
Abstract
Angiogenesis, or neovascularization, refers to development of new vessels from pre-existing vasculature. Retinal and choroidal neovascularization leads to oedema, haemorrhages, and fibrosis, causing visual impairment and blindness. In multiple studies, vascular endothelial growth factor (VEGF) has been shown to be the most important factor in ocular angiogenesis. Recently discovered anti-VEGF treatments have revolutionized the therapy of neovascular diseases in the eye. These agents have been shown not just to stop the angiogenic process and maintain visual acuity but also improve vision in a great proportion of patients at least during a 2-year follow-up. However, there are also problems with these agents and their delivery regimens, and new therapeutic strategies are needed. This review summarizes the most important growth factors participating in the angiogenic process in the retina and the choroid, diseases where angiogenesis plays the most devastating part causing visual impairment, as well as current antiangiogenic treatments for these diseases.
| Abbreviations | ||
| AGE | = | advanced glycation end-products |
| AMD | = | age-related macular degeneration |
| ANG | = | angiopoietin |
| bFGF | = | basic fibroblast growth factor |
| CNV | = | choroidal neovascularization |
| EMA | = | European Medicines Agency |
| FAG | = | fluorescein angiography |
| FDA | = | Food and Drug Administration |
| HIF-1α | = | hypoxia-inducible factor 1α |
| IGF-I | = | insulin-like growth factor I |
| IRMA | = | intraretinal microvascular abnormalities |
| Nrp | = | neuropilin |
| OCT | = | ocular coherence tomography |
| PDGF | = | platelet-derived growth factor |
| PDT | = | photodynamic therapy |
| PKC | = | protein kinase C |
| PlGF | = | placental growth factor |
| RISC | = | RNA-induced silencing complex |
| ROP | = | retinopathy of prematurity |
| ROS | = | reactive oxygen species |
| RPE | = | retinal pigment epithelium |
| VEGF | = | vascular endothelial growth factor |
| VEGFR | = | vascular endothelial growth factor receptor |
Key messages
Vascular endothelial growth factor is the most important factor in ocular angiogenesis.
Antiangiogenic treatments have revolutionized the therapy of neovascular diseases in the eye.
There are also problems with current antiangiogenic agents and their delivery regimens, and new therapeutic strategies are needed.
Introduction
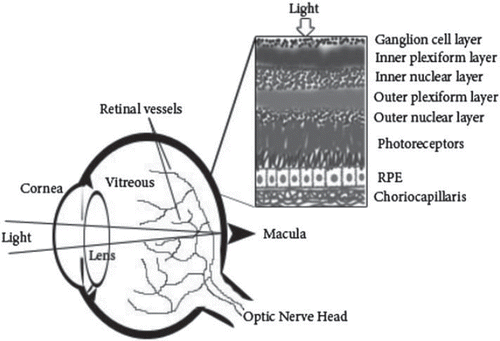
The human eye is an organ composed of many sections important to normal visual acuity. The cornea, lens, vitreous, and outer retina are unique avascular structures allowing light to enter the eye. The light rays focused by the cornea and the lens form an image on the retina, the light-sensing portion of the eye. The photoreceptor cells in the retina convert the image into an electrical signal that travels down the optic nerve to the brain (). Optical transparency of the eye is needed for normal visual function. On the other hand, the adult retina is a neural tissue with high metabolism and the highest oxygen consumption per unit weight of all human tissues. Therefore, the choroid, the most vascular portion of the eye also nourishing the retina, has one of the highest blood-flow rates in the body, 800–1000 mL/100 g tissue/min (Citation1). In healthy adults this delicate ocular vascular system is maintained and controlled by the balance between the angiogenic factors and angiogenic inhibitors (Citation2).
Figure 1. Normal anatomy of the eye.
Diabetic retinopathy and age-related macular degeneration (AMD) are the leading causes of visual impairment in developed countries (). Diabetes mellitus affects over 170 million people worldwide, and the estimated number of patients in the year 2030 is 366 million. The prevalence of diabetes is increasing particularly in low- and middle-income countries and in the working-age population. A total of 75% of both type 1 and type 2 diabetics will have diabetic retinopathy after 20 years of disease (Citation3). The incidence and prevalence of AMD increase with advancing age and will rise as the population of older than 65 years increases. In a combined analysis of population-based eye disease prevalence data, AMD was present in 0.2% of the population aged 55–64 years, rising to 13% of the population older than 85 years (Citation4).
Table I. Statistics of visual impairment in developed countries and the world. Modified from WHO global data on visual impairment in the year 2002. Sample of Western countries includes Australia, Denmark, Finland, Iceland, Ireland, Italy, Netherlands, UK, and USA.
Vascular endothelial growth factors
The first and the most potent member of the vascular endothelial growth factor (VEGF) family, VEGF (also called VEGF-A), was identified in highly vascularized tumours in 1983 (Citation5). After the discovery of VEGF, four other members in the human VEGF family have been identified: VEGF-B, VEGF-C, VEGF-D, and placental growth factor (PlGF) (Citation6–9). In addition to these VEGFs, viral VEGF homologues (VEGF-E) and snake venom VEGFs (VEGF-F) have been found (Citation10,Citation11). VEGF is a 46 kDa homodimeric glycoprotein with several isoforms including VEGF121, VEGF145, VEGF165, VEGF189, and VEGF206 generated by alternative mRNA splicing from the same gene (Citation12). The corresponding rodent isoforms have one amino acid less than human proteins. VEGF is a potent angiogenic stimulator, promoting proliferation, migration, proteolytic activity, and capillary tube formation of endothelial cells, playing a crucial role in both normal and pathological angiogenesis. It can increase vascular permeability, and its synthesis is strongly up-regulated by hypoxia (Citation13). In addition, it plays a major role in embryogenesis, thus heterozygous deletion of the VEGF gene results in embryo death between days 8.5 and 9.5 (Citation14).
In the retina, VEGF is produced by multiple cell types, including the retinal pigment epithelium (RPE), pericytes, endothelial cells, ganglion cells, and Müller cells—a specific type of glial cells found in the inner layers of retina (Citation15,Citation16). RPE and Müller cells are the major sources and endothelial cells the primary target of VEGF in the retina. VEGF levels in the ocular fluid are very low in the normal human eye. However, intravitreal levels of VEGF were elevated in proliferative diabetic retinopathy (Citation17). In addition, significantly elevated VEGF levels in the aqueous humour have been reported in diabetic patients with macular oedema (Citation18). High levels of VEGF are also found in neovascular membranes excised from patients with AMD and in the vitreous humour of a baby with retinopathy of prematurity (ROP) (Citation19,Citation20).
Different VEGF isoforms may have different functions in ocular diseases. In normal adult monkey and mouse eyes, the most abundant isoforms are VEGF120 and VEGF164 (Citation21). Furthermore, VEGF164 has been shown to be the most important isoform in the blood–retina barrier break-down and pathologic intraocular neovascularization (Citation22). In addition, VEGF120 is the main isoform expressed in mouse choroidal neovascular membranes (Citation23). On the other hand, VEGF188 is almost completely absent in adult mouse retina (Citation24). VEGF has also been shown to have survival function in the retina. Intravitreal VEGF164 and VEGF120 displayed a protective effect on apoptotic retinal cells in a rat retinal ischaemia–reperfusion model. This protective effect was not inhibited by blockage of VEGF164, suggesting that VEGF isoforms may differ in their neuroprotection properties (Citation25). Specific VEGF isoform targeting would therefore be beneficial in blocking the vessel growth and sparing the neuroprotective effects at the same time. In addition, VEGF acts as a survival factor for newly formed blood vessels in the retina (Citation26). Furthermore, development of the choriocapillaris is dependent on VEGF produced by RPE (Citation27). VEGF expressed by RPE has a role in the maintenance of the adult choriocapillaris (Citation21). Long-term therapeutic neutralization of VEGF may thus lead to unexpected degeneration of the choroidal circulation (Citation28).
Also other members of the VEGF family stimulate neovascularization and excessive vascular permeability. PlGF, VEGF-B, VEGF-C, and VEGF-D are all endothelial cell mitogens in vitro and in vivo, but their role in ocular angiogenesis remains unclear (Citation6,Citation9). Mice deficient in VEGF-B have normal retinal vascular development, and there is no difference in hypoxia-induced retinal neovascularization compared to wild-type mice, suggesting that VEGF-B does not play a role in retinal neovascularization (Citation29). VEGF-C and VEGF-D are produced as prepropeptides and further processed to biologically fully active forms (Citation9,Citation30). Both VEGF-C and VEGF-D are involved primarily in lymphangiogenesis but also in angiogenesis (Citation31,Citation32). VEGF-C and VEGF-D have been found in subretinal vascular membranes of AMD patients as well as in neovascular samples of diabetic patients, implying a role also in ocular angiogenesis (Citation33,Citation34). It has been shown that cell–cell adhesion and cell attachment independently regulate VEGF-D mRNA expression in human RPE in vitro, suggesting that the break-down of these interactions causes the over-expression of VEGF-D (Citation33). Hypoxic human RPE cells have been shown to produce PlGF, and PlGF has also been directly implicated in proliferative retinopathy (Citation35,Citation36).
VEGF receptors
VEGF exerts its biological activities through multiple receptors: vascular endothelial growth factor receptor (VEGFR)-1 (Flt-1), VEGFR-2 (Flk-1/KDR), and VEGFR-3 (Flt-4), which are expressed predominantly in endothelial cells, and to a lesser extent on monocytes and macrophages (Citation37). The binding of VEGF to its receptors initiates a signal transduction cascade mediating vascular permeability and endothelial cell proliferation and migration. In addition, two co-receptors for VEGF, neuropilin-1 and 2 (Nrp-1 and Nrp-2), have been described (Citation38). VEGFR-1 binds VEGF, VEGF-B, and PlGF, while VEGFR-2 binds VEGF and proteolytically processed forms of VEGF-C and -D. VEGFR-3 binds all forms of VEGF-C and -D, mediating lymphangiogenesis (). Homozygous VEGFR-1 deletion permits endothelial cell differentiation, but the forming vascular channels are grossly abnormal leading to death of the animals in utero (Citation39). Homozygous deletion of VEGFR-2 leads to death of mice between embryonic days 8.5 and 9.5 (Citation40). Normal human choriocapillaris expresses VEGFR-1 and VEGFR-2. Furthermore, VEGFR-3 is present in the choriocapillaris endothelium next to the RPE layer (Citation41). VEGFR-2 is the major mediator of mitogenesis of endothelial cells. Through VEGFR-1, VEGF has been shown to promote assembly of endothelial cells into tubes. However, signalling via VEGFR-1 is ligand-dependent; it may enhance proangiogenic effects via indirect VEGFR-2 activation or act as a negative modulator of angiogenesis induced by VEGFR-2 signalling (Citation42). VEGF also interacts with neuropilins. A recent study by Lahteenvuo et al. also showed that VEGF-B186 leads to activation of Nrp-1 and consequent activation of G-protein-mediated signalling (Citation43). In early development, Nrp-1 is expressed in arteries, and Nrp-2 is expressed in veins (Citation44). The absence of either Nrp-1 or -2 has been shown to suppress ocular neovascularization (Citation45,Citation46). Nrp-1 is involved in retinal neovascularization in diabetic retinopathy and in ischaemic animal models. Co-expression of VEGFR-2 and Nrp-2 was suggested to facilitate fibrovascular proliferation in diabetic retinopathy (Citation47).
Figure 2. VEGF family and VEGF receptors.
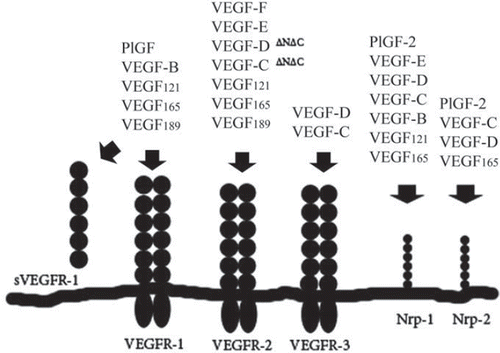
Soluble (s)Flt-1 is a naturally occurring protein antagonist of VEGF formed by alternative splicing of the pre-mRNA for the full-length VEGFR-1. The angiostatic activity of sFlt-1 results from inhibition of VEGF by two mechanisms. It causes both sequestration of VEGF to which it binds with high affinity and forms inactive heterodimers with VEGFR-1 and VEGFR-2 (Citation48,Citation49). It is not clear whether sFlt-1 has a role in normal eye, but several studies have shown that over-expression of sFlt-1 results in corneal avascularity in animal models (Citation50).
Retinal and choroidal neovascular diseases
Retinal neovascularization is defined as a state where new pathologic vessels originate from the existing retinal veins and extend along the inner surface of the retina. On the other hand, choroidal neovascularization (CNV) originates from the choroid and grows through a break in Bruch's membrane beneath the RPE or the retina. Virtually any pathologic process that involves the RPE and damages Bruch's membrane can be complicated by CNV. This review focuses on the most common diseases where either retinal or choroidal neovascularization plays the most devastating part increasing the risk of visual impairment.
Diabetic retinopathy
Clinical signs. Diabetic retinopathy begins as mild, non-proliferative abnormalities and progresses to moderate and severe non-proliferative diabetic retinopathy () and proliferative diabetic retinopathy (). Macular oedema can develop at any time in the progression of diabetic retinopathy. Clinical features of non-proliferative diabetic retinopathy include microaneurysms, intraretinal haemorrhages, soft and hard exudates, venous tortuosity or beading, and intraretinal microvascular abnormalities (IRMA). Microaneurysms, focal dilations of retinal capillaries, are early signs of diabetic retinopathy. Hypoxia caused by capillary dysregulation provokes growth factor expression and nerve fibre layer infarctions creating soft exudates, which are visualized as grey or white lesions and are the result of stasis of axoplasmic flow. Growth factors cause induction of fenestrations in the vascular endothelium and dissolution of tight junctions leading to accumulations of intraretinal plasma proteins and hard exudates. Increased venous tortuosity and beading is caused by dysregulation of the capillaries. Prolonged occlusion of capillaries causes hypoxia, which leads to the development of IRMA, additional routes for blood through arteries to veins.
Figure 3. A: Moderate non-proliferative diabetic retinopathy with hard exudates (large arrows), soft exudates (small arrows), and microaneurysms (arrowheads). B: Proliferative diabetic retinopathy and neovascularization in the optic disc.
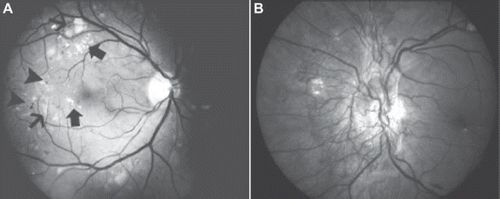
Diabetic retinopathy can be divided into different stages depending on the severity of the disease. Microaneurysms in the retina refer to mild non-proliferative diabetic retinopathy. In moderate non-proliferative diabetic retinopathy there are more changes than microaneurysms, including soft and hard exudates. Severe non-proliferative diabetic retinopathy is defined as more than 20 intraretinal haemorrhages/microaneurysms in each of four quadrants, definite venous beading in at least two quadrants, or prominent IRMA with no signs of proliferative diabetic retinopathy in any quadrant in the retina.
Proliferative disease is diagnosed in the case of neovascularization or preretinal or vitreous haemorrhage in the eye. New vessels traverse the inner limiting membrane and grow into the vitreous. Neovessels are fragile and tend to bleed, causing vitreous haemorrhages. Fibrous scar formation can result, with accompanying tractional retinal detachment, leading to blindness if left untreated. The foveal region is especially susceptible to hypoxia because of the lack of retinal vessels in this area. Diabetic maculopathy is characterized by an increase in vascular permeability, which results from a break-down of the inner blood–retinal barrier and leads to leakage of plasma components like proteins and lipids to the intraretinal space. Clinically significant macular oedema occurs if there is thickening of the retina involving the area within 500 μm of fovea, if there are hard exudates within 500 μm of the fovea with thickening of the adjacent retina, or if there is a zone of retinal thickening one disc area or larger in size, any part of which is within one disc diameter of the fovea. Changes in the macular area cause metamorphopsia, micropsia, decreased colour vision and contrast sensitivity, and are the main reasons for moderate visual loss in diabetic patients (Citation51).
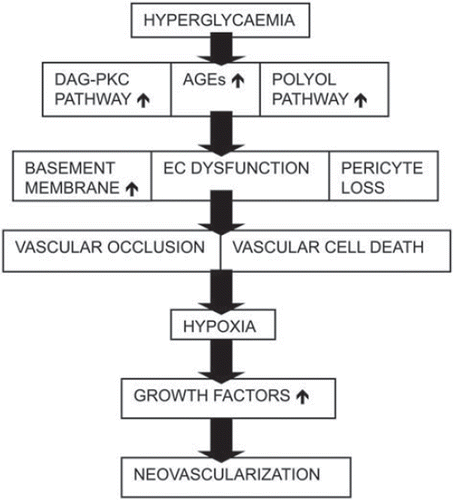
Pathophysiology. Preretinal neovascularization and chronic retinal oedema in the macular area are the two major causes for visual loss in diabetic retinopathy. Hyperglycaemia induces alterations in the pericytes and vascular endothelial cells in the retina, activating a cascade of signalling pathways leading to cellular dysfunction and eventually death. Histological lesions in diabetic retinopathy include capillary basement membrane thickening, endothelial cell dysfunction and loss, increased deposition of extracellular matrix components, pericyte loss, and leucocyte adhesion to the vessel wall (Citation52,Citation53). Dysfunction in the endothelial cells disrupts autoregulation of retinal blood-flow, disturbing the nutrition and oxygenation of the retina in different situations of ocular perfusion and intraocular pressures. Pericytes are responsible for the control of growth and survival of endothelial cells, especially under stress conditions (Citation54). Pericytes are very sensitive to disruptions of the extracellular environment. The level of apoptosis is much higher in pericytes than in endothelial cells under the same amount of glucose concentration fluctuations and diabetic conditions (Citation55). Inflammation plays an important role in the degeneration of retinal capillaries in diabetic patients. NFκB is a widely expressed inducible transcription factor that is an important regulator of many genes involved in the inflammatory and immune responses, proliferation, and apoptosis. NFκB is activated in retinal endothelial cells and pericytes exposed to hyperglycaemia and in the retinas of diabetic rats (Citation56,Citation57). Biochemical abnormalities related to diabetes include increased polyol pathway flux, resulting in elevated levels of intracellular sorbitol. This disrupts the osmotic balance and results in cellular damage (Citation58). In hyperglycaemia, carbohydrates interact with protein side chains to form advanced glycation end-products (AGEs). AGEs may affect such functions as enzyme activity and susceptibility of proteins to proteolysis (Citation59). Production of reactive oxygen species (ROS) has also been implicated in the development of diabetic complications. Diabetes may cause ROS production through glucose auto-oxidation and increased flux through the polyol pathway (Citation60). Experimental studies have shown that protein kinase C (PKC) activity, especially isoforms PKC-β and -δ, and levels of an activator of PKC, diacylglycerol, are increased by hyperglycaemia or oxidative stress (Citation61,Citation62). The PKC-β inhibitor ruboxistaurin has been studied in animals and clinical trials in patients with diabetic retinopathy. The trials have demonstrated a reduction in visual loss and need for laser treatment in patients with moderate to severe diabetic retinopathy over a 3-year follow-up (Citation63).
Extensive loss of retinal microvasculature leads to retinal hypoperfusion, ischaemia, and subsequent tissue hypoxia. Ocular neovascularization is strongly associated with retinal ischaemia, and multiple growth factors up-regulated by hypoxia have been implicated in its pathogenesis. The ischaemic retina secretes growth factors that stimulate pre-existing vessels to proliferate. The most important of these factors are VEGF, basic fibroblast growth factor (bFGF), insulin-like growth factor-I (IGF-I), platelet-derived growth factor (PDGF), and angiopoietins (ANG) (Citation64). Histologic studies have demonstrated the presence of growth factors and their receptors in the preretinal membranes and the vitreous humour of patients with proliferative diabetic retinopathy (Citation17). These factors have also been shown to correlate with the neovascular activity. VEGF expression was found to be up-regulated in rat vessels soon after the induction of experimental diabetes suggesting that VEGF is implicated in the vascular hyperpermeability seen in early diabetic retinopathy () (Citation65).
Figure 4. Consequences of hyperglycaemia.
Retinal vein occlusion
Retinal vein occlusion, occurring in branch or central vein, is the second most common retinal vascular disease after diabetic retinopathy. Central retinal vein occlusion is caused by a thrombosis near lamina cribrosa in the optic nerve head. On the other hand, branch retinal vein occlusion is usually due to a constriction in the site where an arteriosclerotic arteriole crosses and constricts the underlying vein. Branch retinal vein occlusion is three times more common than central retinal vein occlusion. Subsequent events in both cases are reduced blood-flow leading to hypoxia and subsequent macular oedema. Macular oedema is the major cause for visual loss in retinal vein occlusion patients, but also retinal neovascularization occurs in 36% of eyes with an area of non-perfusion greater than five discs diameter (Citation66). The clinical picture of retinal vein occlusion has classical features. It can be asymptomatic or with visual blurring usually involving the sector of visual field corresponding to the area of the retina involved. In macular or central retinal vein occlusion, there is always a visual disturbance. Acute occlusion presents characteristic clinical features with flame-shaped, dot and blot haemorrhage, soft and hard exudates, retinal oedema, and dilated, tortuous veins. Vascular sheathing and venous collaterals are signs of old occlusion. The diagnosis is based on clinical examination under biomicroscopy. Fluorescein angiography (FAG) can be used in determining the extent of macular oedema and ischaemia. Vascular occlusion and subsequent retinal ischaemia induces VEGF production leading to break-down of the blood–retina barrier and increased vascular permeability as well as neovessel growth. The VEGF mRNA expression is up-regulated in experimental retinal vein occlusion in animal models and in human retina with central retinal vein occlusion (Citation67,Citation68). In addition, intravitreal VEGF levels have been shown to be elevated in patients with retinal vein occlusion (Citation69). Current treatment options focus on the complications of the occlusion, such as macular oedema, retinal neovascularization, vitreous haemorrhage, and traction retinal detachment.
Retinopathy of prematurity
ROP is a disease in premature babies characterized by incomplete vascularization of the peripheral retina leading to retinal neovascularization (Citation70). ROP is the leading cause of blindness in childhood in the world. Low birth-weight, low gestational age, and supplemental oxygen therapy following delivery have been consistently associated with ROP (Citation71). The International classification of ROP describes the location and the extent of the developing vasculature, and the progressive staging of the disease. Stage 1 is the least severe, with stages 4 and 5 referring to partial and total retinal detachment. The term ‘plus’ disease indicates signs of on-going ROP activity that can accompany any stage of ROP. ‘Threshold’ ROP describes a prognostically relevant level of disease severity, indicating an increased likelihood of progression to retinal detachment (Citation72).
Animal experiments subsequently revealed that high levels of systemic oxygen resulted in permanent obliteration of blood vessels in the developing neonatal retina (Citation73). In ROP, the normal angiogenesis of the retina is disturbed by the relative hyperoxia of the extrauterine environment. This causes vaso-obliteration and non-vascularization of some areas of the anterior retina. The subsequent hypoxia causes a second chronic phase, characterized by the proliferation of vascular and glial cells and arteriovenous shunt formation, occasionally leading to involution or permanent cicatricial changes and visual impairment (Citation74). In experimental animal model, VEGF levels have been shown to rise resulting in neovascularization (Citation75). This finding was confirmed with elevated VEGF levels in the vitreous of an ROP patient (Citation19). However, VEGF is also involved in normal vascular development of the retina (Citation26). Current therapy consists of monitoring oxygen supplementation, aggressive screening in children who are at risk of having ROP, and laser treatment of the retina once threshold disease is reached.
Age-related macular degeneration
Clinical signs. AMD can develop to either a dry form with RPE and photoreceptor atrophy, or a wet form with CNV. In the dry AMD, RPE and therefore also photoreceptors in the macular area gradually disappear, and finally large scars of retinal atrophy develop. Geographic atrophy leads to gradual progression of visual loss, most likely because photoreceptors overlying areas of RPE atrophy are metabolically dependent on RPE cells (Citation76). Marked apoptosis of the outer nuclear layer and the inner nuclear layer of the retina is found near areas of RPE atrophy (Citation77). The wet AMD is responsible for 90% of cases of severe visual loss in AMD patients (Citation78). In the wet AMD, CNV causes subretinal and intraretinal accumulation of plasma and its components and haemorrhages (). Vision loss occurs through the structural and metabolic damages caused by exudates and haemorrhages leading to cell death. Metamorphopsia is a common symptom. Sustained leakage of blood, serum, and lipid stimulates a disciform scar formation, the end-stage of AMD. The disease starts with subretinal drusen classified morphologically either as hard or soft. Soft drusen have a tendency to become confluent, and multiple soft drusen are an independent risk factor for visual loss from AMD (Citation79). In addition, they can lead to deficits in colour contrast sensitivity and central visual field sensitivity (Citation80,Citation81). Immunohistochemical analyses have shown several agents, such as apolipoproteins B and E, complement C5 and C5b-9 terminal complexes, fibrinogen, and vitronectin to be present in drusen (Citation82,Citation83). These agents suggest a role of immunological and inflammatory processes in drusen pathogenesis and AMD.
Figure 5. Wet form of age-related macular degeneration with oedema, CNV (arrows), and haemorrhages (arrowheads). A: Fundus photograph. B: Fluorescein angiography image in the 30 s time point.
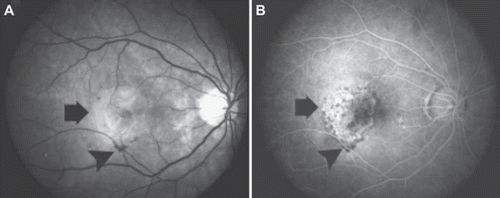
The diagnosis of AMD is made with FAG and ocular coherence tomography (OCT). In dry AMD there is no leakage or oedema but atrophy in the macula. The earliest signs of CNV are subretinal or sub-RPE oedema and/or haemorrhages appearing as a greenish grey subretinal tissue in ocular examination. Angiographic leakage patterns of neovascularization are classified either as classic or occult. The former refers to discrete areas that hyperfluoresce early and continue to exhibit progressive leakage with increasing intensity and extent during the examination. Occult neovascularization refers either to a fibrovascular RPE detachment with elevation of the RPE, hyperfluorescence and late leakage, or to late leakage of undetermined origin. In an OCT image classic CNV may appear as a highly reflective thickening between the retina and the hyperreflective external band that corresponds to the RPE/choroid complex. In addition, intraretinal oedema is present. Occult CNV can appear as an elevation of the RPE or as an irregularity of the external hyperreflective band. In geographic atrophy associated with the dry AMD, soft drusen may be identified as elevations of the RPE. Furthermore, atrophy is seen as thinning of the retina with an increased reflectivity of the choroid caused by the lack of pigment in the RPE (Citation84).
Pathophysiology. Vascular risk factors, smoking, age, race, and family history have been shown to be the most important pathogenetic factors for the development of AMD (Citation85). Genetic predisposition has been demonstrated by familial aggregation studies and twin studies. The chromosomes most commonly implicated are 1q25-31 and 10q26. In particular, variants in the gene for the complement factor H (CFH) and the genes PLEKHA1/LOC387715, Factor B, and complement components 2 and 3 (C2 and C3) have been implicated as major risk or protective factors for the development of AMD. An early pathological change in AMD is the appearance of two distinct types of deposits. Basal laminar deposits consist of membrano-granular material between the plasma membrane and basal lamina of the RPE. Basal linear deposits consist of vesicular material located in the inner collagenous zone of Bruch's membrane. Basal linear deposits may be more specific to AMD than basal laminar deposits (Citation86). Soft drusen can represent focal accentuations of basal linear deposits and a localized accumulation of basal laminar deposits (Citation87).
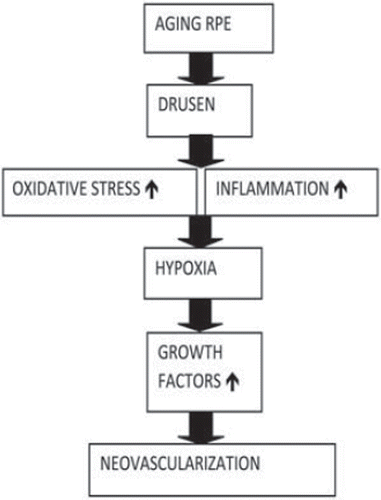
The RPE serves a variety of metabolic and supportive functions that are of vital importance for retinal photoreceptors, including maintenance of the blood–retina barrier, participation in the visual cycle, and phagocytic uptake and degradation of constantly shed apical photoreceptor outer segments (Citation88). Furthermore, in addition to many other growth factors the RPE produces VEGF in vivo under physiologic conditions to maintain the fenestrated choriocapillaris endothelium (Citation21). One of the reasons for the RPE dysfunction is an age-dependent phagocytic and metabolic insufficiency of postmitotic RPE cells. Impaired phagocytosis of photoreceptor outer segments by the RPE leads to a progressive accumulation of lipofuscin granules, a diverse group of autofluorescent lipid and protein aggregates, in the RPE (Citation89). Lipofuscin accumulation in the macula reduces RPE phagocytic capacity and increases with age (Citation90,Citation91). In addition, lipofuscin disrupts RPE function by mechanical distortion of cellular architecture and potentiating phototoxicity. Oxidative stress has a significant role in the pathogenesis of AMD. In vitro studies show that RPE lipofuscin is a photo-inducible generator of ROS that can compromise lysosomal integrity, induce lipid peroxidation, reduce phagocytic capacity, and cause RPE cell death (Citation92,Citation93). It has been postulated that choroidal dendritic cells are activated and recruited by injured RPE and oxidized proteins and lipids in Bruch's membrane. Inflammatory cells and leucocytes are involved in the later stages of AMD (Citation94,Citation95). Activated macrophages and other inflammatory cells secrete enzymes that can damage cells and degrade Bruch's membrane. By releasing cytokines, inflammatory cells might promote the growth of CNV into the sub-RPE space (Citation96). Wet AMD is characterized by the proliferation of CNV. CNV originates from the choroid and extends through a defect in Bruch's membrane and into a plane between RPE and Bruch's membrane, or through the RPE into the subretinal space. CNV arise as capillary-like structures with multiple points of origin (Citation97). CNV can cause serous detachment of the RPE or retina, RPE tears, haemorrhages, and lipid exudation. Morphometric data indicate that the elastic lamina of Bruch's membrane in the macula is 3–6-fold thinner and 2–5-fold less dense relative to that in the mid-periphery in individuals of all ages, rendering the macula more susceptible to the ingrowth of CNV than peripheral retina (Citation98). High concentrations of VEGF and VEGF receptors are found in CNV, surrounding tissue, and RPE cells (Citation20,Citation99). Levels of VEGF are increased in cadaver AMD eyes, in the vitreous, and in the plasma of patients with AMD () (Citation100,Citation101).
Figure 6. Consequences of wet AMD.
Current anti-VEGF therapy for ocular neovascular diseases
VEGF is an attractive target in ophthalmological diseases because of its important role in ocular angiogenesis. Anti-VEGF treatments of neovascular AMD and other ocular neovascular diseases have beneficial effects on disease progression. However, intravitreal administration of current treatment molecules has also several risks including endophthalmitis, retinal detachment, cataract, and uveitis. In patients treated for up to 2 years with pegaptanib, the drug was well tolerated in the eye. Endophthalmitis occurred at a rate of 0.16% per injection, retinal detachment at 0.08% per injection, and traumatic cataract 0.07% per injection during the first year of study. Aseptic technique, including periocular and ocular surface preparation, lid speculum, and drape were found to be essential (Citation102). With improved aseptic techniques, the incidence of endophthalmitis was decreased to 0.10% per injection within the second year of the study. Intravenous bevacizumab in cancer therapy has shown systemic side-effects including hypertension, increased rate of thromboembolic events, gastrointestinal perforations, myocardial infarctions, and death (Citation103). Intravitreal injections lead to detectable but significantly lower serum levels of ranibizumab and pegaptanib than the intravenous administration (Citation104). There was no evidence of an increase in deaths, hypertension, or thromboembolic events in the 2-year safety data from the VISION trial. In the first year of the ANCHOR and MARINA trials the risk of myocardial infarction and stroke was slightly higher in the 0.5 mg ranibizumab treated patients compared to the control group. Current anti-VEGF drugs have a relatively short half-life leading to repeated administrations, and the long-term effects of these multiple injections on the structure and function of the eye are still largely unknown (Citation105). Trials for current and new anti-VEGF therapies are summarized in and .
Table II. Clinical trials for anti-VEGF therapy.
Table III. Clinical trials for new anti-VEGF agents.
Pegaptanib
Pegaptanib, a pegylated ribonucleic acid oligonucleotide aptamer, was the first antiangiogenic aptamer approved by the Food and Drug Administration (FDA) and the European Medicines Agency (EMA) for the treatment of neovascular AMD. Pegaptanib binds specifically to VEGF-A165 and inhibits angiogenesis and pathologic leakage. The recommended dose of pegaptanib is 0.3 mg, and the drug is administered intravitreally every 6 weeks as the molecule is rapidly degraded enzymatically by intraocular nucleases. In the VISION trial 59% of eyes treated with a dose of 0.3 mg pegaptanib lost less than three lines in visual acuity compared to 45% of standard care treated eyes after 2 years (Citation102). A total of 27% of patients discontinuing pegaptanib treatment after the first year experienced an additional mean loss of three lines in visual acuity, suggesting that pegaptanib needs to be injected continuously at least more than 2 years.
Pegaptanib has also been studied in diabetic retinopathy. Safety and efficacy of pegaptanib were assessed with patients with diabetic macular oedema. After 36 weeks, 0.3 mg pegaptanib was significantly superior to sham injections; 34% of patients gained two lines, and also mean central retinal thickness decreased significantly. In addition, a retrospective subgroup analysis revealed that pegaptanib treatment led to the regression of base-line retinal neovascularization in 61% of patients with proliferative diabetic retinopathy (Citation106).
Ranibizumab
Ranibizumab is a recombinant humanized Fab fragment derived from bevacizumab. It was approved by the FDA and the EMA for the treatment of all lesion types in neovascular AMD at a dose of 0.5 mg, and it was the first drug associated with a great proportion of patients with an improvement in visual acuity. It has a rather small molecular weight of 48 kDa, enabling the drug to penetrate the inner limiting membrane and reach the subretinal space when injected intravitreally. It binds all biologically active isotypes of VEGF with high affinity. The half-life of ranibizumab is 2–4 days, resulting in a rapid systemic clearance and good systemic safety. In the MARINA study, 90% of eyes treated with monthly injections of 0.5 mg ranibizumab demonstrated stable vision after 24 months (Citation107). Similar results were announced with the ANCHOR study where monthly injections of 0.3 or 0.5 mg ranibizumab were compared to photodynamic therapy (PDT). At 1 year, 96% of all eyes treated with 0.5 mg ranibizumab demonstrated stable vision (Citation108). The FOCUS trial evaluated the safety, tolerability, and efficacy of ranibizumab in combination with PDT compared to PDT alone. At 1 year, 90% of patients treated with ranibizumab in combination with PDT had stable visual acuity (Citation109). The HORIZON study assessed long-term efficacy of intravitreal 0.5 mg ranibizumab injections administered on an as-needed basis to patients with all types of wet AMD who had completed the MARINA, ANCHOR, or FOCUS trials. HORIZON evaluated three cohorts: ‘Treated initial’ were patients who had been randomized to therapy in one of the earlier studies and had received monthly injections for 2 years; ‘treated cross-over’ were patients who had been randomized to control groups in the earlier trials but were treated with ranibizumab at some point in the initial or HORIZON trial; and ‘untreated’ were those who never received ranibizumab during the full 3 years of study participation. At the end of 1 year, approximately 60% of patients in the treated initial group and 85% of the treated cross-over group had received three to four additional injections of ranibizumab. The majority of patients in all cohorts remained unchanged. Instead, the visual acuity gain seen in the initial trials decreased with less frequent ranibizumab dosing. It may be the result of less regular administration of ranibizumab, or it may be due to the fact that some vision loss is inevitable as the disease progresses, even in the face of treatment. No new safety issues emerged (Citation110). Results from several different studies emphasize the importance of an individual treatment schedule in the treatment of neovascular AMD. The PIER study evaluated the efficacy of ranibizumab administered monthly for three intravitreal injections followed by fixed re-treatments every 3 months. Overall, patients remained stable at base-line visual acuity for 12 months similarly to MARINA and ANCHOR. However, the proportion of gainers of more than three lines was significantly lower than in MARINA or in ANCHOR (Citation111). In the Pronto study, 37 patients with neovascular AMD received three monthly intravitreal injections of 0.5 mg ranibizumab and were re-treated with ranibizumab at each monthly visit if any criterion was fulfilled. In this study, 43% of patients gained more than three lines, and the average number of injections was 9.9 at 24 months (Citation112). The study by Adepegba et al. assessed 78 patients with all subtypes of neovascular AMD treated with intravitreal injections of 0.5 mg ranibizumab with two different protocols. Patients in the first group received a loading dose of three monthly injections and after that were treated as needed (3 + PRN). The other group of patients received injections as needed (PRN). At 12 months, 89%–93% of patients had stable visual acuity in both groups, but a significantly higher number of patients gained more than three lines in the 3 + PRN group compared to PRN group (Citation113).
In the READ study, diabetic patients with macular oedema were treated with 0.5 mg of ranibizumab, focal laser photocoagulation, or a combination of 0.5 mg of ranibizumab and focal laser. At 6 months, improvement of three lines or more occurred in 22% in patients treated with 0.5 mg of ranibizumab, and patients had a significantly better visual outcome than focal laser-treated patients (Citation114). In addition, ranibizumab has recently been approved by the FDA for the treatment of macular oedema following retinal vein occlusion. The safety and efficacy of ranibizumab were analysed in patients with macular oedema following branch or central retinal vein occlusion through the BRAVO and the CRUISE studies. In both of these studies, no new safety events were observed, and improvement in visual acuity was seen in 48%–61% of patients (Citation115).
Bevacizumab
Bevacizumab is a full-length recombinant, humanized antibody of a molecular weight of 149 kDa which binds to all VEGF isoforms. Like ranibizumab, the drug reduces angiogenesis and vascular permeability. The drug was originally approved by the FDA for the treatment of metastatic colorectal cancer. Due to its substantially larger molecular weight, local and systemic clearance of bevacizumab may be delayed, resulting in an extended durability of the treatment but associated with higher systemic toxicity (Citation116). Promising results have been reported from a case series including AMD patients treated with intravitreal injections of bevacizumab. Within 3 months, improvement in function and visual acuity was seen in 38% of treated patients, and the mean central retinal thickness decreased significantly (Citation117). Intravitreal bevacizumab has also been given to diabetic patients in monthly intervals, and functional benefit was seen, with mean visual acuity improvement after 2 months (Citation118,Citation119). Also other neovascular diseases have been treated with bevacizumab. Several studies have shown rapid improvement in visual acuity in patients with retinal vein occlusion after intravitreal bevacizumab injection (Citation120). In a case series of patients with retinal vein occlusion, a single bevacizumab treatment increased visual acuity compared to base-line. Afterwards, visual acuity decreased because of an increase in central macular oedema (Citation121). In addition ROP has been successfully treated with intravitreal bevacizumab with no systemic adverse effects in premature babies (Citation122). Bevacizumab appears to have a beneficial effect in the off-label treatment of intraocular angiogenesis.
New anti-VEGF therapies in clinical trials
Aflibercept
Aflibercept, VEGF Trap, is a 110 kDa soluble protein, which binds with high affinity to all members of VEGF family except unprocessed VEGF-C and -D. In a mouse model with laser-induced CNV, aflibercept induced the regression of CNV by 85% within 10 days (Citation123). Furthermore, CNV disappeared completely in a large percentage of eyes. The safety, tolerability, and bioactivity of intravitreal aflibercept for the treatment of neovascular AMD were evaluated in the CLEAR-IT-1 study. Aflibercept was well tolerated, and there were no serious adverse events. At 6 weeks visual acuity remained stable or improved in 95% of patients, and the total area of CNV was decreased by 35%. In the two highest dose groups combined, 50% of patients demonstrated improvement of three lines in visual acuity (Citation124). The VIEW trial will evaluate the safety and efficacy of two different dosing protocols of intravitreal aflibercept with different doses compared with monthly injections of 0.5 mg ranibizumab in patients with wet AMD.
The DA VINCI trial included 219 patients with clinically significant diabetic macular oedema receiving two different doses of aflibercept with different treatment protocols or macular laser therapy at week 1. Two groups received monthly doses of 0.5 or 2.0 mg of aflibercept, and two groups received three initial monthly doses of 2.0 mg of aflibercept at base-line and weeks 4 and 8, followed by either every 8 week injection of aflibercept or as-needed (PRN) dosing with specific repeat dosing criteria. After 24 weeks, all treatment protocols were superior compared to the control treatment. The best results were accomplished with the loading dose of three monthly injections of aflibercept 2 mg followed by injection as needed. In this group, patients gained an average of 10.3 letters after 4.4 injections (Citation125).
RNA interference
The production of growth factors can be inhibited by RNA interference. Small interfering RNA (siRNA) is a double-stranded RNA, which consists of 21–22 nucleotides. After being further processed in an RNA-induced silencing complex (RISC) by intracellular enzymes, the fragment binds specifically to messenger RNA (mRNA), causing cleavage and further degradation of the mRNA and stop in translation at ribosomal level. The RISC can then bind to other mRNA molecules, and the process is repeated multiple times resulting in a very efficient overall inhibition of the production of the targeted protein (Citation116). Bevasiranib, siRNA targeting VEGF, has been shown to inhibit retinal neovascularization in a mouse model (Citation126). In non-human primates the area of laser-induced CNV was significantly decreased with intravitreal injection of bevasiranib (Citation127). A phase II CARE (Cand5 Anti-VEGF RNAi Evaluation) study assessed the safety and efficacy of bevasiranib with patients with wet AMD. No local or systemic serious adverse events were found. However, there was evidence of continuing deterioration during the first 3 weeks of treatment. It was considered that by targeting a relatively upstream component of the VEGF pathway, bevasiranib has a delayed effect in influencing disease processes. Intravitreous or periocular injection of AGN211745 (Sirna-027), siRNA directed against VEGFR-1, resulted in significant reductions in the area of neovascularization in mouse models of retinal and choroidal neovascularization (Citation128). A phase I dose escalation study with AGN211745 in patients with AMD showed single intravitreal injection of siRNA-027 to be safe and well tolerated. Visual acuity was stabilized in 92% of patients at 3 months, and decreased foveal thickness was seen in some patients. However, the mechanism of action with siRNAs is not completely clear. A recent study by Kleinman et al. shows that untargeted 21-nucleotide or longer siRNAs are as effective as VEGF-A-targeted siRNA at suppressing CNV. According to this study, the antiangiogenesis is not due to target knock-down but due to activation of toll-like receptor 3 and to immunity pathways (Citation129).
Future prospects
Although the role of VEGF in retinal and choroidal angiogenesis is broadly acknowledged, there are controversial data about VEGF and VEGFR-2 polymorphisms exerting a significant influence on risk of neovascular AMD, diabetic retinopathy, or macular oedema (Citation130,Citation131). In addition, the current anti-VEGF treatments for the ocular neovascular diseases in the retina and the choroid have an effect only on VEGF, and the other angiogenic factors involved in the ocular neovascularization are not affected. Potential future approaches to pathological angiogenesis could be based on angiogenesis inhibitors that simultaneously target several angiogenic factors or on blocking some master modulators such as hypoxia-inducible factor (HIF)-1α. HIF-1α is activated in hypoxic conditions preceding the development of various retinopathies and stimulates the expression of proangiogenic growth factors (Citation132). Manipulation of the HIF-1α pathway might be an attractive choice in addition to targeting VEGF and other growth factors. Furthermore, inhibition of some other angiogenic factors like Tie-2 or PDGFs together with VEGF might have more efficient suppression of retinal angiogenesis than inhibition of VEGF alone (Citation133,Citation134). ANG-2 and its signalling pathways provide new molecular targets for the development of new treatments for ocular neovascularization. Also, inhibition of integrins as well as inflammation involved in many ocular neovascular diseases offer alternative treatment pathways (Citation135,Citation136).
However, it must be remembered that angiogenic growth factors also have important roles and protective properties in the normal eye. VEGF is involved in normal vascular development of the retina, and VEGF secreted by the RPE has a role in the maintenance of the choriocapillaris. It has been shown that retinal tissue development occurred normally in an infant with two intravitreal injections of bevacizumab (Citation122). However, it is possible that the developing organs of an ROP infant are at risk to alterations and may be more vulnerable to VEGF blockade than those of an adult. Furthermore, the blood–retinal barrier may be incomplete or compromised in premature babies (Citation137). In a study by Saint-Geniez et al., systemic neutralization of VEGF in mice by adenoviral expression of sFlt1 resulted in significant increase in neural cell death which associated with reduced thickness of the inner and outer nuclear layers and a decline in retinal function as measured by electroretinogram (Citation138). Similarly, transgenic VEGF188/188 mice lacking the other isoforms of VEGF showed a progressive degeneration characterized by choriocapillaris atrophy and RPE loss in aged animals. Increased photoreceptor apoptosis in aged mice led to a decline in visual acuity as detected by electroretinogram (Citation139). Blocking the action of VEGF completely can be harmful in the long run, and anti-VEGF therapies should be administered with caution both in premature babies and elderly patients.
Most of the existing angiogenic inhibitors are large proteins or peptides, with a relatively short half-life and a need for repeated intravitreal injections. With systemic administration the drug is not able efficiently to reach the retina and choroid because of the blood–retina barrier. In addition, phenotypes of retinal diseases vary from early and severe to late and progressive. Slowly progressive diseases have a much wider therapeutic window than more acute, aggressively progressing and those with an earlier onset. In chronic diseases like AMD and diabetic retinopathy permanent expression of therapeutic agent might be required. Also, the therapy should occur early enough before the retina is permanently damaged. Gene therapy, a strategy where genes are delivered into the target cells to treat a disease, could be an option to repeated administrations that are costly and increase the risks of complications. Safer and more sustained regimens need to be developed for intraocular antiangiogenic therapy.
Declaration of interest: This study was supported by the Finnish Academy, Finnish Cultural Foundation, Emil Aaltonen Foundation, Finnish Diabetes Research Foundation and Finnish Cardiovascular Research Foundation. The authors declare no conflicts of interest.
References
- Alm A. Ocular circulation. Hart WM. Adler's Physiology of the Eye. St Louis, USA: Mosby Year Book; 1992. 198–227.
- Folkman J, Ingber D. Inhibition of angiogenesis. Semin Cancer Biol. 1992;3:89–96.
- WHO. Prevent from blindness, WHO report 9–11 November 2005.
- Smith W, Assink J, Klein R, Mitchell P, Klaver CCW, Klein BEK, . Risk factors for age-related macular degeneration: Pooled findings from three continents. Ophthalmology. 2001;108:697–704.
- Senger D, Galli S, Dvorak A, Perruzzi C, Harvey V, Dvorak H. Tumor cells secrete a vascular permeability factor that promotes accumulation of ascites fluid. Science. 1983;219: 983–5.
- Olofsson B, Pajusola K, Kaipainen A, von Euler G, Joukov V, Saksela O, . Vascular endothelial growth factor B, a novel growth factor for endothelial cells. Proc Natl Acad Sci U S A. 1996;93:2576–81.
- Maglione D, Guerriero V, Viglietto G, Delli-Bovi P, Persico M. Isolation of a human placenta cDNA coding for a protein related to the vascular permeability factor. Proc Natl Acad Sci U S A. 1991;88:9267–71.
- Joukov V, Kaipainen A, Jeltsch M, Pajusola K, Olofsson B, Kumar V, . Vascular endothelial growth factors VEGF-B and VEGF-C. J Cell Physiol. 1997;173:211–5.
- Achen M, Jeltsch M, Kukk E, Makinen T, Vitali A, Wilks A, . Vascular endothelial growth factor D (VEGF-D) is a ligand for the tyrosine kinases VEGF receptor 2 (Flk1) and VEGF receptor 3 (Flt4). Proc Natl Acad Sci U S A. 1998; 95:548–53.
- Yamazaki Y, Takani K, Atoda H, Morita T. Snake venom vascular endothelial growth factors (VEGFs) exhibit potent activity through their specific recognition of KDR (VEGF Receptor 2). J Biol Chem. 2003;278:51985–8.
- Ogawa S, Oku A, Sawano A, Yamaguchi S, Yazaki Y, Shibuya M. A novel type of vascular endothelial growth factor, VEGF-E (NZ-7 VEGF), preferentially utilizes KDR/Flk-1 receptor and carries a potent mitotic activity without heparin-binding domain. J Biol Chem. 1998;273: 31273–82.
- Ferrara N, Houck KA, Jakeman LB, Winer J, Leung DW. The vascular endothelial growth factor family of polypeptides. J Cell Biochem. 1991;47:211–8.
- Senger DR, Connolly DT, Van De Water L, Feder J, Dvorak HF. Purification and NH2-terminal amino acid sequence of guinea pig tumor-secreted vascular permeability factor. Cancer Res. 1990;50:1774–8.
- Carmeliet P, Ferreira V, Breier G, Pollefeyt S, Kieckens L, Gertsenstein M, . Abnormal blood vessel development and lethality in embryos lacking a single VEGF allele. Nature. 1996;380:435–9.
- Pe'er J, Shweiki D, Itin A, Hemo I, Gnessin H, Keshet E. Hypoxia-induced expression of vascular endothelial growth factor by retinal cells is a common factor in neovascularizing ocular diseases. Lab Invest. 1995;72:638–45.
- Dorey CK, Aouididi S, Reynaud X, Dvorak HF, Brown LF. Correlation of vascular permeability factor/vascular endothelial growth factor with extraretinal neovascularization in the rat. Arch Ophthalmol. 1996;114:1210–7.
- Aiello LP, Avery RL, Arrigg PG, Keyt BA, Jampel HD, Shah ST, . Vascular endothelial growth factor in ocular fluid of patients with diabetic retinopathy and other retinal disorders. N Engl J Med. 1994;331:1480–7.
- Funatsu H, Yamashita H, Noma H, Mimura T, Yamashita T, Hori S. Increased levels of vascular endothelial growth factor and interleukin-6 in the aqueous humor of diabetics with macular edema. Am J Ophthalmol. 2002;133: 70–7.
- Young TL, Anthony DC, Pierce E, Foley E, Smith LE. Histopathology and vascular endothelial growth factor in untreated and diode laser-treated retinopathy of prematurity. J AAPOS. 1997;1:105–10.
- Kvanta A, Algvere P, Berglin L, Seregard S. Subfoveal fibrovascular membranes in age-related macular degeneration express vascular endothelial growth factor. Invest Ophthalmol Vis Sci. 1996;37:1929–34.
- Kim I, Ryan A, Rohan R, Amano S, Agular S, Miller J, . Constitutive expression of VEGF, VEGFR-1, and VEGFR-2 in normal eyes. Invest Ophthalmol Vis Sci. 1999;40:2115–21.
- Ferrara N, Gerber H, LeCouter J. The biology of VEGF and its receptors. Nat Med. 2003;9:669–76.
- Akiyama H, Mohamedali KA, E Silva RL, Kachi S, Shen J, Hatara C, . Vascular targeting of ocular neovascularization with a vascular endothelial growth factor121/gelonin chimeric protein. Mol Pharmacol. 2005;68:1543–50.
- Saint-Geniez M, Maldonado AE, D'Amore PA. VEGF expression and receptor activation in the choroid during development and in the adult. Invest Ophthalmol Vis Sci. 2006;47:3135–42.
- Shima DT, Nishijima K, Jo N, Adamis AP. VEGF-mediated neuroprotection in ischemic retina. Invest Ophthalmol Vis Sci. 2004;45:3270.
- Alon T, Hemo I, Itin A, Pe'er J, Stone J, Keshet E. Vascular endothelial growth factor acts as a survival factor for newly formed retinal vessels and has implications for retinopathy of prematurity. Nat Med. 1995;1:1024–8.
- Yi X, Mai L, Uyama M, Yew DTW. Time-course expression of vascular endothelial growth factor as related to the development of the retinochoroidal vasculature in rats. Exp Brain Res. 1998;118:155–60.
- Peters S, Heiduschka P, Julien S, Ziemssen F, Fietz H, Bartz-Schmidt KU, . Ultrastructural findings in the primate eye after intravitreal injection of bevacizumab. Am J Ophthalmol. 2007;143:995–1002.
- Reichelt M, Shi S, Hayes M, Kay G, Batch J, Gole GA, . Vascular endothelial growth factor-B and retinal vascular development in the mouse. Clin Experiment Ophthalmol. 2003;31:61–5.
- Joukov V, Sorsa T, Kumar V, Jeltsch M, Claesson-Welsh L, Cao Y, . Proteolytic processing regulates receptor specificity and activity of VEGF-C. EMBO J. 1997;16:3898–911.
- Cao Y, Linden P, Farnebo J, Cao R, Eriksson A, Kumar V, . Vascular endothelial growth factor C induces angiogenesis in vivo. Proc Natl Acad Sci U S A. 1998;95:14389–94.
- Jeltsch M, Kaipainen A, Joukov V, Meng X, Lakso M, Rauvala H, . Hyperplasia of lymphatic vessels in VEGF-C transgenic mice. Science. 1997;276:1423–5.
- Ikeda Y, Yonemitsu Y, Onimaru M, Nakano T, Miyazaki M, Kohno R, . The regulation of vascular endothelial growth factors (VEGF-A, -C, and -D) expression in the retinal pigment epithelium. Exp Eye Res. 2006;83:1031–40.
- Kinnunen K, Puustjarvi T, Terasvirta M, Nurmenniemi P, Heikura T, Laidinen S, . Differences in retinal neovascular tissue and vitreous humour in patients with type 1 and type 2 diabetes. Br J Ophthalmol. 2009;93:1109–15.
- Ohno-Matsui K, Yoshida T, Uetama T, Mochizuki M, Morita I. Vascular endothelial growth factor upregulates pigment epithelium-derived factor expression via VEGFR-1 in human retinal pigment epithelial cells. Biochem Biophys Res Commun. 2003;303:962–7.
- Khaliq A, Foreman D, Ahmed A, Weich H, Gregor Z, McLeod D, . Increased expression of placenta growth factor in proliferative diabetic retinopathy. Lab Invest. 1998;78: 109–16.
- Terman BI, Dougher-Vermazen M, Carrion ME, Dimitrov D, Armellino DC, Gospodarowicz D, . Identification of the KDR tyrosine kinase as a receptor for vascular endothelial cell growth factor. Biochem Biophys Res Commun. 1992;187: 1579–86.
- Soker S, Takashima S, Miao HQ, Neufeld G, Klagsbrun M. Neuropilin-1 is expressed by endothelial and tumor cells as an isoform-specific receptor for vascular endothelial growth factor. Cell. 1998;92:735–45.
- Fong GH, Rossant J, Gertsenstein M, Breitman ML. Role of the Flt-1 receptor tyrosine kinase in regulating the assembly of vascular endothelium. Nature. 1995;376:66–70.
- Shalaby F, Rossant J, Yamaguchi TP, Gertsenstein M, Wu XF, Breitman ML, . Failure of blood-island formation and vasculogenesis in Flk-1-deficient mice. Nature. 1995; 376:62–6.
- Blaauwgeers HGT, Holtkamp GM, Rutten H, Witmer AN, Koolwijk P, Partanen TA, . Polarized vascular endothelial growth factor secretion by human retinal pigment epithelium and localization of vascular endothelial growth factor receptors on the inner choriocapillaris: evidence for a trophic paracrine relation. Am J Pathol. 1999;155: 421–8.
- Gille H, Kowalski J, Li B, LeCouter J, Moffat B, Zioncheck TF, . Analysis of biological effects and signaling properties of Flt-1 (VEGFR-1) and KDR (VEGFR-2). A reassessment using novel receptor-specific vascular endothelial growth factor mutants. J Biol Chem. 2001;276:3222–30.
- Lahteenvuo JE, Lahteenvuo MT, Kivela A, Rosenlew C, Falkevall A, Klar J, . Vascular endothelial growth factor-b induces myocardium-specific angiogenesis and arteriogenesis via vascular endothelial growth factor receptor-1- and neuropilin receptor-1-dependent mechanisms. Circulation. 2009;119:845–56.
- Herzog Y, Kalcheim C, Kahane N, Reshef R, Neufeld G. Differential expression of neuropilin-1 and neuropilin-2 in arteries and veins. Mech Dev. 2001;109:115–9.
- Shen J, Samul R, Zimmer J, Liu H, Liang X, Hackett S, . Deficiency of neuropilin 2 suppresses VEGF-induced retinal neovascularization. Mol Med. 2004;10:12–18.
- Oh H, Takagi H, Otani A, Koyama S, Kemmochi S, Uemura A, . Selective induction of neuropilin-1 by vascular endothelial growth factor (VEGF): A mechanism contributing to VEGF-induced angiogenesis. Proc Natl Acad Sci U S A. 2002;99:383–8.
- Ishida S, Shinoda K, Kawashima S, Oguchi Y, Okada Y, Ikeda E. Coexpression of VEGF receptors VEGF-R2 and neuropilin-1 in proliferative diabetic retinopathy. Invest Ophthalmol Vis Sci. 2000;41:1649–56.
- He Y, Smith SK, Day KA, Clark DE, Licence DR, Charnock-Jones DS. Alternative splicing of vascular endothelial growth factor (VEGF)-R1 (FLT-1) pre-mRNA is important for the regulation of VEGF activity. Mol Endocrinol. 1999;13:537–45.
- Kendall RL, Wang G, Thomas KA. Identification of a natural soluble form of the vascular endothelial growth factor receptor, FLT-1, and its heterodimerization with KDR. Biochem Biophys Res Commun. 1996;226:324–8.
- Honda M, Sakamoto T, Ishibashi T, Inomata H, Ueno H. Experimental subretinal neovascularization is inhibited by adenovirus-mediated soluble VEGF/flt-1 receptor gene transfection: a role of VEGF and possible treatment for SRN in age-related macular degeneration. Gene Ther. 2000;7:978–85.
- Daley ML, Watzke RC, Riddle MC. Early loss of blue-sensitive color vision in patients with type I diabetes. Diabetes Care. 1987;10:777–81.
- Speiser P, Gittelsohn AM, Patz A. Studies on diabetic retinopathy. 3. Influence of diabetes on intramural pericytes. Arch Ophthalmol. 1968;80:332–7.
- Miyamoto K, Ogura Y. Pathogenetic potential of leukocytes in diabetic retinopathy. Semin Ophthalmol. 1999;14:233–9.
- Hammes H, Lin J, Renner O, Shani M, Lundqvist A, Betsholtz C, . Pericytes and the pathogenesis of diabetic retinopathy. Diabetes. 2002;51:3107–12.
- Li W, Liu X, Yanoff M, Cohen S, Ye X. Cultured retinal capillary pericytes die by apoptosis after an abrupt fluctuation from high to low glucose levels: a comparative study with retinal capillary endothelial cells. Diabetologia. 1996; 39:537–47.
- Zheng L, Szabo C, Kern TS. Poly(ADP-ribose) polymerase is involved in the development of diabetic retinopathy via regulation of nuclear factor-kappaB. Diabetes. 2004;53: 2960–7.
- Kowluru RA, Koppolu P, Chakrabarti S, Chen S. Diabetes-induced activation of nuclear transcriptional factor in the retina, and its inhibition by antioxidants. Free Radic Res. 2003;37:1169–80.
- Gabbay KH. Hyperglycemia, polyol metabolism, and complications of diabetes mellitus. Annu Rev Med. 1975;26: 521–36.
- Brownlee M, Vlassara H, Cerami A. Nonenzymatic glycosylation and the pathogenesis of diabetic complications. Ann Intern Med. 1984;101:527–37.
- Giugliano D, Ceriello A, Paolisso G. Oxidative stress and diabetic vascular complications. Diabetes Care. 1996;19: 257–67.
- Taher MM, Garcia JG, Natarajan V. Hydroperoxide-induced diacylglycerol formation and protein kinase C activation in vascular endothelial cells. Arch Biochem Biophys. 1993;303:260–6.
- Inoguchi T, Battan R, Handler E, Sportsman J, Heath W, King G. Preferential elevation of protein kinase c isoform {beta}II and diacylglycerol levels in the aorta and heart of diabetic rats: differential reversibility to glycemic control by islet cell transplantation. Proc Natl Acad Sci U S A. 1992;89: 11059–63.
- Aiello LP, Clermont A, Arora V, Davis MD, Sheetz MJ, Bursell S. Inhibition of PKC {beta} by oral administration of ruboxistaurin is well tolerated and ameliorates diabetes-induced retinal hemodynamic abnormalities in patients. Invest Ophthalmol Vis Sci. 2006;47:86–92.
- Paques M, Massin P, Gaudric A. Growth factors and diabetic retinopathy. Diabetes Metab. 1997;23:125–30.
- Vinores SA, Youssri AI, Luna JD, Chen YS, Bhargave S, Vinores MA, . Upregulation of vascular endothelial growth factor in ischemic and non-ischemic human and experimental retinal disease. Histol Histopathol. 1997;12:99–109.
- Argon laser scatter photocoagulation for prevention of neovascularization and vitreous hemorrhage in branch vein occlusion. A randomized clinical trial. Branch Vein Occlusion Study Group. Arch Ophthalmol. 1986;104:34–41.
- Pierce E, Avery R, Foley E, Aiello L, Smith L. Vascular endothelial growth factor/vascular permeability factor expression in a mouse model of retinal neovascularization. Proc Natl Acad Sci U S A. 1995;92:905–9.
- Pe'er J, Folberg R, Itin A, Gnessin H, Hemo I, Keshet E. Vascular endothelial growth factor upregulation in human central retinal vein occlusion. Ophthalmology. 1998;105: 412–6.
- Noma H, Funatsu H, Yamasaki M, Tsukamoto H, Mimura T, Sone T, . Pathogenesis of macular edema with branch retinal vein occlusion and intraocular levels of vascular endothelial growth factor and interleukin-6. Am J Ophthalmol. 2005; 140:256–61.
- Terry TL. Fibroblastic overgrowth of persistent tunica vasculosa lentis in infants born prematurely: II. Report of cases—clinical aspects. Trans Am Ophthalmol Soc. 1942; 40:262–84.
- Seiberth V, Linderkamp O. Risk factors in retinopathy of prematurity. A multivariate statistical analysis. Ophthalmologica. 2000;214:131–5.
- An international classification of retinopathy of prematurity. II. The classification of retinal detachment. The International Committee for the Classification of the Late Stages of Retinopathy of Prematurity. Arch Ophthalmol. 1987;105:906–12.
- Ashton N, Ward B, Serpell G. Effect of oxygen on developing retinal vessels with particular reference to the problem of retrolental fibroplasia. Br J Ophthalmol. 1954;38:397–432.
- Chan-Ling T, Tout S, Hollander H, Stone J. Vascular changes and their mechanisms in the feline model of retinopathy of prematurity. Invest Ophthalmol Vis Sci. 1992;33: 2128–47.
- Smith L, Wesolowski E, McLellan A, Kostyk S, D'Amato R, Sullivan R, . Oxygen-induced retinopathy in the mouse. Invest Ophthalmol Vis Sci. 1994;35:101–11.
- Ambati J, Ambati BK, Yoo SH, Ianchulev S, Adamis AP. Age-related macular degeneration: etiology, pathogenesis, and therapeutic strategies. Surv Ophthalmol. 2003;48:257–93.
- Dunaief JL, Dentchev T, Ying G, Milam AH. The role of apoptosis in age-related macular degeneration. Arch Ophthalmol. 2002;120:1435–42.
- Ferris FL 3rd, Fine SL, Hyman L. Age-related macular degeneration and blindness due to neovascular maculopathy. Arch Ophthalmol. 1984;102:1640–2.
- Risk factors for choroidal neovascularization in the second eye of patients with juxtafoveal or subfoveal choroidal neovascularization secondary to age-related macular degeneration. Macular Photocoagulation Study Group. Arch Ophthalmol. 1997;115:741–7.
- Frennesson C, Nilsson UL, Nilsson SE. Colour contrast sensitivity in patients with soft drusen, an early stage of ARM. Doc Ophthalmol. 1995;90:377–86.
- Midena E, Segato T, Blarzino MC, Degli Angeli C. Macular drusen and the sensitivity of the central visual field. Doc Ophthalmol. 1994;88:179–85.
- Anderson DH, Ozaki S, Nealon M, Neitz J, Mullins RF, Hageman GS, . Local cellular sources of apolipoprotein E in the human retina and retinal pigmented epithelium: implications for the process of drusen formation. Am J Ophthalmol. 2001;131:767–81.
- Hageman G, Mullins R, Russell S, Johnson L, Anderson D. Vitronectin is a constituent of ocular drusen and the vitronectin gene is expressed in human retinal pigmented epithelial cells. FASEB J. 1999;13:477–84.
- Hee MR, Baumal CR, Puliafito CA, Duker JS, Reichel E, Wilkins JR, . Optical coherence tomography of age-related macular degeneration and choroidal neovascularization. Ophthalmology. 1996;103:1260–70.
- Risk factors associated with age-related macular degeneration: A case-control study in the age-related eye disease study: Age-Related Eye Disease Study Report Number 3. Age-Related Eye Disease Study Research Group. Ophthalmology. 2000;107:2224–32.
- Curcio CA, Millican CL. Basal linear deposit and large drusen are specific for early age-related maculopathy. Arch Ophthalmol. 1999;117:329–39.
- Bressler NM, Silva JC, Bressler SB, Fine SL, Green WR. Clinicopathologic correlation of drusen and retinal pigment epithelial abnormalities in age-related macular degeneration. Retina. 1994;14:130–42.
- Strauss O. The retinal pigment epithelium in visual function. Physiol Rev. 2005;85:845–81.
- Kennedy CJ, Rakoczy PE, Constable IJ. Lipofuscin of the retinal pigment epithelium: a review. Eye. 1995;9:763–71.
- Sundelin S, Wihlmark U, Nilsson SE, Brunk UT. Lipofuscin accumulation in cultured retinal pigment epithelial cells reduces their phagocytic capacity. Curr Eye Res. 1998;17:851–7.
- Delori FC, Goger DG, Dorey CK. Age-related accumulation and spatial distribution of lipofuscin in RPE of normal subjects. Invest Ophthalmol Vis Sci. 2001;42:1855–66.
- Holz F, Schutt F, Kopitz J, Eldred G, Kruse F, Volcker H, . Inhibition of lysosomal degradative functions in RPE cells by a retinoid component of lipofuscin. Invest Ophthalmol Vis Sci. 1999;40:737–43.
- Boulton M, Dontsov A, Jarvis-Evans J, Ostrovsky M, Svistunenko D. Lipofuscin is a photoinducible free radical generator. J Photochem Photobiol B. 1993;19:201–4.
- Penfold PL, Killingsworth MC, Sarks SH. Senile macular degeneration: the involvement of immunocompetent cells. Graefes Arch Clin Exp Ophthalmol. 1985;223:69–76.
- Killingsworth MC, Sarks JP, Sarks SH. Macrophages related to Bruch's membrane in age-related macular degeneration. Eye. 1990;4:613–21.
- Oh H, Takagi H, Takagi C, Suzuma K, Otani A, Ishida K, . The potential angiogenic role of macrophages in the formation of choroidal neovascular membranes. Invest Ophthalmol Vis Sci. 1999;40:1891–8.
- Green WR, Key SN 3rd. Senile macular degeneration: a histopathologic study. 1977. Retina. 2005;25:180–250.
- Chong NHV, Keonin J, Luthert PJ, Frennesson CI, Weingeist DM, Wolf RL, . Decreased thickness and integrity of the macular elastic layer of Bruch's membrane correspond to the distribution of lesions associated with age-related macular degeneration. Am J Pathol. 2005;166: 241–51.
- Kliffen M, Sharma HS, Mooy CM, Kerkvliet S, de Jong, PT. Increased expression of angiogenic growth factors in age-related maculopathy. Br J Ophthalmol. 1997;81:154–62.
- Wells JA, Murthy R, Chibber R, Nunn A, Molinatti PA, Kohner EM, . Levels of vascular endothelial growth factor are elevated in the vitreous of patients with subretinal neovascularisation. Br J Ophthalmol. 1996;80:363–6.
- Lip P, Blann AD, Hope-Ross M, Gibson JM, Lip GYH. Age-related macular degeneration is associated with increased vascular endothelial growth factor, hemorheology and endothelial dysfunction. Ophthalmology. 2001;108:705–10.
- D'Amico DJ. Pegaptanib sodium for neovascular age- related macular degeneration: two-year safety results of the two prospective, multicenter, controlled clinical trials. Ophthalmology. 2006;113:992–1001.
- Hurwitz H, Fehrenbacher L, Novotny W, Cartwright T, Hainsworth J, Heim W, . Bevacizumab plus irinotecan, fluorouracil, and leucovorin for metastatic colorectal cancer. N Engl J Med. 2004;350:2335–42.
- Gaudreault J, Fei D, Rusit J, Suboc P, Shiu V. Preclinical pharmacokinetics of ranibizumab (rhuFabV2) after a single intravitreal administration. Invest Ophthalmol Vis Sci. 2005; 46:726–33.
- Drolet D, Nelson J, Tucker C, Zack P, Nixon K, Bolin R, . Pharmacokinetics and safety of an anti-vascular endothelial growth factor aptamer (NX1838) following injection into the vitreous humor of rhesus monkeys. Pharm Res. 2000;17:1503–10.
- Adamis AP, Altaweel M, Bressler NM, Cunningham ET Jr, Davis MD, Goldbaum M, . Macugen Diabetic Retinopathy Study Group. Changes in retinal neovascularization after pegaptanib (Macugen) therapy in diabetic individuals. Ophthalmology. 2006;113:23–8.
- Rosenfeld PJ, Brown DM, Heier JS, Boyer DS, Kaiser PK, Chung CY, . Ranibizumab for neovascular age-related macular degeneration. N Engl J Med. 2006;355:1419–31.
- Brown DM, Kaiser PK, Michels M, Soubrane G, Heier JS, Kim RY, . Ranibizumab versus verteporfin for neovascular age-related macular degeneration. N Engl J Med. 2006;355:1432–44.
- Antoszyk AN, Tuomi L, Chung CY, Singh A. Ranibizumab combined with verteporfin photodynamic therapy in neovascular age-related macular degeneration (FOCUS): Year 2 results. Am J Ophthalmol. 2008;145:862–74.
- Singer M, Wong P, Wang P, Scott L. HORIZON Extension Trial of Ranibizumab (LUCENTIS®) for Neovascular Age-Related Macular Degeneration (AMD): Two-year safety and efficacy results. Invest Ophthalmol Vis Sci. 2009;50:3093.
- Regillo CD, Brown DM, Abraham P, Yue H, Ianchulev T, Schneider S, . Randomized, double-masked, sham-controlled trial of ranibizumab for neovascular age-related macular degeneration: PIER study year 1. Am J Ophthalmol. 2008;145:239–48.
- Lalwani GA, Rosenfeld PJ, Fung AE, Dubovy SR, Michels S, Feuer W, . A variable-dosing regimen with intravitreal ranibizumab for neovascular age-related macular degeneration: Year 2 of the PrONTO Study. Am J Ophthalmol. 2009; 148:43–58.
- Adepegba OA, Gupta B, Adewoyin T, Patel S, Sivaprasad S. Comparison of two treatment schedules of intravitreal ranibizumab for neovascular age related macular degeneration. Invest Ophthalmol Vis Sci. 2010;51:71.
- Nguyen QD, Shah SM, Heier JS, Do DV, Lim J, Boyer D, . Primary end point (six months) results of the Ranibizumab for Edema of the mAcula in Diabetes (READ-2) Study. Ophthalmology. 2009;116:2175–81.
- Ho AC, Gray S, Rundle A, Li Z, Yee Murahashi W, Rubio R, . Ranibizumab in patients with macular edema following retinal vein occlusion: 12-month outcomes of BRAVO and CRUISE. Invest Ophthalmol Vis Sci. 2010;51:6452.
- Schmidt-Erfurth UM, Pruente C. Management of neovascular age-related macular degeneration. Prog Retin Eye Res. 2007;26:437–51.
- Spaide RF, Laud K, Fine HF, Klancnik JM Jr, Meyerle CB, Yannuzzi LA, . Intravitreal bevacizumab treatment of choroidal neovascularization secondary to age-related macular degeneration. Retina. 2006;26:383–90.
- Avery RL, Pearlman J, Pieramici DJ, Rabena MD, Castellarin AA, Nasir MA, . Intravitreal bevacizumab (Avastin) in the treatment of proliferative diabetic retinopathy. Ophthalmology. 2006;113:1695–705.
- Spaide RF, Fisher YL. Intravitreal bevacizumab (Avastin) treatment of proliferative diabetic retinopathy complicated by vitreous hemorrhage. Retina. 2006;26:275–8.
- Kreutzer TC, Alge CS, Wolf AH, Kook D, Burger J, Strauss R, . Intravitreal bevacizumab for the treatment of macular oedema secondary to branch retinal vein occlusion. Br J Ophthalmol. 2008;92:351–5.
- Stahl A, Agostini H, Hansen L, Feltgen N. Bevacizumab in retinal vein occlusion-results of a prospective case series. Graefes Arch Clin Exp Ophthalmol. 2007;245:1429–36.
- Kong L, Mintz-Hittner HA, Penland RL, Kretzer FL, Chevez-Barrios P. Intravitreous bevacizumab as anti-vascular endothelial growth factor therapy for retinopathy of prematurity: a morphologic study. Arch Ophthalmol. 2008;126: 1161–3.
- Saishin Y, Saishin Y, Takahashi K, Lima e Silva R, Hylton D, Rudge JS, . VEGF-TRAP(R1R2) suppresses choroidal neovascularization and VEGF-induced breakdown of the blood-retinal barrier. J Cell Physiol. 2003;195:241–8.
- Nguyen QD, Shah SM, Browning DJ, Hudson H, Sonkin P, Hariprasad SM, . A phase I study of intravitreal vascular endothelial growth factor trap-eye in patients with neovascular age-related macular degeneration. Ophthalmology. 2009; 116:2141–8.
- Major JC Jr, Brown DM; DA VINCI Study Group. DA VINCI: DME and VEGF Trap-Eye: INvestigation of Clinical Impact: Phase 2 Study in patients with diabetic macular edema (DME). Invest Ophthalmol Vis Sci. 2010;51:6426.
- Reich SJ, Fosnot J, Kuroki A, Tang W, Yang X, Maguire AM, . Small interfering RNA (siRNA) targeting VEGF effectively inhibits ocular neovascularization in a mouse model. Mol Vis. 2003;9:210–6.
- Tolentino MJ, Brucker AJ, Fosnot J, Ying GS, Wu IH, Malik G, . Intravitreal injection of vascular endothelial growth factor small interfering RNA inhibits growth and leakage in a nonhuman primate, laser-induced model of choroidal neovascularization. Retina. 2004;24:660.
- Shen J, Samul R, Silva RL, Akiyama H, Liu H, Saishin Y, . Suppression of ocular neovascularization with siRNA targeting VEGF receptor 1. Gene Ther. 2006;13:225–34.
- Kleinman ME, Yamada K, Takeda A, Chandrasekaran V, Nozaki M, Baffi JZ, . Sequence- and target-independent angiogenesis suppression by siRNA via TLR3. Nature. 2008;452:591–7.
- Fang AM, Lee AY, Kulkarni M, Osborn MP, Brantley MA Jr. Polymorphisms in the VEGFA and VEGFR-2 genes and neovascular age-related macular degeneration. Mol Vis. 2009; 15:2710–9.
- Abhary S, Burdon KP, Gupta A, Lake S, Selva D, Petrovsky N, . Common sequence variation in the VEGFA gene predicts risk of diabetic retinopathy. Invest Ophthalmol Vis Sci. 2009;50:5552–8.
- Semenza GL, Wang GL. A nuclear factor induced by hypoxia via de novo protein synthesis binds to the human erythropoietin gene enhancer at a site required for transcriptional activation. Mol Cell Biol. 1992;12:5447–54.
- Boyer DS; Ophthotech Anti-PDGF in AMD Study Group. Combined inhibition of platelet derived (PDGF) and vascular endothelial (VEGF) growth factors for the treatment of neovascular age-related macular degeneration (NV-AMD)—results of a phase 1 study. Invest Ophthalmol Vis Sci. 2009; 50:1260.
- Takagi H, Koyama S, Seike H, Oh H, Otani A, Matsumura M, . Potential role of the angiopoietin/Tie2 system in ischemia-induced retinal neovascularization. Invest Ophthalmol Vis Sci. 2003;44:393–402.
- Apte RS; Ophthotech Anti-Complement in AMD Study Group. Targeting complement factor 5 in neovascular age-related macular degeneration (NV-AMD)—Results of a Phase 1 Study. Invest Ophthalmol Vis Sci. 2009;50:5011.
- Zahn G, Vossmeyer D, Stragies R, Wills M, Wong CG, Loffler KU, . Preclinical evaluation of the novel small-molecule integrin {alpha}5{beta}1 inhibitor JSM6427 in monkey and rabbit models of choroidal neovascularization. Arch. Ophthalmol. 2009;127:1329–35.
- Avery RL. Extrapolating anti-vascular endothelial growth factor therapy into pediatric ophthalmology: Promise and concern. J AAPOS. 2009;13:329–31.
- Saint-Geniez M, Maharaj ASR, Walshe TE, Tucker BA, Sekiyama E, Kurihara T, . Endogenous VEGF is required for visual function: evidence for a survival role on müller cells and photoreceptors. PLoS ONE. 2008; 3:e3554.
- Saint-Geniez M, Kurihara T, Sekiyama E, Maldonado AE, D'Amore PA. An essential role for RPE-derived soluble VEGF in the maintenance of the choriocapillaris. Proc Natl Acad Sci U S A. 2009;106:18751–6.

