the OPC, shows elevated expression in these NE cells. Strong expression of Notch target genes, such as the Enhancer of Split (E(spl)) family member mγ, indicates that Delta activates Notch signaling in immediately adjacent lateral NE cells (also defined as progenitors PI) (CitationYasugi et al., 2010; CitationEgger et al., 2011). In L'sc expressing NE cells, high levels of Delta have been proposed to attenuate Notch signaling by cell-autonomous cis-inhibition (CitationEgger et al., 2010; CitationReddy et al., 2010; CitationWeng et al., 2012). Additionally, activity of Numb, an endocytic adapter protein, contributes to the negative regulation of Notch signaling in forming NBs (CitationWang et al., 2011b). Knockdown or loss of Notch signaling during early larval development leads to the formation of a small medulla cortex with few medulla NBs. In optic lobes, in which Notch, Delta or Notch pathway components have been genetically manipulated, mutant NE cells transform precociously into more lateral NBs, which can be identified by expression of the NB-specific bHLH transcription factor Deadpan (Dpn) (CitationBier et al., 1992). Moreover, clones lacking Notch function are frequently extruded from the OPC neuroepithelium, and arising NBs are located in ectopic positions in the underlying medulla cortex (CitationEgger et al., 2010). These findings are consistent with the notion that Notch signaling is required for maintaining the OPC NE state, especially in lateral cells immediately adjacent to the transition zone, and for preventing the premature conversion into NBs. Hence, the specific function of Notch signaling in NE maintenance initially contributes to the expansion of the neuroepithelium during early larval stages, while timely attenuation of pathway activity is essential for the transformation of NE cells into NBs (CitationEgger et al., 2010; CitationNgo et al., 2010; CitationReddy et al., 2010; CitationYasugi et al., 2010; CitationOrihara-Ono et al., 2011; CitationWang et al., 2011b; CitationWeng et al., 2012).
The EGFR pathway provides crucial input to drive the sequential progression of the proneural wave across the OPC (CitationYasugi et al., 2010). EGFR signaling activates the Ras pathway to phosphorylate the ETS domain transcription factor Pointed P2 (PntP2) and to induce the expression of the Pnt isoform PntP1 (CitationO’Neill et al., 1994). pnt mutant clones show reduced growth, suggesting a role of EGFR signaling in initial NE cell amplification (CitationYasugi et al., 2010). Expression analysis of PntP1 indicates that this pathway is active in three to four NE cells at the medial edge of the OPC crescent. This transition zone includes the L'sc expressing progenitors PII and the two immediately adjacent lateral NE cells, the progenitors PI (CitationYasugi et al., 2010). The latter express low levels of Dpn and likely correspond to mγ-positive NE cells characterized by high N signaling (CitationEgger et al., 2011). EGFR pathway activation is necessary and sufficient for proneural wave progression, because: (1) in Egfr or pnt deficient clones, L'sc expression is absent, NBs fail to form and the neuroepithelium expands medially; and (2) over-expression of a constitutively active form of the EGFR receptor causes the proneural wave front marked by L'sc to move laterally and NBs to form prematurely (CitationYasugi et al., 2010). mRNA of the EGFR ligand Spi is generally widely expressed. The seven-pass transmembrane protein Rhomboid (Rho) posttranslationally regulates the proteolytic cleavage of Spi in the Golgi apparatus to generate a soluble form of this activating ligand (sSpi) (CitationFreeman, 2008). Genetic analysis indicates that EGFR signaling induces the expression of Rho in L'sc-positive progenitors PII, which in turn secrete sSpi to activate the EGFR pathway in immediately adjacent NE cells in the transition zone. This positive feedback loop enables the sequential activation of EGFR signaling (CitationYasugi et al., 2010). Importantly, EGFR signaling induces the expression of both Delta and L'sc (CitationYasugi et al., 2010), and L'sc is able to downregulate Notch expression in NE cells (CitationEgger et al., 2010). Conversely, over-expression of activated Notch delays PntP1 expression, indicating that N and EGFR signaling are mutually dependent (CitationYasugi et al., 2010). Thus, not an epistatic hierarchy of the EGFR and Notch pathways, but the crosstalk between these two central signaling pathways orchestrates the directional progression of the proneural wave.
The Janus Kinase (JAK)/signal transducer and activator of transcription (STAT) pathway plays a dual role in promoting initial NE expansion in the OPC and inhibiting proneural wave progression (CitationYasugi et al., 2008; CitationNgo et al., 2010; CitationWang et al., 2011a; CitationWang et al., 2013). Loss of JAK/STAT signaling leads to loss of epithelial integrity, whereas over-activation of this pathway induces NE overgrowth (CitationWang et al., 2011a). NE growth is mediated in part by the direct STAT target gene Nop56, a regulator of ribosome biogenesis and cell growth (CitationWang et al., 2013). In clones that are mutant for components of the JAK/STAT pathway, NE cells prematurely convert into NBs (CitationYasugi et al., 2008; CitationNgo et al., 2010; CitationWang et al., 2011a). The JAK/STAT ligand Unpaired (Upd) is expressed in the most lateral NE cells close to the lamina furrow. Moreover, a 10xSTAT92E-GFP transgene reports graded pathway activation that decreases from the lateral to the medial OPC. These observations support the model that medial NE cells are successively released from negative JAK/STAT signaling enabling them to express L'sc and to transform into NBs (CitationYasugi et al., 2008). Although it has not been fully explored at what level pathways intersect, JAK/STAT signaling is thought to act upstream of Notch and EGFR signaling and to influence the timing of pathway activation via Delta and Rho expression (CitationYasugi et al., 2010).
The Fat/Hippo pathway mediates controlled cell proliferation in OPC NE cells in line with its well-established conserved function in regulating growth and survival (CitationReddy et al., 2010). The transmembrane molecule Fat and the cytoskeleton-binding protein Expanded activate the Hippo and Warts kinase complex. This results in the phosphorylation and inhibition of the transcriptional co-activator Yorkie (Yki) and consequently the repression of proliferative target genes, such as the microRNA (miRNA) bantam (ban) (CitationZhao et al., 2011). Loss of ban reduces OPC growth (CitationLi & Padgett, 2012), while ectopic expression of an activated form of Yki leads to the tumorous expansion of the OPC neuroepithelium without affecting NB proliferation (CitationReddy et al., 2010; CitationKawamori et al., 2011). In addition, this pathway is required for the timely conversion of NE cells into NBs, because this step is blocked or delayed by expression of activated Yki or accelerated in yki loss-of-function clones (CitationReddy et al., 2010; CitationKawamori et al., 2011).
OPC NE cells in the transition zone undergo an elongated G1 phase before transforming into NBs (CitationReddy et al., 2010; CitationOrihara-Ono et al., 2011; CitationWeng et al., 2012). This transient pause in the cell cycle is regulated by Fat/Hippo signaling in part through the cell cycle regulator E2f1, and may be essential for the accumulation of sufficient levels of Delta and in consequence attenuation of Notch signaling by cis inhibition (CitationReddy et al., 2010). Alternatively, since ectopic expression of constitutive-active RasV12 or rho partially rescues NE to NB transition defects in fat mutants, Fat/Hippo signaling may control EGF ligand secretion or EGFR signal transduction and therefore proneural wave progression (CitationKawamori et al., 2011). The crucial link between the cell cycle progression rate and NE cell to NB conversion is further supported by studies of replication protein A (RPA) and other core cell cycle regulators, because their loss results in precocious medulla NB formation due to perturbation of Notch signaling by Numb mislocalization and simultaneous EGFR pathway activation (CitationZhou & Luo, 2013).
Contribution of Systemic Signals and Local Glial Cell Niche-Derived Cues
Systemic signals ensure that stem cell proliferation in individual tissues and the general growth of a developing animal are coordinated. Consistently, postembryonic NBs have been shown to exit quiescence and begin to proliferate in response to a dietary nutrient-dependent signal released from the fat body, a tissue that plays a central role in controlling metabolism and growth in insects (CitationBritton & Edgar, 1998; reviewed in: CitationBrand & Livesey, 2011). The initial expansion and maintenance of the OPC neuroepithelium by symmetric cell divisions depend on Insulin Receptor/Target of Rapamycin (InR/TOR) signaling and are sensitive to nutrient restriction. However, OPC NBs are insensitive, and thus able to generate diverse offspring by asymmetric divisions (CitationLanet et al., 2013). The steroid hormone Ecdysone (20-HE) constitutes another major systemic signal released by the prothoracic gland that is essential for coordinating developmental transitions such as larval molts and metamorphosis (reviewed in: CitationYamanaka et al., 2013). Analysis of CNS explants and feeding experiments hinted that Ecdysone can activate proliferation of quiescent NBs (CitationDatta, 1999; CitationPark et al., 2001). Indeed recent studies uncovered that a pulse of Ecdysone during the mid third instar larval stage leads to pathway activation in OPC NE cells, which promotes NE to NB conversion by reducing Delta expression (CitationLanet et al., 2013).
The Drosophila CNS is ensheathed by the layered processes of two glial cell subtypes, the perineurial and subperineurial glia (CitationFreeman & Rowitch, 2013). The latter form pleated septate junctions and both cell types form the blood-brain barrier. In addition, processes of cortex glia wrap groups or individual neuron cell bodies (reviewed in: CitationFreeman & Rowitch, 2013). Earlier observations have linked gliogenesis and neurogenesis in the optic lobe by suggesting that glial cells secrete the glycoprotein Anachronism (Ana) to inhibit proliferation (CitationEbens et al., 1993). The release of Ana-mediated repression and activation of proliferation requires Perlecan, a heparan sulfate proteoglycan encoded by the gene terribly reduced optic lobes (trol) (CitationDatta, 1995; CitationVoigt et al., 2002). However, it is not known, which glial cell type is responsible and whether NE expansion or NB proliferation are affected. Recent studies revealed that perineurial and subperineurial surface glia express Serrate, a second activating ligand of Notch signaling. Serrate is required in glia to regulate their proliferation and to non-autonomously promote the maintenance of the OPC neuroepithelium while inhibiting premature progression of the proneural wave (CitationPerez-Gomez et al., 2013). Moreover, a distinct subpopulation of cortex glia has been identified, that specifically expresses the miRNA miR-8 and is closely associated with the OPC neuroepithelium (CitationMorante et al., 2013). miR-8 controls the sprouting of long processes by this glial subtype, and the local release of the ligand sSpi by directly binding the 3’UTR of its mRNA. Glia-derived sSpi nonautonomously activates EGFR signaling in the underlying neuroepithelium, promoting its expansion and timely NB formation. Glial cells thus provide local molecular cues in a defined specialized niche microenvironment.
The combined action of the above outlined molecular mechanisms must ultimately result in the formation of a sufficient number of NBs. These in turn have to generate appropriate numbers of neuron subtypes innervating each medulla neuropil column. The easiest assumption would be that the number of NBs equals that of medulla neuropil columns and, in consequence, that of the 730–780 ommatidial clusters and lamina cartridges. Medulla neuron development is initiated during the early third instar larval stage prior to R-cell innervation and lamina neuron differentiation (CitationHofbauer & Campos-Ortega, 1990). Moreover, the reduced size of the medulla in animals lacking R-cell input can be largely attributed to neuronal degeneration during the first half of pupal development (CitationFischbach, 1983; CitationFischbach & Technau, 1984). Medulla neurogenesis has therefore been thought to rely on regulatory mechanisms independent of afferent input from the eye or the lamina. However, in Stat mutant clones, Dac-positive LPCs fail to form in the lamina and medulla NBs expand into this area (CitationYasugi et al., 2008), suggesting that neurogenesis from the lateral and medial edges of the OPC neuroepithelium may be interdependent. The detailed mechanisms coordinating eye and lamina development with medulla neurogenesis thus deserve further exploration in the future.
Sequential Generation of Medulla Neurons by Neuroblasts
As soon as medulla NBs have formed, they begin to undergo asymmetric cell divisions (). Similar to their counterparts in the central brain and VNC, these NBs express the bHLH transcription factors Dpn and Ase, as well as Miranda (Mira) (CitationWallace et al., 2000; CitationEgger et al., 2007). Dpn is a member of the Hairy-E(spl) (HES)-family of bHLH repressors (CitationBier et al., 1992), while Ase belongs to the Achaete-Scute complex family of proneural bHLH activators (CitationGarcia-Bellido & de Celis, 2009). Mira is a coiled-coil protein that serves as an adaptor for basally localized components of the asymmetric cell division machinery (CitationIkeshima-Kataoka et al., 1997). GMCs express Ase and the homeodomain transcription factor Prospero (Pros) (CitationDoe et al., 1991). The latter is asymmetrically localized in the cytoplasm of NBs and segregates into GMCs, where it enters the nucleus (reviewed in: CitationBrand & Livesey, 2011; CitationHomem & Knoblich, 2012). After GMCs have completed one symmetric division, Pros levels remain high in newly formed medulla neurons (CitationColonques et al., 2011). dpn plays a central role in promoting NB proliferation by repressing the expression of the Cyclin dependent kinase inhibitor Dacapo (Dap), whereas ase terminates mitotic activity in NB progeny by upregulating Dap (CitationWallace et al., 2000). Moreover, transient upregulation of Pros in medulla neurons promotes cell cycle exit by inhibiting dpn and in consequence activation of dap (CitationColonques et al., 2011). In addition, two optic lobe-derived TGF-β family members, Activin-β and Dawdle, have been implicated in redundantly regulating the generation and proliferation of NBs by activating the type I receptor Baboon and its transcriptional regulator Smad2 in these progenitors (CitationZhu et al., 2008).
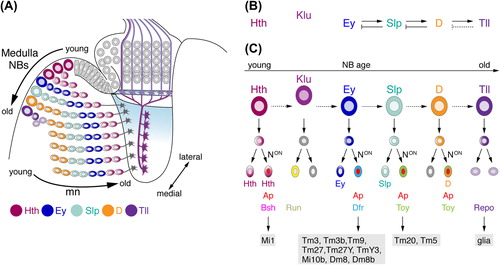
Each medulla NBs sequentially produces a set of postmitotic neurons, which initially align in columns. The relative positions of individual medulla neurons within the cortex correlate with their birth order along two temporal axes of differentiation (CitationWhite & Kankel, 1978; CitationHofbauer & Campos-Ortega, 1990; CitationHasegawa et al., 2011; CitationMorante et al., 2011) (): First, the youngest columns are situated in the vicinity of the OPC neuroepithelium, whereas older columns gradually become displaced to more medial locations close to the central brain. Second, within each column, the youngest neurons are located adjacent to their NBs beneath the cortex surface and the oldest neurons close to the emerging neuropil. During the first 24 hours of pupal development, clonally related medulla neurons in each column disperse within the cortex in radial and tangential orientation (CitationHasegawa et al., 2011; CitationMorante et al., 2011), consistent with the observed salt and pepper distribution of medulla neuron clones in the adult cortex (CitationBazigou et al., 2007). While the significance of this process is still unclear, the homeodomain transcription factor Eyeless (Ey) has been suggested to control this process because expression of a dominant-negative form of this determinant inhibits the repositioning of medulla neuron cell bodies due to increased E-cadherin expression (CitationMorante et al., 2011).
Figure 4. Temporal patterning of medulla NBs and Notch (N) signaling contribute to the generation of diverse neuron subtypes. (A) NE cells gradually convert into NBs. As medulla NBs age, they sequentially express the transcription factors Homothorax (Hth), Eyeless (Ey), Sloppy paired 1 and 2 (Slp), Dichaete (D), and Tailless (Tll). Each NB produces a column of medulla neurons. Progeny maintain the expression of the determinant present in the NB at the time of their birth. Older NBs and their offspring are located more medially. In each lineage, the oldest neurons are positioned closest to the medulla neuropil. Klumpfuss (Klu) is expressed in NBs but not in progeny and therefore has not been included in this schematic. Interim stages, during which NBs express more than one factor and neurons downregulating factors are also not shown. (B) Ey, Slp, and D are required for the transition to the next determinant. Slp, D, and Tll are necessary to repress the preceding factor in the series. Tll is sufficient but not required to repress D (dashed line). (C) N-mediated binary cell fate choices further diversify lineages. Progeny, in which N signaling is on, express Apterous (Ap). Depending on the combination of transcription factors present, the expression of subtype-identity determinants such as Brain-specific homeobox (Bsh), Runt (Run), Drifter (Dfr), or Twin of Eyeless (Toy) is induced. Lineages defined by the combinatorial expression of these determinants give rise to specific neuronal subtypes. Tll-positive NBs generate Reversed polarity (Repo)-positive glia. Dm, distal medulla neurons; Mi, medulla intrinsic neurons; Tm, transmedullary neurons; TmY, transmedullary Y neurons.
Generating Medulla Neuron Subtype Diversity
Temporal Patterning
As NBs begin to produce postmitotic neurons, distinct mechanisms must come into play that enable them to differentiate into diverse subtypes. One fundamental strategy links birth order and fate of neural progeny. Seminal studies in the embryonic CNS of Drosophila uncovered that each NB gives rise to distinct neuron subtypes in a defined time window and order and that this is controlled by temporal changes in transcription (CitationKambadur et al., 1998; CitationBrody & Odenwald, 2000; CitationIsshiki et al., 2001; CitationKanai et al., 2005; CitationMettler et al., 2006; reviewed in: CitationKohwi & Doe, 2013). NBs sequentially express the transcription factors Hunchback (Hb), Krüppel, Pou domain proteins Pdm1 and 2, Castor and Grainy head. Their expression is maintained in the GMC and its progeny, and confers distinct temporal identities to neuron subtypes born at a given time during development. Specific positive and negative cross-regulatory interactions, as well as input from additional factors provide a cell intrinsic mechanism that ensures the transition from one transcription factor to the next, while preventing the expression of the previous and the second following determinant. Finally, analysis of hb indicates that the competence of NBs to generate neuron subtypes with earlier temporal identities is suppressed in older NBs by subnuclear genome reorganization, silencing the hb locus and its targets (CitationKohwi et al., 2013).
Highlighting that temporal patterning of NBs constitutes a general principle, while the contributing factors vary, recent studies uncovered that medulla NBs rely on their own distinct cascade of temporal identity transcription factors (CitationLi et al., 2013a; CitationLi et al., 2013b; CitationSuzuki et al., 2013): they sequentially express Homothorax (Hth), Klumpfuss (Klu), Ey, Sloppy paired 1 and 2 (Slp), Dichaete (D), and Tailless (Tll) with increasing age (). Unlike in the embryo new medulla NBs are still added, while the already existing ones transition through the temporal cascade and each express one or two overlapping transcription factors depending on their age. It is therefore possible to see the range of temporal identity transcription factors across all medulla NBs at a given time point. Genetic analyses revealed specific cross-regulatory interactions between these factors (). ey, slp, and D are required for the transition to the next determinant. However, hth is not essential for expression of klu and ey, and klu not for the expression of ey. slp is essential for repression of Ey, while D inhibits Slp. Over-expression of Hth, Ey, Slp1 and 2, or D is not sufficient to induce the next temporal identity factor, and tll is sufficient but not required to repress D. Thus, additional so far unidentified determinants must act in concert with the temporal identity factors to control the timely transitions (CitationLi et al., 2013b; CitationSuzuki et al., 2013).
Binary Fate Choices by Asymmetric Notch Signaling
With the exception of Klu, which is solely expressed in NBs (CitationSuzuki et al., 2013), temporal identity factors are inherited by GMCs and by a subset of their progeny born in a specific time window. To further increase the diversity of potential subtypes, a second fundamental strategy comes into play, which relies on Notch-dependent binary fate decisions (CitationLi et al., 2013b). When GMCs give rise to two daughter cells, Notch signaling is activated solely in one sibling, inducing expression of the Lim homeodomain transcription factor Apterous (Ap) (). For offspring of Ey and Slp1/2 expressing GMCs, the sibling with active Notch signaling expresses Ap and loses the expression of the temporal identity factor, while the sibling with inactive Notch signaling is Ap-negative and maintains expression of the respective temporal identity factor. Daughter cells of Hth-positive GMCs both maintain Hth expression and the sibling with active Notch signaling upregulates Ap in addition to Hth. By contrast, offspring of D-positive GMCs with active Notch signaling co-express Ap and D, whereas siblings activate so far uncharacterized determinants.
Transcription Factors Conferring Neuronal Subtype Identity
Both strategies lead to the activation of the next set of transcription factors that can be co-expressed with temporal identity factors in conjunction with or without Ap, and are essential for neuronal subtype specification (). This includes Bsh, Runt (Run), Drifter (Dfr), and Twin-of-eyeless (Toy), as well as Lim3 and Distalless (Dll) (CitationMorante & Desplan, 2008; CitationHasegawa et al., 2011; CitationLi et al., 2013b; CitationSuzuki et al., 2013). Because of the birth-order dependent positioning, neuron subtypes that express combinations of temporal and subtype identity transcription factors are arranged in concentric rings within the medulla cell body cortex (CitationHasegawa et al., 2011). During the Hth competence window, NBs generate Bsh-positive medulla neurons. Hth expression and Notch signaling are essential for Bsh expression, and bsh is required and sufficient for specification of medulla intrinsic neurons Mi1 (CitationHasegawa et al., 2011; CitationHasegawa et al., 2013; CitationLi et al., 2013b). In the absence of bsh, Mi1 neurons remarkably adopt the branching pattern of transmedullary neurons and innervate the lobula (Tm1-like; CitationHasegawa et al., 2013). klu is required for the production of Run-positive neurons (CitationSuzuki et al., 2013). Subsequently, during the Ey competence window, NBs produce nine Dfr-positive medulla neuron subtypes, i.e., Tm3, Tm3b, Tm9, Tm27, Tm27Y, TmY3, Mi10b, Dm8, Dm8b (CitationHasegawa et al., 2011). Although Ey expression is not maintained, Dfr expression depends on ey in NBs, as well as Notch pathway activation in progeny (CitationLi et al., 2013b; CitationSuzuki et al., 2013). dfr controls axon targeting and dendritic arborizations, because mutant Tm27Y medulla neurons show altered branching patterns in specific layers of the medulla and lobula (CitationHasegawa et al., 2011). Finally, slp is essential for the generation of Toy-positive Tm20 and Tm5 neurons (CitationLi et al., 2013b).
A Potential Role for Spatial Input along the Dorso-Ventral Axis
Medulla NBs are thought to generally progress through the described series of temporal identity factors, while their progeny express similar sets of downstream transcription factors to differentiate into specific neuron subtypes. As dividing NBs mature, they can co-express two of the temporal identity factors, which may further diversify generated neuron subtypes (CitationLi et al., 2013b). However, there are also some indications for spatial variations along the dorso-ventral axis of the OPC. For instance, the homeodomain containing transcription factor Vsx1 shows spatially restricted expression in the central part of the OPC NE crescent and is required for NE cell proliferation during early larval development. Vsx1 is later expressed in transmedullary neuron subtypes, suggesting a second role in neuron subtype specification (CitationErclik et al., 2008). By contrast, the Wnt gene wingless (wg) and the TGF-β family member decapentaplegic (dpp) are expressed in adjacent domains at the dorsal and ventral tips of the OPC crescent (CitationKaphingst & Kunes, 1994). The most dorsal and ventral Wg expressing domains are commonly described as the glial precursor cell (GPC) areas, because they give rise to both glial cells migrating into the lamina, as well as medulla neurons (CitationPerez & Steller, 1996; CitationHuang & Kunes, 1998; CitationPoeck et al., 2001; CitationDearborn & Kunes, 2004; CitationChotard et al., 2005). Although the details of lineages are still unclear, they include neuron subtypes with tangential arbors extending into proximal medulla layers (CitationDearborn & Kunes, 2004). Future studies need to determine whether GPC areas could constitute distinct subdomains, dedicated to generating specific neuron subtypes, and explore the mechanisms mediating their diversity.
wg is required for inducing and maintaining dpp expression in adjacent domains. Loss of either determinant during early development decreases cell proliferation in the OPC, consistent with their general role in regulating both growth and patterning (CitationKaphingst & Kunes, 1994). NE clones mutant for adenomatous polyposis coli (APC), a tumor suppressor and negative regulator of Wg signaling, show increased proliferation in the central OPC subdomain (CitationHayden et al., 2007). The zinc finger protein Combgap (Cg) represses Wg target genes such as dpp, optomotor blind (omb), and aristaless (al) in the optic lobe (CitationSong et al., 2000). Wg pathway activation leads to downregulation of Cg, and in turn to the de-repression of Wg target genes, thus regulating the extent of their expression domains. Expression of wg can causes ectopic target gene activation in the OPC midline region. Because Cg is absent from this area, a different molecular system likely regulates Wg target gene expression outside the Cg expression domains (CitationSong et al., 2000). These observations suggest that Wg functions as a spatial cue, which patterns the OPC along the dorso-ventral axis and, thus, could provide additional input into medulla neuron subtype specification.
Together, these findings in the medulla highlight the fundamental principles, by which birth-order dependent neuron subtype specification is achieved. Yet, the picture is far from complete, and we have undoubtedly just begun the journey of identifying the transcription factors and their cross-regulatory interactions—with each other and with signaling pathways that are essential for specifying the more than 70 different neuron subtypes in the medulla.
Terminating Neurogenic Divisions
The number of neurons generated by NBs can be controlled by targeting their proliferation rate and/or timely termination of divisions. One mechanism to achieve the latter involves the switching of NBs to a final differentiative division mediated by Pros-dependent cell cycle exit (CitationChoksi et al., 2006; CitationSousa-Nunes et al., 2010). Because the oldest medulla NBs defined by Tll expression co- express nuclear Pros (CitationLi et al., 2013b) this strategy may also be used in the OPC. Moreover, final divisions involve a switch from neuro- to gliogenesis, since progeny begin to express the glial-specific marker Reversed polarity (Repo) (CitationLi et al., 2013b). In addition, a second mechanism that ends proliferation by apoptosis of NBs may apply. Around 20 hours after the onset of metamorphosis, neurogenesis in the medulla ceases. Although the identity of concerned cell types is still unclear, this coincides with the detection of markers for local cell death in proliferation zones and the cell body cortex (CitationHofbauer & Campos-Ortega, 1990; CitationTogane et al., 2012). While NBs generally stop dividing during early pupal development, a recent study using a modified lineage-labeling method unexpectedly provided evidence for continued neurogenesis in the adult medulla, albeit at low levels (CitationFernandez-Hernandez et al., 2013). Indeed, a small number of scattered cells in the medulla cortex could constitute quiescent NBs based on their expression of cytoplasmic Dpn and the ability to proliferate in response to injury.
From Neurogenesis to Connectivity
The mechanisms controlling neurogenesis in time and space shape those regulating the formation of neuronal connections within a retinotopic map. Soon after neurons are born, they are thought to extend neurites into the most anterior regions of the lamina and medulla neuropils (CitationHofbauer & Campos-Ortega, 1990; CitationMeinertzhagen & Hanson, 1993). These projections are gradually displaced posteriorly by newly arriving neurites from younger neurons. The basic coordinates of initial projections in the lamina and medulla along the anterior-posterior axis are therefore tightly linked with birth order. Because proximity of afferent and target neurons facilitates the formation of specific synaptic connections, this provides the basis for correct retinotopic map formation across ganglia at least along one axis (CitationClandinin & Feldheim, 2009).
A general challenge, which applies to all parts of the CNS, will be to understand, how the identified transcription factor combinations regulate the dynamic expression of distinct cell surface molecules or guidance cues at the single cell level. These must (1) enable individual neuron subtypes to form their characteristic dendritic and axonal arborizations, and (2) control interactions and eventually synapse formation between specific neuron subtypes within columns and layers. At present, only few links have been established between the transcriptional control of cell identity and expression of guidance molecules. For instance in the medulla, hth has been proposed to promote N-cadherin expression in Mi1 neurons and in this way to influence their branching pattern within medulla neuropil layers (CitationHasegawa et al., 2011).
Another pivotal challenge, which specifically applies to the visual system because of its distinct neurogenesis modes and the task of forming a functional topographic map, will be to determine to what extent larval columnar lineages in the cell body cortex and columnar units in the synaptic neuropil are linked. Clearly, R-cells within an ommatidium and lamina neurons in a cartridge are not lineage-related. Instead cell types are recruited into their respective units by precisely orchestrated cell–cell interactions (see section 4). Moreover, following the rules underlying the neural superposition principle, individual R1–R6 axons from a precise group of adjacent ommatidia converge into one lamina cartridge (see section 1). By contrast in the medulla, postmitotic neurons within cell body columns are derived from single NBs and thus are lineage-related (see section 5). Hence, a column at the third instar larval stage at least in part could provide the foundation for a neuropil column. Each adult synaptic column is innervated by identical sets of some medulla neuron subtypes (CitationTakemura et al., 2013). However, it is not yet known, as to whether these subtypes are lineage-related and derived from the same NB, and whether each NB generates the same set of progeny. If this is the case,—because some neuron subtypes are thought to innervate several columns and occur in lower numbers (CitationMeinertzhagen & Hanson, 1993)—supernumerary neurons may be eliminated by apoptosis. Alternatively, different NBs could produce different sets of neuron subtypes and more than one of each subtype. In this scenario, additional mechanisms need to be identified in the future, that control the coordinated distribution of neuron subtypes across synaptic columns.
CONCLUSIONS
With the arrival of new genetic tools and cellular markers our understanding of the fundamental rules governing neurogenesis in the Drosophila optic lobe made a considerable leap forward during the recent years. While the basic division pattern of NBs in the medulla is shared with the central brain and VNC, the optic lobe employs its own distinct modes of neurogenesis in the lamina and medulla. Moreover, the largely unexplored development of the IPC, which is responsible for generating neurons in the lobula plate and lobula, has yet another mode of neurogenesis for us in store (CitationHofbauer & Campos-Ortega, 1990; H.A., I.S., unpublished observations). Together, this likely reflects the specific requirement of generating a large number of diverse neuron subtypes, which eventually connect within a functional retinotopic map across several ganglia. For studies focusing on the assembly of the fly visual circuitry during development, the recent discoveries highlight that the mechanisms, by which neurons are generated in this part of the brain set the framework for how they connect. Moreover, they inform us about potential steps that axons and dendrites of newly born neurons need to accomplish. These all have to be genetically controlled by distinct guidance mechanisms and await discovery. Finally, the recent findings began to highlight unexpected similarities between neurogenesis in the Drosophila optic lobe and the vertebrate cerebral cortex and retina. Common features are for instance that (1) neurogenesis starts out in neuroepithelia (reviewed in: CitationBrand & Livesey, 2011) and (2) NE cells initially undergo symmetric, proliferative divisions and then switch to an asymmetric, differentiative division mode (reviewed in: CitationBreunig et al., 2011). Further similarities, which also extend to neurogenesis in the fly VNC and brain, include the temporal specification of neural progenitors and their offspring in the vertebrate CNS (reviewed in: CitationKohwi & Doe, 2013; CitationNaka et al., 2008). Moreover recent studies used the fly optic lobe to uncover the elusive function of the microcephaly protein Abnormal spindle protein (Asp) in neuroepithelial morphogenesis (CitationRujano et al., 2013). It is thus tempting to consider the optic lobe as a “mini-cortex” and a powerful genetic model for gaining novel insights into the molecular mechanisms that potentially also regulate cortical neural stem cell biology and neurogenesis in vertebrates.
ACKNOWLEDGEMENTS
We would like to thank K. F. Fischbach for encouraging us to contribute to this special issue with a review. We apologize to those of our colleagues, whose work was not included because of our specific focus on neurogenesis in the optic lobe. We are grateful to B. Egger, A. Gould, I.A. Meinertzhagen, and T. Tabata for advice, and K. Dolan, R. Kaschula, B. Richier, and N. Shimosako for critically reading this manuscript.
H.A. and I.S. are supported by the Medical Research Council (U117581332).
Declaration of interest: The authors report no conflict of interest. The authors alone are responsible for the content and writing of the paper.
REFERENCES
- Bayraktar, O. A., & Doe, C. Q. (2013). Combinatorial temporal patterning in progenitors expands neural diversity. Nature, 498, 449–455.
- Bazigou, E., Apitz, H., Johansson, J., Loren, C. E., Hirst, E. M., Chen, P. L., et al. (2007). Anterograde Jelly belly and Alk receptor tyrosine kinase signaling mediates retinal axon targeting in Drosophila. Cell, 128, 961–975.
- Bello, B. C., Izergina, N., Caussinus, E., & Reichert, H. (2008). Amplification of neural stem cell proliferation by intermediate progenitor cells in Drosophila brain development. Neural Dev, 3, 5.
- Bier, E., Vaessin, H., Younger-Shepherd, S., Jan, L. Y., & Jan, Y. N. (1992). deadpan, an essential pan-neural gene in Drosophila, encodes a helix-loop-helix protein similar to the hairy gene product. Genes Dev, 6, 2137–2151.
- Boone, J. Q., & Doe, C. Q. (2008). Identification of Drosophila type II neuroblast lineages containing transit amplifying ganglion mother cells. Dev Neurobiol, 68, 1185–1195.
- Bowman, S. K., Rolland, V., Betschinger, J., Kinsey, K. A., Emery, G., & Knoblich, J. A. (2008). The tumor suppressors Brat and Numb regulate transit-amplifying neuroblast lineages in Drosophila. Dev Cell, 14, 535–546.
- Braitenberg, V. (1967). Patterns of projection in the visual system of the fly. I. Retina-lamina projections. Exp Brain Res, 3, 271–298.
- Brand, A. H., & Livesey, F. J. (2011). Neural stem cell biology in vertebrates and invertebrates: more alike than different? Neuron, 70, 719–729.
- Brand, M., Jarman, A. P., Jan, L. Y., & Jan, Y. N. (1993). asense is a Drosophila neural precursor gene and is capable of initiating sense organ formation. Development, 119, 1–17.
- Breunig, J. J., Haydar, T. F., & Rakic, P. (2011). Neural stem cells: historical perspective and future prospects. Neuron, 70, 614–625.
- Britton, J. S., & Edgar, B. A. (1998). Environmental control of the cell cycle in Drosophila: nutrition activates mitotic and endoreplicative cells by distinct mechanisms. Development, 125, 2149–2158.
- Brody, T., & Odenwald, W. F. (2000). Programmed transformations in neuroblast gene expression during Drosophila CNS lineage development. Dev Biol, 226, 34–44.
- Cabrera, C. V., Martinez-Arias, A., & Bate, M. (1987). The expression of three members of the achaete-scute gene complex correlates with neuroblast segregation in Drosophila. Cell, 50, 425–433.
- Campos-Ortega, J. A. (1993). Early neurogenesis in Drosophila melanogaster. In M. Bate & A. Martinez Arias (Eds.), The development of Drosophila melanogaster (Vol. II, pp. 1091–1129): Cold Spring Harbor Laboratory Press.
- Cheyette, B. N., Green, P. J., Martin, K., Garren, H., Hartenstein, V., & Zipursky, S. L. (1994). The Drosophila sine oculis locus encodes a homeodomain-containing protein required for the development of the entire visual system. Neuron, 12, 977–996.
- Choksi, S. P., Southall, T. D., Bossing, T., Edoff, K., de Wit, E., Fischer, B. E., et al. (2006). Prospero acts as a binary switch between self-renewal and differentiation in Drosophila neural stem cells. Dev Cell, 11, 775–789.
- Chotard, C., Leung, W., & Salecker, I. (2005). glial cells missing and gcm2 cell autonomously regulate both glial and neuronal development in the visual system of Drosophila. Neuron, 48, 237–251.
- Chu, T., Chiu, M., Zhang, E., & Kunes, S. (2006). A C-terminal motif targets Hedgehog to axons, coordinating assembly of the Drosophila eye and brain. Dev Cell, 10, 635–646.
- Clandinin, T. R., & Feldheim, D. A. (2009). Making a visual map: mechanisms and molecules. Curr Opin Neurobiol, 19, 174–180.
- Clandinin, T. R., & Zipursky, S. L. (2000). Afferent growth cone interactions control synaptic specificity in the Drosophila visual system. Neuron, 28, 427–436.
- Colonques, J., Ceron, J., Reichert, H., & Tejedor, F. J. (2011). A transient expression of Prospero promotes cell cycle exit of Drosophila postembryonic neurons through the regulation of Dacapo. PLoS One, 6, e19342.
- Datta, S. (1995). Control of proliferation activation in quiescent neuroblasts of the Drosophila central nervous system. Development, 121, 1173–1182.
- Datta, S. (1999). Activation of neuroblast proliferation in explant culture of the Drosophila larval CNS. Brain Res, 818, 77–83.
- Dearborn, R., Jr., & Kunes, S. (2004). An axon scaffold induced by retinal axons directs glia to destinations in the Drosophila optic lobe. Development, 131, 2291–2303.
- Doe, C. Q., Chu-LaGraff, Q., Wright, D. M., & Scott, M. P. (1991). The prospero gene specifies cell fates in the Drosophila central nervous system. Cell, 65, 451–464.
- Ebens, A. J., Garren, H., Cheyette, B. N., & Zipursky, S. L. (1993). The Drosophila anachronism locus: a glycoprotein secreted by glia inhibits neuroblast proliferation. Cell, 74, 15–27.
- Egger, B., Boone, J. Q., Stevens, N. R., Brand, A. H., & Doe, C. Q. (2007). Regulation of spindle orientation and neural stem cell fate in the Drosophila optic lobe. Neural Dev, 2, 1.
- Egger, B., Gold, K. S., & Brand, A. H. (2010). Notch regulates the switch from symmetric to asymmetric neural stem cell division in the Drosophila optic lobe. Development, 137, 2981–2987.
- Egger, B., Gold, K. S., & Brand, A. H. (2011). Regulating the balance between symmetric and asymmetric stem cell division in the developing brain. Fly (Austin), 5, 237–241.
- Erclik, T., Hartenstein, V., Lipshitz, H. D., & McInnes, R. R. (2008). Conserved role of the Vsx genes supports a monophyletic origin for bilaterian visual systems. Curr Biol, 18, 1278–1287.
- Fernandez-Hernandez, I., Rhiner, C., & Moreno, E. (2013). Adult neurogenesis in Drosophila. Cell Rep, 3, 1857–1865.
- Fischbach, K. F. (1983). Neural cell types surviving congenital sensory deprivation in the optic lobes of Drosophila melanogaster. Dev Biol, 95, 1–18.
- Fischbach, K. F., & Dittrich, A. P. M. (1989). The optic lobe of Drosophila melanogaster. I. A Golgi analysis of wild-type structure. Cell Tissue Res, 258, 441–475.
- Fischbach, K. F., Linneweber, G. A., Andlauer, T. F., Hertenstein, A., Bonengel, B., & Chaudhary, K. (2009). The irre cell recognition module (IRM) proteins. J Neurogenet, 23, 48–67.
- Fischbach, K. F., & Technau, G. (1984). Cell degeneration in the developing optic lobes of the sine oculis and small- optic-lobes mutants of Drosophila melanogaster. Dev Biol, 104, 219–239.
- Freeman, M. (2008). Rhomboid proteases and their biological functions. Annu Rev Genet, 42, 191–210.
- Freeman, M. R., & Rowitch, D. H. (2013). Evolving concepts of gliogenesis: a look way back and ahead to the next 25 years. Neuron, 80, 613–623.
- Garcia-Bellido, A., & de Celis, J. F. (2009). The complex tale of the achaete-scute complex: a paradigmatic case in the analysis of gene organization and function during development. Genetics, 182, 631–639.
- Green, P., Hartenstein, A. Y., & Hartenstein, V. (1993). The embryonic development of the Drosophila visual system. Cell Tissue Res, 273, 583–598.
- Hadjieconomou, D., Timofeev, K., & Salecker, I. (2011). A step-by-step guide to visual circuit assembly in Drosophila. Curr Opin Neurobiol, 21, 76–84.
- Hasegawa, E., Kaido, M., Takayama, R., & Sato, M. (2013). Brain-specific-homeobox is required for the specification of neuronal types in the Drosophila optic lobe. Dev Biol, 377, 90–99.
- Hasegawa, E., Kitada, Y., Kaido, M., Takayama, R., Awasaki, T., Tabata, T., et al. (2011). Concentric zones, cell migration and neuronal circuits in the Drosophila visual center. Development, 138, 983–993.
- Hayden, M. A., Akong, K., & Peifer, M. (2007). Novel roles for APC family members and Wingless/Wnt signaling during Drosophila brain development. Dev Biol, 305, 358–376.
- Hofbauer, A., & Campos-Ortega, J. A. (1990). Proliferation and and early differentiation of the optic lobes in Drosophila melanogaster. Roux’s Arch Dev Biol, 198, 264–274.
- Homem, C. C., & Knoblich, J. A. (2012). Drosophila neuroblasts: a model for stem cell biology. Development, 139, 4297–4310.
- Huang, Z., & Kunes, S. (1996). Hedgehog, transmitted along retinal axons, triggers neurogenesis in the developing visual centers of the Drosophila brain. Cell, 86, 411–422.
- Huang, Z., & Kunes, S. (1998). Signals transmitted along retinal axons in Drosophila: Hedgehog signal reception and the cell circuitry of lamina cartridge assembly. Development, 125, 3753–3764.
- Huang, Z., Shilo, B. Z., & Kunes, S. (1998). A retinal axon fascicle uses spitz, an EGF receptor ligand, to construct a synaptic cartridge in the brain of Drosophila. Cell, 95, 693–703.
- Ikeshima-Kataoka, H., Skeath, J. B., Nabeshima, Y., Doe, C. Q., & Matsuzaki, F. (1997). Miranda directs Prospero to a daughter cell during Drosophila asymmetric divisions. Nature, 390, 625–629.
- Isshiki, T., Pearson, B., Holbrook, S., & Doe, C. Q. (2001). Drosophila neuroblasts sequentially express transcription factors which specify the temporal identity of their neuronal progeny. Cell, 106, 511–521.
- Ito, K., & Hotta, Y. (1992). Proliferation pattern of postembryonic neuroblasts in the brain of Drosophila melanogaster. Dev Biol, 149, 134–148.
- Ito, M., Masuda, N., Shinomiya, K., Endo, K., & Ito, K. (2013). Systematic analysis of neural projections reveals clonal composition of the Drosophila brain. Curr Biol, 23, 644–655.
- Jarman, A. P., Brand, M., Jan, L. Y., & Jan, Y. N. (1993). The regulation and function of the helix-loop-helix gene, asense, in Drosophila neural precursors. Development, 119, 19–29.
- Joesch, M., Plett, J., Borst, A., & Reiff, D. F. (2008). Response properties of motion-sensitive visual interneurons in the lobula plate of Drosophila melanogaster. Curr Biol, 18, 368–374.
- Kambadur, R., Koizumi, K., Stivers, C., Nagle, J., Poole, S. J., & Odenwald, W. F. (1998). Regulation of POU genes by castor and hunchback establishes layered compartments in the Drosophila CNS. Genes Dev, 12, 246–260.
- Kanai, M. I., Okabe, M., & Hiromi, Y. (2005). Seven-up controls switching of transcription factors that specify temporal identities of Drosophila neuroblasts. Dev Cell, 8, 203–213.
- Kaphingst, K., & Kunes, S. (1994). Pattern formation in the visual centers of the Drosophila brain: wingless acts via decapentaplegic to specify the dorsoventral axis. Cell, 78, 437–448.
- Kawamori, H., Tai, M., Sato, M., Yasugi, T., & Tabata, T. (2011). Fat/Hippo pathway regulates the progress of neural differentiation signaling in the Drosophila optic lobe. Dev Growth Differ, 53, 653–667.
- Kirschfeld, K. (1967). [The projection of the optical environment on the screen of the rhabdomere in the compound eye of the Musca]. Exp Brain Res, 3, 248–270.
- Kohwi, M., & Doe, C. Q. (2013). Temporal fate specification and neural progenitor competence during development. Nat Rev Neurosci, 14, 823–838.
- Kohwi, M., Lupton, J. R., Lai, S. L., Miller, M. R., & Doe, C. Q. (2013). Developmentally regulated subnuclear genome reorganization restricts neural progenitor competence in Drosophila. Cell, 152, 97–108.
- Kumar, J. P. (2010). Retinal determination the beginning of eye development. Curr Opin Neurobiol, 93, 1–28.
- Lanet, E., Gould, A. P., & Maurange, C. (2013). Protection of neuronal diversity at the expense of neuronal numbers during nutrient restriction in the Drosophila visual system. Cell Rep, 3, 587–594.
- Li, X., Chen, Z., & Desplan, C. (2013a). Temporal patterning of neural progenitors in Drosophila. Curr Top Dev Biol, 105, 69–96.
- Li, X., Erclik, T., Bertet, C., Chen, Z., Voutev, R., Venkatesh, S., et al. (2013b). Temporal patterning of Drosophila medulla neuroblasts controls neural fates. Nature, 498, 456–462.
- Li, Y., & Padgett, R. W. (2012). bantam is required for optic lobe development and glial cell proliferation. PLoS One, 7, e32910.
- Meinertzhagen, I. A., & Hanson, T. E. (1993). The development of the optic lobe. In M. Bate & A. Martinez Arias (Eds.), The development of Drosophila melanogaster (Vol. II, pp. 1363–1491): Cold Spring Harbor Laboratory Press
- Mettler, U., Vogler, G., & Urban, J. (2006). Timing of identity: spatiotemporal regulation of hunchback in neuroblast lineages of Drosophila by Seven-up and Prospero. Development, 133, 429–437.
- Morante, J., & Desplan, C. (2008). The color-vision circuit in the medulla of Drosophila. Curr Biol, 18, 553–565.
- Morante, J., Erclik, T., & Desplan, C. (2011). Cell migration in Drosophila optic lobe neurons is controlled by eyeless/Pax6. Development, 138, 687–693.
- Morante, J., Vallejo, D. M., Desplan, C., & Dominguez, M. (2013). Conserved miR-8/miR-200 defines a glial niche that controls neuroepithelial expansion and neuroblast transition. Dev Cell, 27, 174–187.
- Naka, H., Nakamura, S., Shimazaki, T., & Okano, H. (2008). Requirement for COUP-TFI and II in the temporal specification of neural stem cells in CNS development. Nat Neurosci, 11, 1014–1023.
- Nassif, C., Noveen, A., & Hartenstein, V. (2003). Early development of the Drosophila brain: III. The pattern of neuropile founder tracts during the larval period. J Comp Neurol, 455, 417–434.
- Ngo, K. T., Wang, J., Junker, M., Kriz, S., Vo, G., Asem, B., et al. (2010). Concomitant requirement for Notch and Jak/Stat signaling during neuro-epithelial differentiation in the Drosophila optic lobe. Dev Biol, 346, 284–295.
- O’Neill, E. M., Rebay, I., Tjian, R., & Rubin, G. M. (1994). The activities of two Ets-related transcription factors required for Drosophila eye development are modulated by the Ras/MAPK pathway. Cell, 78, 137–147.
- Orihara-Ono, M., Toriya, M., Nakao, K., & Okano, H. (2011). Downregulation of Notch mediates the seamless transition of individual Drosophila neuroepithelial progenitors into optic medullar neuroblasts during prolonged G1. Dev Biol, 351, 163–175.
- Otsuna, H., & Ito, K. (2006). Systematic analysis of the visual projection neurons of Drosophila melanogaster. I. Lobula-specific pathways. J Comp Neurol, 497, 928–958.
- Park, Y., Fujioka, M., Kobayashi, M., Jaynes, J. B., & Datta, S. (2001). even skipped is required to produce a trans-acting signal for larval neuroblast proliferation that can be mimicked by ecdysone. Development, 128, 1899–1909.
- Perez, S. E., & Steller, H. (1996). Migration of glial cells into retinal axon target field in Drosophila melanogaster. J Neurobiol, 30, 359–373.
- Perez-Gomez, R., Slovakova, J., Rives-Quinto, N., Krejci, A., & Carmena, A. (2013). A Serrate-Notch-Canoe complex mediates essential interactions between glia and neuroepithelial cells during Drosophila optic lobe development. J Cell Sci, 126, 4873–4884.
- Poeck, B., Fischer, S., Gunning, D., Zipursky, S. L., & Salecker, I. (2001). Glial cells mediate target layer selection of retinal axons in the developing visual system of Drosophila. Neuron, 29, 99–113.
- Power, M. E. (1943). The effect of reduction in numbers of ommatidia upon the brain of Drosophila melanogaster. J Exp Zool, 94, 33–71.
- Raghu, S. V., & Borst, A. (2011). Candidate glutamatergic neurons in the visual system of Drosophila. PLoS One, 6, e19472.
- Raghu, S. V., Claussen, J., & Borst, A. (2013). Neurons with GABAergic phenotype in the visual system of Drosophila. J Comp Neurol, 521, 252–265.
- Raghu, V. S., Reiff, D. F., & Borst, A. (2011). Neurons with cholinergic phenotype in the visual system of Drosophila. J Comp Neurol, 519, 162–176.
- Ramon y Cajal, S., & Sanchez y Sanchez, D. (1915). Contribucion al conociemento de los centros nerviosos de los insectos. Trabajos del Laboratorio de Investigaciones biológicas de la Universidad de Madrid.
- Reddy, B. V., Rauskolb, C., & Irvine, K. D. (2010). Influence of fat-hippo and notch signaling on the proliferation and differentiation of Drosophila optic neuroepithelia. Development, 137, 2397–2408.
- Robinow, S., & White, K. (1991). Characterization and spatial distribution of the ELAV protein during Drosophila melanogaster development. J Neurobiol, 22, 443–461.
- Rujano, M. A., Sanchez-Pulido, L., Pennetier, C., le Dez, G., & Basto, R. (2013). The microcephaly protein Asp regulates neuroepithelium morphogenesis by controlling the spatial distribution of myosin II. Nat Cell Biol, 15, 1294–1306.
- Sanes, J. R., & Zipursky, S. L. (2010). Design principles of insect and vertebrate visual systems. Neuron, 66, 15–36.
- Selleck, S. B., Gonzalez, C., Glover, D. M., & White, K. (1992). Regulation of the G1-S transition in postembryonic neuronal precursors by axon ingrowth. Nature, 355, 253–255.
- Selleck, S. B., & Steller, H. (1991). The influence of retinal innervation on neurogenesis in the first optic ganglion of Drosophila. Neuron, 6, 83–99.
- Song, Y., Chung, S., & Kunes, S. (2000). Combgap relays wingless signal reception to the determination of cortical cell fate in the Drosophila visual system. Mol Cell, 6, 1143–1154.
- Sousa-Nunes, R., Cheng, L. Y., & Gould, A. P. (2010). Regulating neural proliferation in the Drosophila CNS. Curr Opin Neurobiol, 20, 50–57.
- Strausfeld, N. J. (2005). The evolution of crustacean and insect optic lobes and the origins of chiasmata. Arthropod Struct Dev, 34, 235–256.
- Strausfeld, N. J., Sinakevitch, I., & Okamura, J. Y. (2007). Organization of local interneurons in optic glomeruli of the dipterous visual system and comparisons with the antennal lobes. Dev Neurobiol, 67, 1267–1288.
- Sugie, A., Umetsu, D., Yasugi, T., Fischbach, K. F., & Tabata, T. (2010). Recognition of pre- and postsynaptic neurons via nephrin/NEPH1 homologs is a basis for the formation of the Drosophila retinotopic map. Development, 137, 3303–3313.
- Suzuki, T., Kaido, M., Takayama, R., & Sato, M. (2013). A temporal mechanism that produces neuronal diversity in the Drosophila visual center. Dev Biol, 380, 12–24.
- Takemura, S. Y., Bharioke, A., Lu, Z., Nern, A., Vitaladevuni, S., Rivlin, P. K., et al. (2013). A visual motion detection circuit suggested by Drosophila connectomics. Nature, 500, 175–181.
- Ting, C. Y., McQueen, P. G., Pandya, N., Lin, T. Y., Yang, M., Reddy, O. V., et al. (2014). Photoreceptor-derived activin promotes dendritic termination and restricts the receptive fields of first-order interneurons in Drosophila. Neuron, 81, 830–846.
- Togane, Y., Ayukawa, R., Hara, Y., Akagawa, H., Iwabuchi, K., & Tsujimura, H. (2012). Spatio-temporal pattern of programmed cell death in the developing Drosophila optic lobe. Dev Growth Differ, 54, 503–518.
- Treisman, J. E. (2013). Retinal differentiation in Drosophila. Wiley Interdiscip Rev Dev Biol, 2, 545–557.
- Truman, J. W., Schuppe, H., Shepherd, D., & Williams, D. W. (2004). Developmental architecture of adult-specific lineages in the ventral CNS of Drosophila. Development, 131, 5167–5184.
- Tuthill, J. C., Nern, A., Holtz, S. L., Rubin, G. M., & Reiser, M. B. (2013). Contributions of the 12 neuron classes in the fly lamina to motion vision. Neuron, 79, 128–140.
- Umetsu, D., Murakami, S., Sato, M., & Tabata, T. (2006). The highly ordered assembly of retinal axons and their synaptic partners is regulated by Hedgehog/Single-minded in the Drosophila visual system. Development, 133, 791–800.
- Urbach, R., & Technau, G. M. (2003). Molecular markers for identified neuroblasts in the developing brain of Drosophila. Development, 130, 3621–3637.
- Urbach, R., & Technau, G. M. (2004). Neuroblast formation and patterning during early brain development in Drosophila. BioEssays, 26, 739–751.
- Viallanes, H. (1883). Recherches sur l’histologie des insectes et sur les phenomenes histologiques qui accompagnent le developpment postembryonnaire de ces animaux. Annales des Sciences Naturelles (Zoologie et Biologie Animale), ser 6, 14, 1–348.
- Voigt, A., Pflanz, R., Schafer, U., & Jackle, H. (2002). Perlecan participates in proliferation activation of quiescent Drosophila neuroblasts. Dev Dyn, 224, 403–412.
- Wallace, K., Liu, T. H., & Vaessin, H. (2000). The pan-neural bHLH proteins DEADPAN and ASENSE regulate mitotic activity and cdk inhibitor dacapo expression in the Drosophila larval optic lobes. Genesis, 26, 77–85.
- Wang, H., Chen, X., He, T., Zhou, Y., & Luo, H. (2013). Evidence for tissue-specific Jak/STAT target genes in Drosophila optic lobe development. Genetics, 195, 1291–1306.
- Wang, W., Li, Y., Zhou, L., Yue, H., & Luo, H. (2011a). Role of JAK/STAT signaling in neuroepithelial stem cell maintenance and proliferation in the Drosophila optic lobe. Biochem Biophys Res Commun, 410, 714–720.
- Wang, W., Liu, W., Wang, Y., Zhou, L., Tang, X., & Luo, H. (2011b). Notch signaling regulates neuroepithelial stem cell maintenance and neuroblast formation in Drosophila optic lobe development. Dev Biol, 350, 414–428.
- Weng, M., Haenfler, J. M., & Lee, C. Y. (2012). Changes in Notch signaling coordinates maintenance and differentiation of the Drosophila larval optic lobe neuroepithelia. Dev Neurobiol, 72, 1376–1390.
- White, K., Grether, M. E., Abrams, J. M., Young, L., Farrell, K., & Steller, H. (1994). Genetic control of programmed cell death in Drosophila. Science, 264, 677–683.
- White, K., & Kankel, D. R. (1978). Patterns of cell division and cell movement in the formation of the imaginal nervous system in Drosophila melanogaster. Dev Biol, 65, 296–321.
- Wolff, T., & Ready, D. F. (1993). Pattern formation in the Drosophila retina. In M. Bate & A. Martinez-Arias (Eds.), The development of Drosophila melanogaster (Vol. II, pp. 1277–1325). Cold Spring Harbor: Cold Spring Harbor Laboratory Press.
- Yamanaka, N., Rewitz, K. F., & O’Connor, M. B. (2013). Ecdysone control of developmental transitions: lessons from Drosophila research. Annu Rev Entomol, 58, 497–516.
- Yasugi, T., Sugie, A., Umetsu, D., & Tabata, T. (2010). Coordinated sequential action of EGFR and Notch signaling pathways regulates proneural wave progression in the Drosophila optic lobe. Development, 137, 3193–3203.
- Yasugi, T., Umetsu, D., Murakami, S., Sato, M., & Tabata, T. (2008). Drosophila optic lobe neuroblasts triggered by a wave of proneural gene expression that is negatively regulated by JAK/STAT. Development, 135, 1471–1480.
- Yogev, S., Schejter, E. D., & Shilo, B. Z. (2010). Polarized secretion of Drosophila EGFR ligand from photoreceptor neurons is controlled by ER localization of the ligand-processing machinery. PLoS Biol, 8, e1000505.
- Yu, H. H., Awasaki, T., Schroeder, M. D., Long, F., Yang, J. S., He, Y., et al. (2013). Clonal development and organization of the adult Drosophila central brain. Curr Biol, 23, 633–643.
- Zhao, B., Tumaneng, K., & Guan, K. L. (2011). The Hippo pathway in organ size control, tissue regeneration and stem cell self-renewal. Nat Cell Biol, 13, 877–883.
- Zhou, L., & Luo, H. (2013). Replication protein a links cell cycle progression and the onset of neurogenesis in Drosophila optic lobe development. J Neurosci, 33, 2873–2888.
- Zhu, C. C., Boone, J. Q., Jensen, P. A., Hanna, S., Podemski, L., Locke, J., et al. (2008). Drosophila Activin- and the Activin-like product Dawdle function redundantly to regulate proliferation in the larval brain. Development, 135, 513–521.