Abstract
Polyphenols are widely distributed in the plant kingdom and represent an abundant antioxidant component of the human diet. The review offers a brief description of the chemistry and occurrence in plant food of four important groups of polyphenols: phenolic acids, flavonoids, stilbenes, and lignans. Interest in the possible health benefits of polyphenols has increased due to corresponding antioxidant capacity. Considerable evidence is now available showing anticarcinogenic effects of polyphenolic compounds, as well as potential to prevent cardiovascular and cerebrovascular diseases. The use of botanicals has received a large amount of attention in recent years. Epidemiological studies have shown that a reduced risk of cancer is associated with diets rich in vegetables and fruits, and methods for the discovery and characterization of active compounds from plant sources are available. These results are promising, but additional research on the molecular mechanism of action of polyphenols and their application to human health is required.
Introduction
Polyphenols form a complex group of molecules associated with most plant cell walls. They range in chemical complexity from simple phenolic acids (e.g., caffeic acid) to high-molecular-weight tannins. More than 8000 phenolic and polyphenolic compounds have been identified in various plant species, where they have several important functions. Examples include inhibition of pathogen development, decay of microorganisms, and protection against UV radiation, photosynthetic stress, reactive oxygen species, wounds, and herbivores. Some polyphenols are essential for plant physiology, being involved in diverse functions such as pigmentation, pollination, enhanced predator resistance (by acting as phytoalexins), or increased astringency, thus making plants unpalatable as foods. They protect crops from plague and preharvest seed germination. Polyphenols have many industrial applications, such as in the production of paints, paper, cosmetics, as tanning agents, and in the food industry as natural colorants and preservatives. In addition, some phenolic compounds are antibiotics and antidiarrheal, antiulcer, and anti-inflammatory agents. They can be used in the treatment of diseases such as hypertension, vascular fragility, allergies, and hypercholesterolemia (Bravo, Citation1998; Higdon & Frei, Citation2003).
Chemoprevention is a promising new approach in cancer research wherein agents are administered to inhibit, delay, or reverse the process of carcinogenesis. Epidemiological data have suggested that high consumption of vegetables and fruits is associated with a low risk of cancer and cardiovascular diseases (CitationAmerican Institute of Cancer Research/World Cancer Research Fund, 1997; van Duyn & Pivonka, Citation2000; Byers, Citation2002; Bub et al., Citation2003), whereas a high intake of red meat may increase some types of cancer (Hollman et al., Citation1999). A variety of in vitro studies have shown that polyphenols mediate antioxidative and immunomodulatory activities (Middleton, Citation1998; Ross & Kasum, Citation2002; Dragsted, Citation2003). Further, polyphenols have potential to prevent genotoxicity by reducing exposure to oxidative and carcinogenic factors (Pool-Zobel et al., Citation1999). There has been considerable scientific interest in the possibility that increased intake of dietary antioxidants may protect against some chronic diseases. Among the dietary phenolic compounds, flavonoids have extensively been studied, and there are several reviews relating to the antioxidant activity of these compounds (Lairon & Amiot, Citation1999; Chan et al., Citation2003; Steinberg et al., Citation2003; Urso & Clarksonn, Citation2003).
There is growing interest in medicinal botanicals as part of complementary medicine in the United States. Approximately 40% of Americans use alternative remedies, including herbal medicine, for disease prevention and therapy (Eisenberg et al., Citation1998). The high cost, side effects, and therapeutic limitations of conventional medications are key factors that drive the revival of herbal remedies. People can easily obtain herbal medicines or botanical supplements on an over-the-counter basis and feel comfortable consuming these products, even though safety and efficacy are generally not established on a rigorous scientific basis. This may be problematic due to factors such as drug-drug interactions (von Gyuenigen & Hopkins, Citation2000; Go et al., Citation2001). On the other hand, these preparations may mediate beneficial responses, either due to active chemical constituents or placebo effects. In any case, botanicals have been used by human beings throughout history.
Conventional medical literature also documents the widespread use of herbal remedies in many countries throughout the world, and the presence of certain phytochemical components supports the pharmacological and physiological efficacy of some ethnomedical treatment regimens. The cancer inhibitory potential of human nutrients derived from plants, as well as of non-nutrient constituents of phytochemicals, has been confirmed in various animal models (Pezzuto, Citation1997). For more than 30 years, the prevention of cancer has been emphasized as a preferable option relative to chemotherapy. Significant success has been realized through the control of tobacco-related cancers by means of education and prohibition of smoking in various public settings. In addition, cancer chemoprevention has been developed as a major field of scientific investigation. Accordingly, a great deal of evidence has suggested the protective role of a diet rich in fruits and vegetables (van Duyn & Pivonka, Citation2000; Terry et al., Citation2001). Overall, diets high in vegetables and fruits (more than 400 g/day) may prevent at least 20% of all cancers. Some of the most convincing evidence for the health benefits of fruit and vegetable consumption relates to the reduced risk of gastrointestinal cancers, such as those associated with the mouth, pharynx, esophagus, stomach, colon, and rectum. Vegetables are more effective than fruits (Terry et al., Citation2001). The mechanisms by which vegetables and fruits reduce cancer are likely to be multiple and complex. Various stages of carcinogenesis may be inhibited, and various in vitro or in vivo systems may be used to model these inhibitory effects. It is logical to acquire compelling in vitro data prior to performing tests with animal models, and it is generally necessary to isolate and characterize active chemical principles.
The current review is focused on characterization, bioavailability, and cancer chemopreventive potential of several classes of polyphenols. In addition, we describe the process of cancer chemopreventive drug discovery from plant sources with characterization of experimental approaches, with special emphasis on work performed in our laboratory.
Nature and occurrence of phenolic phytochemicals
Phenolic compounds comprise one of the largest groups of plant metabolites, and they are an important part of human diets. All plant phenolic compounds arise from a common intermediate, phenylalanine, or a close precursor, shikimic acid. They arise biogenetically from two main synthetic pathways: the shikimate pathway and the acetate pathway (Herrmann, Citation1995). Plants polyphenols can range from simple species, such as phenolic acids, to highly polymerized compounds, such as tannins. They occur primarily in conjugated forms, with one or more sugar residues linked to hydroxyl groups, although direct linkages of the sugar to an aromatic carbon also exists. Sugars can be monosaccharides, disaccharides, or oligosaccharides. Glucose is the most common sugar residue. Association with other compounds, such as carboxylic and organic acids, amines, and lipids, and linkages with other phenols are also common. They can be divided into at least 10 different classes based on their general chemical structure (Bravo, Citation1998). For this review, we have chosen four groups for discussion, with emphasis on their possible applications for cancer prevention. Flavonoids and phenolic acids most commonly are widespread in foods.
Phenolic acids
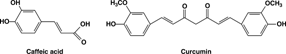
Hydroxybenzoic acids and hydroxycinnamic acids account for about a third of the phenolic compounds in our diet. The most frequently encountered species is caffeic acid (). Caffeic acid is found in many dietary substances, such as apples, plums, tomatoes, and grapes. Derivatives of hydroxycinnamic acid are found in nearly every plant. One of the most widespread representatives is curcumin (). It is widely used as a food preservative and yellow coloring agent for foods, drugs, and cosmetics. Phenolic acids can act as antioxidants by a number of pathways, but perhaps the most significant is by free-radical scavenging in which the phenolic compound can quench the free-radical reaction.
Figure 1 Phenolic acids and derivatives.
Flavonoids
The flavonoids are a complex group of polyphenolic plant metabolites found in the food of human beings (Havsteen, Citation2002). All share a three-ring structure of two aromatic centers, rings A and B, and a central oxygenated heterocycle moiety, ring C (). Biogenetically, the A ring usually arises from a molecule of resorcinol, and the B ring is derived from the shikimate pathway. Flavonoids can further be subdivided into six major subclasses, based on variations in the heterocyclic C-ring, including flavones, flavonols, flavonones, catechins, anthocyanidins, and isoflavones (Ross & Kasum, Citation2002). Flavonoids are widely distributed in beverages of plant origin, such as tea, cocoa, and wine. Tea and onions are the main dietary sources of flavonols and flavones. It has been estimated that, even in industrialized societies, intakes of flavonoids could be as high as 1 g per day, but this is probably an overestimate based on inadequate analytical data (Hertog & Hollman, Citation1996). A substantial body of experimental work has established that flavonoids can suppress carcinogenesis in animal models, and there is considerable interest in the biological effects of these compounds at the cellular level. Flavonoids interact with cellular signal pathways that control the cell cycle, differentiation, and apoptosis. Their antineoplastic effects can involve antioxidant activity, induction of phase II enzyme activity, inhibition of protein kinases, and interaction with type II estrogen binding sites (Gee & Johnson, Citation2001).
Figure 2 Basic flavonoid structure.
Stilbenes
Stilbenes contain two phenyl moieties connected by a two-carbon methylene bridge (). Most stilbenes in plants act as antifungal phytoalexins, compounds that are synthesized only in response to infection or injury. The most extensively studied stilbene is resveratrol.
Figure 3 Representative structures of stilbenes and lignans.
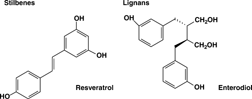
Resveratrol was first recognized as a biologically active compound by Siemann and Creasy (Citation1992). Resveratrol (trans-3,5,4′-trihydroxystilbene) is a phytoalexin, or a class of antibiotics of plant origin, produced primarily in grapes and peanuts as edible plants. The highest concentration of resveratrol (50–100 µg/g) is found in the skin of the grape. The reason for synthesis of resveratrol in grapes is associated with natural stress factors such as gray mold or fungal infections. It is considered to be a part of the plant defense system (Schwekendiek et al., Citation1992). Significant amounts of resveratrol were detected in healthy fruit clusters, prior to visual detection of mold lesions, thus suggesting resveratrol was synthesized soon after recognition of the pathogen by the plant (Jeandet et al., Citation2002). It is conceivable that resveratrol mediates beneficial effects with human cells by mechanisms similar to those mediated in plant cells.
Resveratrol has been shown to have anticancer (chemopreventive), anti-inflammatory, antifungal, and antimicrobial properties. It has a long history of being used as an herbal remedy, especially in oriental medicine for the treatment of lipid, inflammatory, and heart disorders. Most early studies were associated with antioxidant or cardiovascular activities and “stickiness” of blood platelets. Resveratrol has been reported to have a diverse range of pharmacological properties, including anti-inflammatory, estrogen receptor agonist (Gehm et al., Citation1997), and effects on cell signaling pathways, cell proliferation, tumor growth, and apoptosis (CitationStewart et al., 1999). Some experiments suggest that resveratrol inhibits the development of cancer and can be used as a chemopreventive agent, but metabolic pathways and the biological effects of metabolites remain to be characterized (Bhat et al., Citation2001; Bhat & Pezzuto, Citation2002).
Lignans
Lignans are diphenolic compounds that contain a 2,3-dibenzylbutane structure that is formed by the dimerization of two cinnamic acid residues (). Lignans are naturally occurring chemicals that are widespread within plants. Several lignans, such as secoisolariciresinol, are considered to be phytoestrogens. These are especially abundant in flax seed. Bacteria in human intestines convert them into two other lignans, enterolactone and enterodiol, which also have estrogen-like effects. Lignans are being studied for possible use in cancer prevention, particularly against breast cancer. If there is little estrogen in the body, lignans may act like a weak estrogen, but when natural estrogen is abundant in the body, lignans may reduce the effect of estrogen by displacing it from cells. This displacement of the hormone may help prevent some cancers, such as breast cancer, that depend on estrogen. Very early evidence suggests that lignans may also be antioxidants, although the strength of their antioxidant activity is not clear (Kitts et al., Citation1999).
Polyphenols as antioxidants
Plant polyphenols are well recognized for their antioxidant activities. These compounds scavenge free radicals and disrupt the free-radical chain reaction of lipid peroxidation. These antioxidants can be classified as water-soluble or lipid-soluble, depending on whether they act primarily in the aqueous phase or in the lipophilic region of cell membranes. Hydrophilic antioxidants include ascorbic acid and urate. Ubiquinols, retinoids, carotenoids, flavonoids, and tocopherol are representative lipid-soluble antioxidants. Plasma proteins, glutathione, and urate are endogenous, whereas ascorbic acid, carotenoids, retinoids, flavonoids and tocopherols constitute some of the dietary antioxidants. Certain radical scavengers are not recyclable, whereas others are recycled through the intervention of a series of enzyme systems or other nonenzymic antioxidant systems. Dietary exposure to flavonoids is significant. As noted above, the average diet in the U.K. and the U.S. may contain up to 1 g of mixed flavonoids per day. Dietary intake far exceeds that of vitamin E, a monophenolic antioxidant, and of β-carotene (Kandaswami & Middleton, Citation1994).
Flavonoids act as potent metal chelators and free-radical scavengers. They are powerful chain-breaking antioxidants (Santos et al., Citation1998). Moreover, flavonoids are known to possess vitamin C–stabilizing and antioxidant-dependent vitamin C–sparing activities. They are also known to increase the absorption of vitamin C. In addition, flavonoids are known to modify the activities of a host of enzyme systems including protein kinase C, protein tyrosine kinase and various other kinases, aldose reductase, myeloperoxidase, NADPH oxidase, xanthine oxidase, phospholipase, reverse transcriptase, ornithine decarboxylase, lipoxygenase, cyclooxygenase, and so on (Park & Pezzuto, Citation2002a). Some of these enzyme systems are critically involved in immune function, carcinogenesis, cellular transformation, and tumor growth and metastasis. The physiologic and pathologic processes affected by flavonoids are diverse and numerous and include secretion, mitogenesis, platelet aggregation and adhesion to endothelial surface, cell motility, malignant cell proliferation, cancer metastasis, and function/expression of adhesion molecules in various mammalian cell types. The antioxidant function and enzyme-modifying actions of flavonoids could account for many of their pharmacological activities (Santos et al., Citation1998).
Quercetin and other flavonoids are effective inhibitors of O2ċ − production by cells. Quercetin is a potent inhibitor of human neutrophil degranulation and O2ċ − production, and also inhibits the phosphorylation of neutrophil proteins accompanying neutrophil activation by phorbol myristate acetate (Blackburn et al., Citation1987). Quercetin can also suppress lipid peroxidation in several biological systems, such as mitochondria, microsomes, chloroplasts, and erythrocytes. Silymarin, a 3-hydroxyflavone present in Silybum marianum Gaertne (the European milk thistle), protects rat liver mitochondria and microsomes from lipid peroxide formation induced by Fe2+-ascorbate and NADPH-Fe3+-ADP systems (Valenzuela et al., Citation1987). Soybean isoflavonoids have shown antioxidative potency and prevent peroxidative hemolysis of sheep, rat, and rabbit erythrocytes (Santos et al., Citation1998). Quercetin and silybin were reported to exert a protective effect by preventing the decrease in the xanthine dehydrogenase/oxygenase ratio observed during ischemia/reperfusion in the rat (Valenzuela et al., Citation1985). The enzyme xanthine oxidase, implicated in tissue oxidative injury after ischemia/reperfusion, is a source of reactive oxygen species that is formed from a dehydrogenase during ischemia. The protective effect of quercetin and silybin on the xanthine dehydrogenase/oxidase ratio is due to inhibition of the dehydrogenase. The bioactivity of phenolics may be related to their antioxidant behavior, which is attributed to their ability to chelate metals, inhibit lipoxygenase, and scavenge free radicals. However, phenolics can also function as pro-oxidants by chelating metals in a manner that maintains or increases their catalytic activity.
Flavonoids, especially those with catechol or pyrogallol groups, obviously are prone to autoxidation reactions (Bolton et al., Citation1997). The pro-oxidative and antioxidative properties of phenolics from soybeans and other legumes were documented by Morgan et al. (Citation1997). The phenolic acids and flavonoids were able to reduce ferric to ferrous ions and were able to chelate and alter the catalytic activity of iron. Most of the phenolics tested were also able to inhibit the oxidation of linoleic acid micelles and ferrous ion-catalyzed oxidation of glutamine synthase, presumably through free-radical scavenging and removal of iron from catalytic sites via chelation. Although phenolics inhibited oxidation in certain systems, they did not protect against all forms of oxidative damage. Even though the phenolics chelated iron, this metal ion was still catalytically active and able to oxidize both deoxyribose and DNA. Pro-oxidant activity of phenolics has also been observed for carnosol, carnosic acid, quercetin, rutin, and luteolin (Morgan et al., Citation1997).
It was also found that pH was essential in determining the oxidative role of phenolics. In general, a decrease in pH increased iron-reducing activity and reduced the ability of phenolics to chelate and inhibit the catalytic activity of iron. Increasing pH increased deoxyribose and DNA oxidation. Inhibition of lipid oxidation was also influenced by pH, with γ-resorcyclic acid being antioxidative at pH 5.8 and pro-oxidative at pH 7.4. Hydrobenzoic acid was antioxidative, and apigenin-7-glucoside was pro-oxidative at pH 7.4, yet neither had an effect on lipid oxidation at pH 5.8. These results suggest that the pH of biological tissues could also influence the antioxidative/pro-oxidative activity of phenolics (Decker, Citation1997).
A possible mechanism of polyphenol cytotoxicity may be related to their pro-oxidant properties (Sergediene et al., Citation1999). Flavonoids autoxidize in aqueous medium and may form highly reactive HOċ radicals in the presence of transition metals. In addition, polyphenols and flavonoids may act as substrates for peroxidase and other metalloenzymes, yielding quinone- or quinonemethide-type pro-oxidant and/or alkylating products. The pro-oxidant character of polyphenol cytotoxicity is supported by the formation of activated oxygen species during gallic acid–induced apoptosis and by the enhancement of gallic and caffeic acid–induced apoptosis by nontoxic concentrations of copper ions (Yamanaka et al., Citation1997).
Catechins, such as ( − )-epicatechin and ( − )-epigallocatechin, abundant in green tea, possess the antioxidative and pro-oxidative characteristics of Cu2+-induced low-density lipoprotein (LDL) oxidation. In the initiation phase, LDL oxidation was inhibited by addition of catechin. In contrast, during the propagation phase of LDL oxidation, catechins served as accelerators of oxidation. Depending on redox status, they might form reactive oxidation products such as semiquinones and quinones and function to stimulate oxidative reactions (Yamanaka et al., Citation1997).
Quercetin, a highly studied antioxidant flavonoid, has potential to inhibit free-radical processes in cells by (a) scavenging , (b) blocking lipid peroxidation, (c) reacting with peroxyl or lipid peroxyl radicals, (d) inhibiting formation of HOċ, and (e) chelating iron ions. The biological effects of quercetin are believed to result from antioxidant properties. It was demonstrated clearly that quercetin could function both as an antioxidant and a pro-oxidant, depending on concentration and free-radical sources and their location in the cell.
Data on the antioxidant capability of natural polyphenols allows speculation about the antioxidant activities of tea and red wines. Green tea, which is not fermented, is very rich in pyrogallol derivatives. It is customary to assume these components are the dominant reason for the very high antioxidant activity of green tea. Red wines, during the course of aging, exhibit progressive oxidative transformation of polyphenols and a visible decrease in total content. Most likely, this is compensated for by the elevated antioxidant activities of transformation products including dimmers/oligomers. It was recently shown that products of oxidative transformation display a higher ability to inhibit lipid oxidation than original polyphenols (Roginsky, Citation2003).
Anticarcinogenic effects of polyphenols
Carcinogenesis can be viewed as a multistage, microevolutionary process. The progression of tumor formation may be slow, often taking 10 or more years. It is generally agreed that tumors can be derived from single abnormal cells, and work with experimental systems shows that carcinogenesis is divisible into three major stages: initiation, promotion, and progression. Initiation is a heritable aberration of a cell. Cells so initiated can undergo transformation to malignancy if promotion and progression follow. Initiation appears to be irreversible and can result from DNA damage. Promotion, on the other hand, is affected by factors that do not alter DNA sequences and involves the selection and clonal expansion of initiated cells. This process is partly reversible and accounts for a major portion of the lengthy latent period of carcinogenesis. The final stage of tumor formation is the progression of a benign growth to a malignant neoplasm. There is loss of growth control, an escape from the host defense mechanism, and metastasis.
Certain initiators, such as radiation or chemical carcinogens, can induce the production of various free radicals and subsequent DNA base sequence alteration. In addition, cells of the immune system, such as neutrophils and macrophages, produce and H2O2 that have been associated with the induction of experimental cancers. Oxygen free-radicals and methyl radicals are known to damage DNA. In some cases, such free radicals may arise in reactions catalyzed by ferric and cupric ions localized in the vicinity of cellular DNA. Free radical–mediated DNA damage can have serious consequences on an organism unless the damage is repaired. Although oxygen free-radical effects can lead to DNA damage, they may also directly affect the protein components of the DNA repair apparatus. Unrepaired DNA alterations are inherited as mutations. Phase I metabolizing enzymes (e.g., cytochrome P450) play an important role in the initiation stage. Through the catalytic activity of these enzymes, a polar reactive group is added to lipophilic carcinogens/xenobiotics to form an electrophile, which can react with DNA. Later, the xenobiotic can be detoxified by phase II metabolizing enzymes, with sugars, amino acids, and glutathione (Galati et al., Citation2000), for example. Green and black tea extracts strongly inhibit neoplastic transformation in mammary organ cultures or epithelial cells (Steele et al., Citation2000). In animals, green or black tea also induce the phase I metabolizing enzyme CYP1A2.
It is generally believed that the generation of growth promotion oxidants is a major trigger of the tumor promotion and progression stages. Tumor-promoting phorbol esters not only can induce changes in cellular genes leading to some of the phenotypic characteristics of tumor cells, but they also can stimulate inflammatory leukocytes to release superoxide. The release of superoxide by phagocytic cells following stimulation with phorbol esters is proportional to their tumor promoting activity. Low levels of both O2ċ − and H2O2, products of the “respiratory burst”, can promote fibroblast growth, possibly fibroblasts that harbor an oncogene or a mutated protooncogene. Also, low levels of superoxide can stimulate growth or growth responses in a variety of cell types when added exogenously to culture medium. In particular, these species stimulate the activation and the translocation of protein kinase C as well as the expression of early growth-regulated genes, such as the protooncogenes c-fos and c-myc. Superoxide and/or hydrogen peroxides might function as mitogenic stimuli through biochemical processes common to natural growth factors. Thus, signaling of growth responses involving released superoxide or hydrogen peroxide may be mediated through the oxidative modification of components of the signal transduction pathway. It is also possible that oxidative inactivation of serum protein inhibitors allows proteases to remodel the cell surface, thereby facilitating, or modulating, the action of normal growth factors (Burdon, Citation1993). Nonsteroidal anti-inflammatory drugs or COX-2 inhibitors have shown potent chemopreventive activity in animal colorectal carcinogenesis models (Cuendet & Pezzuto, Citation2000; Krishnan et al., Citation2000). Dietary polyphenolics, such as curcumin, chlorogenic acid, caffeic acid, resveratrol, or the flavonoid silymarin have also been shown to prevent colon carcinogenesis.
A final and decisive step in carcinogenesis is the invasion and metastatic spread of the tumor to various body spaces and cavities. This appears to be facilitated by the activation of genes for the release of proteolytic enzymes. Whereas high levels of immune cells appear to favor cell killing, lower numbers of immune cells can favor metastasis. Again, the release of superoxide may serve to promote metastatic growth. Alternatively, superoxide could inactivate serum antiproteases, some of which are extremely sensitive to oxidative inactivation (Floyd, Citation1990). Chemoprevention by flavonoids or polyphenolics could also result from tumor cell death, apoptosis, caused by the cytotoxic effect of flavonoids/polyphenolics. An accumulation of evidence has shown some anticancer properties of resveratrol are related to downregulating of the activation of NF-κB, which contributes to the progression of the androgen-independence of prostate cancer and increases invasive and metastatic properties (Tsai et al., Citation1999). Recently, another possible antitumor molecular mechanism of resveratrol was shown, which involves mitogen-activated protein kinase-mediated p53 activation and subsequent induction of apoptosis (She et al., Citation2002).
Lipid peroxidation is associated with some phases of carcinogenesis. There is increasing evidence that covalent binding of carcinogens or toxic substances to cellular macromolecules, particularly those carrying genetic information, is a primary event in the initiation of carcinogenesis. Thus, covalent binding to macromolecules could be the basis of many pathological changes induced by toxic substances. The ultimate forms of xenobiotics are believed to be reactive electrophilic metabolites, which combine with nucleophilic groups of macromolecules. It is also possible that miscoding or mutagenesis may be of minor importance in the initial events of chemical carcinogenesis and that genetic transpositions, including relatively large regions of the genome, may be more relevant (O'Brien, Citation1994). The DNA adducts, deoxyadenosine and deoxyguanosine, which are induced by malondialdehyde, the end-product of lipid peroxidation, accumulate in human breast cancer cells compared to normal breast cells (Wang et al., Citation1996). Serum antioxidative vitamin levels and lipid peroxidation were compared in gastric cancer patients (Choi et al., Citation1999). The level of serum ascorbic acid, α-tocopherol, β-carotene, and retinol were assessed. The levels of ascorbic acid in patients with gastric carcinoma were less that one-fifth of that in the control group, and the production of β-carotene and α-tocopherol were decreased, as well.
In summary, current information supports the importance of polyphenols in cancer protection (Dragsted, Citation2003; Rasmussen & Breinholt, Citation2003). A number of studies suggest that consumption of fruits and vegetables is associated with decreased risk of colon, breast, lung, stomach, and esophageal cancer (World Cancer Research Fund, 1997). Nonetheless, the overall incidence of cancers such as lung, breast, and prostate has increased. It is reasonable to expect that the incidence of cancer can be substantially reduced by diet modification, regular exercise, and avoiding tobacco smoke.
Cardiovascular and cerebrovascular disease
Polyphenolic compounds are generally known to possess antioxidant properties (Tapiero et al., Citation2002; Bub et al., Citation2003). Despite intake of a high-fat diet, the low incidence of coronary heart disease in France—the “French Paradox”—has been attributed partly to consumption of red wine (Sun et al., Citation2002). Compounds such as resveratrol, quercetin, catechin, and proanthocyanidins are enriched in grape skins and seeds, and the ability of these compounds to inhibit platelet aggregation and protect low-density lipoproteins from oxidation has been demonstrated (Xia et al., Citation1998). Also, the intake of flavonoids in relationship to cardiovascular disease has been explored by several groups of investigators. Hertog et al. (Citation1997) reported that a high intake of flavonols was associated with a decreased risk of coronary heart disease mortality. They also reported a general decrease in mortality with increasing flavonol intake. Knekt et al. (Citation1996) reported similar results, with a decreased risk of coronary mortality associated with flavonoids. In contrast, a U.S. study of heart disease in males 40–75 years of age found no significant association with flavonoid intake (Rimm et al., Citation1996). Finally, in a 10-year follow-up study of more than 34,000 postmenopausal women from Iowa, flavonoids intake was associated with a decreased risk of heart disease. However, no association was found between flavonoid intake and stroke mortality after 10 years of follow-up (Yochum et al., Citation1999).
The syndrome of ischemia /reperfusion (I/R) injury has been characterized for the heart, brain, intestine, kidney, and other organs. This phenomenon consists of a paradoxical increase in tissue injury during the reperfusion period in an organ that has sustained relatively minor damage during a period of ischemia. It is now evident that reperfusion tissue injury is mediated through oxidant mechanisms associated with the generation of oxygen-based radicals. ROS have been implicated in both the myocardial dysfunctions that are observed during reperfusion following short periods of ischemia (the stunned myocarium) and the irreversible injury to cardiac myocytes that occurs during reperfusion after longer periods of ischemia (Ferrari, Citation1994).
Infusions of high concentrations of the catecholamines epinephrine or norepinephrine into experimental animals are known to produce myocellular mitochondrial swelling, myofibrillar disruption, plasma membrane blebbing, and myocardial necrosis. It has been suggested that these cardiotoxic effects result not from the catecholamines themselves but from the production of and H2O2 formed by a complicated series of reactions during the autoxidation of catecholamines. It was observed that vitamin E–deficient rats were more sensitive to the cardiotoxic effects of isoproterenol, whereas myocardial damage induced by this synthetic catecholamine was reduced when the diet was supplemented with vitamin E. However, the results of studies demonstrating protection by antioxidants against catecholamine-induced myocardial necrosis must be interpreted with caution, as accumulation of neutrophils, a major source of oxygen radicals, has been observed in such models (Singal et al., Citation1982).
Carvedilol, a potent antioxidant, prevents the lipoperoxidation of mitochondrial membranes, which suggests a strong contribution to the known cardioprotective activity of this compound through protection of mitochondrial function (Moreno et al., Citation1998). A similar cardioprotective benefit is achieved by agents and antioxidant enzymes that scavenge hydroxyl radicals (or reduce their formation), but not agents that reduce superoxide anion production. Some examples of compounds of plant origin that have shown protective effects against ischemic injury are procyanidine from Vitis vinifera L. (Facino et al., Citation1996), resveratrol from red wine (Ray, Citation1999), and ginseng extract (Facino et al., Citation1999).
The ability of polyphenolic compounds to offer cardiovascular system protection has also stimulated efforts to investigate whether these compounds may offer neuroprotective effects. Few studies have explored flavonoid intake and risk of stroke. An inverse association has been shown with increasing dietary quercetin consumption. Tea consumption, which comprised the major source of flavonoids intake, was associated with decreased risk of stroke (Keli et al., Citation1996). Knekt et al. (Citation2000) showed that quercetin intake is not associated with cerebrovascular disease.
Lipid-peroxidation of biological membranes gives rise to degeneration of synapses and neurons and may be observed in stroke or neuronal disorders such as Alzheimer, Parkinson, and Huntington diseases. Oxidative stress and damage are accepted features of neural degeneration. The pathological presentation of Alzheimer disease, the leading cause of senile dementia, involves regionalized neuronal death and accumulation of intraneuronal and extracellular lesions (Smith et al., Citation1997). 4-Hydroxynonenal mediates oxidation-induced impairment of glutamate transport and mitochondrial function in synapses (Keller et al., Citation1997) Amyloid β-protein may be related to modulation of membrane lipid peroxidation. Amyloid β-protein fragment 25–35 [A-beta (25–35)] inhibits lipid peroxidation at low concentrations as a result of physicochemical interactions with the membrane lipid layer (Walter et al., Citation1997). Further, there is close association between increased levels of the antioxidant enzymes superoxide dismutase and heme oxygenase-1 and cytoskeleton abnormalities found in Alzheimer disease (Smith & Perry, Citation1998).
More studies will be needed to test specific effects of individual polyphenols and understand their molecular mechanism of action at the subcellular level during heart and neuronal diseases.
Discovery of new chemopreventive agents from plants
Botanicals have been used for the treatment of various human diseases throughout history. In addition to treatment, however, botanicals can play a role in disease prevention. There is growing interest in medicinal botanicals as part of complementary medicine in the United States. Approximately 40% of Americans use alternative remedies, including herbal medicine, for disease prevention and therapy (Eisenberg et al., Citation1998). The high cost, side effects, and therapeutic limitations of conventional medications are key factors that are driving the revival of herbal remedies. People can easily obtain herbal medicines or botanical supplements on an over-the-counter basis and feel comfortable consuming these products, even though safety and efficacy are generally not established on a rigorous scientific basis. This may be problematic due to factors such as drug-drug interactions (Go et al., Citation2001; von Gyuenigen & Hopkins, Citation2000). On the other hand, these preparations may mediate beneficial responses, either due to active chemical constituents or placebo effects.
During the past several years, we have instituted a multidisciplinary project wherein plant materials procured from throughout the world are used for activity-guided fractionation schemes that yield novel and otherwise unpredictable chemical entities with desirable biological potential. The overall experimental approach for procuring cancer chemopreventive agents in this manner has been described in the literature (Pezzuto, Citation1997; CitationPezzuto et al., 1998, 2004). In the field of natural product drug discovery, a time-tested method to enhance success is bioassay-guided fractionation. However, definition of the “best” bioassay system for monitoring cancer chemoprevention remains subjective. Use of in vitro assays enables the identification of a suitable number of lead starting materials, and bioassay-directed isolation facilitates the procurement of active agents from a reasonable number of sources in a finite period of time. There are many assays that can be used in drug discovery process. Several of them, which have been extensively used in our laboratory, are presented below.
Induction of quinone reductase activity utilizing cultured hepa 1c1c7 cells
Induction of phase II drug-metabolizing enzymes, such as glutathione S-transferase (GST) and quinone reductase (QR), is an important mechanism of cancer chemoprevention. In searching for novel cancer chemopreventive agents, we have used a rapid, sensitive QR assay to identify potential detoxification enzyme inducers (Talalay et al., Citation1995; Kang & Pezzuto, Citation2004). In order to understand the possible mechanism of induction, we have also employed a series of methods to further investigate induction patterns, protein expression, and mRNA expression, using transient transfection, Western blotting, Northern blotting, and reverse transcription-polymerase chain reaction (RT-PCR) techniques (Gerhäuser et al., Citation1997a; Song et al., Citation1999). Potent enzyme inducers are evaluated for potential to inhibit carcinogen-induced lesion formation in mouse mammary organ culture (MMOC) (Mehta et al., Citation1995). Chemopreventive activities of promising agents are studied for their inhibitory effects in animal models, such as the two-stage mouse skin or the rat mammary gland carcinogenesis models.
Antioxidant activity
Despite the abundance of published experimental approaches devoted to the antioxidant activity of polyphenols, only a few studies present reproducible information. Because reactive oxygen radicals play an important role in carcinogenesis, antioxidants present in consumable fruits, vegetables, and beverages have received considerable attention as cancer chemopreventive agents (CitationLee et al., 1998, 1999b). In order to identify antioxidants in plant extracts, activity has been assessed by determining scavenging activity with stable 2,2-diphenyl-1-picryhydrazyl (DPPH) free radicals (Fujita et al., Citation1988), inhibition of 12-O-tetradecanoyl-13-phorbol acetate (TPA)-induced free-radical formation with cultured HL-60 cells (Sharma et al., Citation1994), and inhibition of superoxide anion production in xanthine/xanthine oxidase systems (Sheu et al., Citation1998).
HL-60 cell differentiation
The HL-60 cell line has been used to provide a unique in vitro model system for studying the cellular and molecular events involved in the proliferation and differentiation of normal and leukemic cells. Terminal differentiation of human promyelocytic leukemia cells can be induced by a variety of chemical agents, and this process can be monitored readily by the generation of morphologically, histochemically, and functionally mature granulocytes and monocytes/macrophages (Suh et al., Citation1995; Mata-Greenwood et al., Citation2001). Induction of terminal differentiation is of interest for the therapy of leukemia, and human promyelocytic leukemia cell lines serve as convenient in vitro models to study differentiation. The usefulness of active compounds could be more far reaching, however, as substances such as vitamin A and D metabolites facilitate cell differentiation.
Inhibition of cyclooxygenase
Cyclooxygenase, a key enzyme in prostaglandin (PG) biosynthesis, catalyzes oxygenation of arachidonic acid to prostaglandin G2 (PGG2) and reduction of PGG2 to prostaglandin H2 (PGH2), an immediate precursor for production of eicosanoids. Arachidonic acid metabolites derived from PGH2 are important mediators of inflammatory responses, immunological effects, and tumor development. Several epidemiological studies have demonstrated that nonsteroidal anti-inflammatory drugs (NSAIDs) have cancer chemopreventive and tumor inhibitory effects in the human colon (Logan et al., Citation1993). Overproduction of PGs may influence tumor growth, carcinogen metabolism, and metastatic potential in human beings and experimental animals (Cuendet & Pezzuto, Citation2000). COX-2 is expressed in colorectal adenoma and carcinomas. Inhibition of COX-2 has been shown to decrease the incidence of carcinogen-induced neoplasia in rats and to reduce the incidence of adenomas in murine models. Several COX-2 inhibitors, with the potential for less toxicity than that associate with traditional NSAIDs, are currently available (Lynch, Citation2001).
Anti-estrogenic activity
For the evaluation of anti-estrogenic activities, we have employed an Ishikawa cell-based assay and an assay to determine inhibition of aromatase. The Ishikawa cell system has been employed to determine antiestrogenic activities (Pisha & Pezzuto, Citation1997). The Ishikawa cell line is a stable human endometrial carcinoma that expresses an estrogen-dependent alkaline phosphatase. Induction of this enzyme can be estimated kinetically with a microtiter plate reader. Thus, the assay is adaptable to large-scale screening using a 96-well plate format. In addition, the aromatase enzyme complex is responsible for the conversion of androgens to estrogens (Grubjesic et al., Citation2002). In the case of estrogen-dependent tumorigenesis, aromatase expression may be an important factor in the regulation of tumor growth. Therefore, inhibition of aromatase, the terminal step in estrogen biosynthesis, provides a method for treating hormone-dependent breast cancer (Santen & Harvey, Citation1999). Third-generation aromatase inhibitors have replaced megestrol acetate as second-line hormonal therapy in advanced breast cancer, and large clinical trials are designed to establish their efficacy, relative to tamoxifen (Ingle, Citation2001).
Unfortunately, no short-term in vitro systems are capable of fully representing the physiological complexity of mammals. Thus, to help overcome this disadvantage, we have assembled a panel of bioassay procedures that reflect various stages of carcinogenesis. The composition of the panel varies over time but generally includes assays that are suitable for monitoring inhibition of carcinogenesis at the stages of initiation (e.g., antioxidant activity and induction of quinone reductase activity in cell culture), promotion (e.g., inhibition of TPA-induced ODC activity in cell culture and inhibition of cyclooxygenase activity), and progression (e.g., induction of cell differentiation and anti-estrogenic activity). More than 15,000 tests have been performed, and the number of plants characterized as “active” is in the range of 3% of the total. In general, these “active” plant extracts are tested in a secondary model of greater physiological complexity. One example is a test to assess potential to inhibit carcinogen-induced preneoplastic lesion formation in the mouse mammary organ culture model (Mehta & Pezzuto, Citation2002; Pezzuto, Citation1997; CitationPezzuto et al., 1998, 2004). Active leads in the secondary model are subjected to bioassay-guided fractionation using an in vitro system as a monitor, and active isolates are considered for evaluation in full-term animal studies. Examples of edible plant isolates are summarized in .
Table 1 Potential chemopreventive agents isolated from edible plants.
As compared with edible plants that are used in daily life as a food, we consider medicinal plants as those with pharmacological activities to treat disease. There have been many scientific reports describing diverse clinical uses and bioactive components of medicinal plants. Animal trials to evaluate the chemopreventive activity of some well-known traditional medicinal plants are summarized in (Park & Pezzuto, Citation2002b). Various groups of compounds have been classified as cancer chemopreventive agents, largely based on the results of animal studies and epidemiological data. A series of studies to evaluate the chemopreventive activity of medicinal plants has been performed in Japan. The approach has typically involved primary screening of many natural products followed by a two-stage carcinogenesis assay to assess inhibitory effects on mouse skin tumors induced by the DMBA/TPA protocol. As a primary screen, compounds have been tested for their inhibitory effects on Epstein-Barr virus early antigen (EBV-EA) induction by treatment of Raji cell with a tumor promoter. Using a battery of chemopreventive assay systems, we have reported various candidate agents. Some chemopreventive agents from plants are summarized in .
Table 2 Chemopreventive medicinal plants reported in animal carcinogenesis models.
Table 3 Potential chemopreventive agents isolated from medicinal plants.
At present, brassinin (Mehta et al., Citation1995), sulforaphane and sulforamate (Gerhäuser et al., Citation1997a), 4′-bromoflavone (Song et al., Citation1999), withanolides (strong inducers of phase II drug-metabolizing enzymes) (Suh et al., Citation1995), deguelin (strong ODC inhibitor) (Kennelly et al., Citation1997), brusatol (HL-60 cell differentiation) (Mata-Greenwood et al., Citation2001), two aromatase inhibitors (Talalay et al., Citation1995), and resveratrol (Bhat et al., Citation2001; Bhat & Pezzuto, Citation2002) are undergoing further investigation as lead compounds. Clinical trials are anticipated in due course.
Conclusions
The results of studies outlined in this review provide a solid rationale for improvements in dietary habits, with special emphasis on enhanced consumption of fruits and vegetables. As described herein, there is substantial merit in this approach, as many fruits and vegetables have been found to contain polyphenols—active cancer chemopreventive agents.
In the past few years, proof-of-principle has been accomplished for cancer chemoprevention, and two agents, tamoxifen and celecoxib, are available as prescription drugs for the prevention of breast cancer and familial polyposis, respectively. These are tremendous breakthroughs. Nonetheless, a great deal of additional work is required. For example, dilemmas such as the near 100% probability of developing prostate cancer given a sufficient lifespan, and the lack of effectiveness of agents such as tamoxifen against estrogen receptor–negative breast cancer, need to be recognized. Some large-scale clinical trials are ongoing, but new cancer chemopreventive agents are still required, and plant materials are a promising source for the identification of active lead compounds. Certainly, to move from a crude starting material such as a plant extract to a prescription drug proven effective as a cancer chemopreventive agent is a long and arduous process. Nonetheless, it is necessary to bear in mind the significance of the task. The morbidity and mortality associated with metastatic cancer should be of concern for every living human being; no therapeutic option offers greater promise than cancer chemoprevention. Having the potential of administering an effective agent through the diet or as a prescription drug is a major advantage. This flexibility offers great hope for making significant progress in the near future.
Human trials on the biological effects of polyphenols have been limited. One problem is that it is difficult to determine which polyphenols are the most protective against cancers or cardiovascular diseases. Certain polyphenols inhibit COX, LOX, and phospholipase A2 activities, and they are considered potentially beneficial. Verification of the physiological relevance of such activities and the development of reliable methods to measure individual phenolic compounds in body fluids and tissues are necessary steps. Some polyphenolic compounds have been considered as chemopreventive agents on the basis of their ability to modulate signal transduction pathways, cell proliferation, and apoptosis. It is important to consider the concentrations of test agents used in these studies and assess the ability to conduct investigations needed in terms of comparison of the effective concentration on cell lines to attain comparable levels in animals and humans. It is also important to take into account the dose and mode of administering polyphenols when interpreting the results of the animal experiments. There is clearly a large amount of future research that remains to be done, but polyphenols offer great hope for the prevention of human disease.
Acknowledgments
Experimental work in the laboratory of the authors in the area of cancer chemoprevention is supported by grant no. P01 CA48112 awarded by the National Cancer Institute. Additional cancer chemopreventive testing is conducted under NCI contracts N01-CN-05024 (WS no. 73), N01-CN-05124 (WS no. 79), N01-CN-15017-44 (WS no. 80), and N01-CN-15017-44 (WS no. 85).
References
- Ahmed S, Rahman A, Mathur M, Athar M, Sultana S. 2001. Anti-tumor promoting activity of Asteracantha longifolia against experimental hepatocarcinogenesis in rats. Food Chem Toxicol. 39: 19–28. [PUBMED], [INFOTRIEVE], [CSA]
- American Institute of Cancer Research/World Cancer Research Fund.. Food, Nutrition, and the Prevention of Cancer: A Global Perspective.
- Banerjee S, Prashar R, Kumar A, Rao AR. 1996. Modulatory influence of alcoholic extract of Ocimum leaves on carcinogen-metabolizing enzyme activities and reduced glutathione levels in mouse. Nutr Cancer. 25: 205–217. [PUBMED], [INFOTRIEVE], [CSA]
- Bhat KPL, Pezzuto JM. 2002. Cancer chemopreventive activity of resveratrol. Ann NY Acad Sci. 957: 210–229. [PUBMED], [INFOTRIEVE], [CSA]
- Bhat KPL, Kosmeder II JW, Pezzuto JM. 2001. Biological effects of resveratrol. Antiox Redox Signal. 3: 1047–1064
- Blackburn WD, Jr, Heck LW, Wallace RW. 1987. The bioflavonoid quercetin inhibits neutrophil degranulation, superoxide production, and the phosphorylation of specific neutrophil proteins. Biochem Biophys Res Commun. 144: 1229–1236. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Bolton JL, Pisha E, Shen L, Krol ES, Iverson SL, Huang Z, van Breemen RB, Pezzuto JM. 1997. The reactivity of o-quinones which do not isomerize to quinone methides correlates with alkylcatechol-induced toxicity in human melanoma cells. Chem-Biol Interact. 106: 133–148. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Bravo L. 1998. Polyphenols: Chemistry, dietary sources, metabolism, and nutritional significance. Nutr Rev. l56: 317–333
- Bub A, Watzl B, Blockhaus M, Brivida K, Liegibel U, Muller H, Pool-Zobel BL, Rechkemmer G. 2003. Fruit juice consumption modulates antioxidative status, immune status and DNA damage. J Nutr Biochem. 14: 90–98. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Burdon RH, Gill V. 1993. Cellular generated active oxygen species and HeLa cell proliferation. Free Radic Res Commun. 19: 203–213. [PUBMED], [INFOTRIEVE]
- Byers T. 2002. The role of nutrition and nutrients in the prevention of chronic diseases. Prim Care. 29: 615–627. [PUBMED], [INFOTRIEVE]
- Chan TS, Galati G, Pannala AS, Rice-Evans C, O'Brien PJ. 2003. Simultaneous detection of the antioxidant and pro-oxidant activity of dietary polyphenolics in a peroxidase system. Free Radic Res. 37: 787–794. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Chang LC, Gerhäuser C, Song LL, Farnsworth NR, Pezzuto JM, Kinghorn AD. 1997. Activity-guided isolation of constituents of Tephrosia purpurea with the potential to induce the phase II enzyme, quinone reductase. J Nat Prod. 60: 869–873. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Chang LC, Bhat KP, Pisha E, Kennelly EJ, Fong HHS, Pezzuto JM, Kinghorn AD. 1998. Activity-guided isolation of steroidal alkaloid antiestrogen-binding site inhibitors from Pachysandra procumbens. J Nat Prod. 61: 1257–1262. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Chang LC, Gills JJ, Bhat KP, Luyengi L, Farnsworth NR, Pezzuto JM, Kinghorn AD. 2000a. Activity-guided isolation of constituents of Cerbera manghas with antiproliferative and antiestrogenic activities. Bioorg Med Chem Lett. 10: 2431–2434. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Chang LC, Song LL, Park EJ, Luyengi L, Lee KJ, Farnsworth NR, Pezzuto JM, Kinghorn AD. 2000b. Bioactive constituents of Thuja occidentalis. J Nat Prod. 63: 1235–1238. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Chewonarin T, Kinouchi T, Kataoka K, Arimochi H, Kuwahara T, Vinitketkumnuen U, Ohnishi Y. 1999. Effects of reselle (Hibiscus sabdariffa Linn.), a Thai medicinal plant, on the mutagenicity of various known mutagens in Salmonella typhimurium and on formation of aberrant crypt foci induced by the colon carcinogens azoxymethane and 2-amino-1-methyl-6-phenylimidazo [4,5-b]pyridine in F344 rats. Food Chem Toxicol. 37: 591–601. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Choi M-A, Kim B-S, Yu R. 1999. Serum antioxidative vitamin levels and lipid peroxidation in gastric carcinoma patients. Cancer Lett. 136: 89–93. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Chung HS, Chang LC, Lee SK, Shamon LA, van Breemen RB, Mehta RG, Farnsworth NR, Pezzuto JM, Kinghorn AD. 1999. Flavonoid constituents of Chorizanthe diffusa with potential cancer chemopreventive activity. J Agric Food Chem. 47: 36–41. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Cuendet M, Pezzuto JM. 2000. The role of cyclooxygenase and lipoxygenase in cancer research chemoprevention. Drug Metabol Drug Interact. 17: 109–157. [PUBMED], [INFOTRIEVE], [CSA]
- Decker EA. 1997. Phenolics: Prooxidants or antioxidants?. Nutr Rev. 55: 396–407. [PUBMED], [INFOTRIEVE], [CSA]
- Dragsted LO. 2003. Antioxidant actions of polyphenols in humans. Int J Vitam Nutr Res. 73: 112–119. [PUBMED], [INFOTRIEVE]
- Eisenberg DM, Davis RB, Ettner SL, Appel S, Wilkey S, Von Rompay M, Kessler RC. 1998. Trends in alternative medicine use in the United States, 1990-1997. JAMA. 280: 1569–1575. [PUBMED], [INFOTRIEVE], [CROSSREF]
- Facino RM, Carini M, Aldini G, Berti F, Rossoni G, Bombardelli E, Morazzoni P. 1996. Procyanidines from Vitis vinifera seeds protect rabbit heart from ischemia/reperfusion injury: Antioxidant intervention and/or iron and copper sequestering ability. Planta Med. 62: 495–502. [CSA]
- Facino RM, Carini M, Aldini G, Berti F, Rossoni G. 1999. Panax Ginseng administration in the rat prevents myocardial ischemia-reperfusion damage induced by hyperbaric oxygen: Evidence for an antioxidant intervention. Planta Med. 65: 614–619. [CSA], [CROSSREF]
- Ferrari R. 1994. Oxygen-free radicals at myocardial level: Effects of ischaemia and reperfusion. Adv Exp Med Biol. 366: 99–111. [PUBMED], [INFOTRIEVE]
- Floyd RA. 1990. Role of oxygen free radicals in carcinogenesis and brain ischemia. FASEB J. 4: 2587–2597. [PUBMED], [INFOTRIEVE]
- Fujita Y, Uehara I, Morimoto Y, Nakashima M, Hatano T, Okuda T. 1988. Studies on inhibition of mechanism of antioxidation by tannins and flavonoids. II. Inhibition of mechanism of coffee tannins isolated from leaves of Artemisia species on lipoxygenase dependent lipid peroxidation. Yakugaku Zasshi. 108: 129–135. [PUBMED], [INFOTRIEVE]
- Fukutake M, Yokota S, Kawamura H, Iizuke A, Amagaya S, Fukuda K, Komatsu Y. 1998. Inhibitory effect of Coptidis Rhizoma and Scutellariae Radix on azoxymethane-induced aberrant crypt foci formation in rat colon. Biol Pharm Bull. 21: 814–817. [PUBMED], [INFOTRIEVE], [CSA]
- Galati G, Teng S, Moridani MY, Chan TS, O'Brien PJ. 2000. Cancer chemoprevention and apoptosis mechanisms induced by dietary polyphenolics. Drug Metabol Drug Interact. 17: 311–349. [PUBMED], [INFOTRIEVE], [CSA]
- Gamez EJ, Luyengi L, Lee SK, Zhu LF, Zhou BN, Fong HHS, Pezzuto JM, Kinghorn AD. 1998. Antioxidant flavonoids glycosides from Daphniphyllum calycinum. J Nat Prod. 61: 706–708. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Gee JM, Johnson IT. 2001. Polyphenolic compounds: Interactions with the gut and implications for human health. Curr Med Chem. 8: 1245–1255. [PUBMED], [INFOTRIEVE]
- Gehm BD, McAndrews JM, Chien PY, Jameson JL. 1997. Resveratrol, a polyphenolic compound found in grapes and wine, is an agonist for the estrogen receptor. Proc Natl Acad Sci USA. 9: 14138–14143. [CROSSREF]
- Gerhäuser C, You M, Liu J, Moriaty RM, Hawthorne M, Mehta RG, Moon RC, Pezzuto JM. 1997a. Cancer chemopreventive potential of sulforamate, a novel analog of sulforaphane that induces phase 2 drug-metabolizing enzymes. Cancer Res. 57: 272–278. [PUBMED], [INFOTRIEVE]
- Gerhäuser C, Lee SK, Kosmeder JW, Moriaty RM, Hamel E, Mehta RG, Moon RC, Pezzuto JM. 1997b. Regulation of ornithine decarboxylase induction by deguelin, a natural product cancer chemopreventive agent. Cancer Res. 57: 3429–3435
- Go VLW, Wong DA, Resnick MS, Heber D. 2001. Evaluation of botanicals and dietary supplements therapy in cancer patients. J Nutr. 131: 179S–180S. [PUBMED], [INFOTRIEVE]
- Grubjesic S, Moriarty RM, Pezzuto JM. 2002. Aromatase inhibitors. Expert Opin Ther Patents. 12: 1647–1661. [CROSSREF]
- Gu J-Q, Park EJ, Hawthorne M, Mehta RG, Farnsworth NR, Pezzuto JM, Kinghorn AD. 2001. Constituents of Eugenia sandwicensis with potential cancer chemopreventive activity. Phytochemistry. 58: 121–127. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Havsteen BH. 2002. The biochemistry and medical significance of the flavonoids. Pharmacol Ther. 96: 67–202. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Herrmann KM. 1995. The shikimate pathway: Early steps in the biosynthesis of aromatic compounds. Plant Cell. 7: 907–919. [PUBMED], [INFOTRIEVE], [CROSSREF]
- Hertog MG, Hollman PC. 1996. Potential health effects of the dietary flavonol quercetin. Eur J Clin Nutr. 50: 63–71. [PUBMED], [INFOTRIEVE], [CSA]
- Hertog MGL, Feskens EJM, Kromhout D. 1997. Antioxidant flavonols and coronary heart disease risk: Ten year follow-up of the Zutphen Elderly Study. Lancet. 349: 699, [PUBMED], [INFOTRIEVE], [CROSSREF]
- Higdon JV, Frei B. 2003. Tea catechins and polyphenols: Health effects, metabolism, and antioxidant functions. Crit Rev Food Sci Nutr. 43: 89–143. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Hollman PCH, Feskens EJM, Katan MB. 1999. Tea flavonols in cardiovascular disease and cancer epidemiology. Proc Soc Exp Biol Med. 220: 198–202. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Ingle JN. 2001. Aromatase inhibition and antiestrogen therapy in early breast cancer treatment and chemoprevention. Oncology. 15: 28–34. [PUBMED], [INFOTRIEVE]
- Ito H, Miyake M, Nishitani E, Mori K, Hatano T, Okuda T, Konoshima T, Takasaki M, Kozuka M, Mukainaka T, Tokuda H, Nishino H, Yoshida T. 1999. Anti-tumor promoting activity of polyphenols from Cowania mexicana and Coleogyne ramosissima. Cancer Lett. 143: 5–13. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Itoigawa M, Ito C, Tan H T-W, Kuchide M, Tokuda H, Nishino H, Furukawa H. 2001. Cancer preventive agents, 4-phenylcoumarins from Calophyllum inophyllum. Cancer Lett. 169: 15–19. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Jeandet P, Douillet-Breuil AC, Bessis R, Debord S, Sbaghi M, Adrian M. 2002. Phytoalexins from the Vitaceae: Biosynthesis, phytoalexin gene expression in transgenic plants, antifungal activity, and metabolism. J Agric Food Chem. 50: 2731–2741. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Jeong HJ, Chang C, Kim HK, Kim IH, Kinghorn AD, Pezzuto JM. 2000. Aromatase inhibitors form Isodon excisus var. coreanus. Arch Pharm Res. 23: 243–245. [PUBMED], [INFOTRIEVE], [CSA]
- Kandaswami C, Middleton E, Jr. 1994. Free radical scavenging and antioxidant activity of plant flavonoids. Adv Exp Med Biol. 366: 351–176. [PUBMED], [INFOTRIEVE]
- Kang Y-H, Pezzuto JM. 2004. Induction of quinone reductase as a primary screen for natural product anticarcinogens. Meth Enzymol. 382: 380–414. [PUBMED], [INFOTRIEVE]
- Keli SO, Hertog MG, Feskens EJM, Kromhout D. 1996. Dietary flavonoids, antioxidant vitamins, and incidence of stroke: The Zutphen Study. Arch Int Med. 156: 637–642. [CSA]
- Keller JN, Mark RJ, Bruce AJ, Blanc E, Rothstein JD, Uchida K, Waeg G, Mattson MP. 1997. 4-Hydroxynonenal, an aldehydic product of membrane lipid peroxidation, impairs glutamate transport and mitochondrial function in synaptosomes. Neuroscience. 80: 685–696. [PUBMED], [INFOTRIEVE]
- Kennelly EJ, Gerhäuser C, Song LL, Graham JG, Beecher CWW, Pezzuto JM, Kinghorn AD. 1997. Induction of quinone reductase by withanolides isolated from Physalis philadelphica (tomatillos). J Agric Food Chem. 45: 3771–3777. [CROSSREF]
- Kim HS, Lee BM. 1997. Inhibition of benzo[a]pyrene-DNA adduct formation by Aloe barbadensis Miler. Carcinogenesis. 18: 771–776. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Kim DJ, Ahn B, Han BS, Tsuda H. 1997. Potential preventive effects of Chelidonium majis L. (Papaveraceae) herb extract on glandular stomach tumor development in rats treated with N-methyl-N′-nitro-N-nitrosoguanidine (MNNG) and hypertonic sodium chloride. Cancer Lett. 112: 203–208. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Kim HS, Kacew S, Lee BM. 1999. In vitro chemopreventive effects of plant polysaccharides (Aloe barbadensis Miller, Lentinus edodes, Ganoderma lucidum and Coriolus versicolor). Carcinogenesis. 20: 1637–1640. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Kitts DD, Yuan YV, Wijewickreme AN, Thompson LU. 1999. Antioxidant activity of the flaxseed lignan secoisolariciresinol diglycoside and its mammalian lignan metabolites enterodiol and enterolactone. Mol Cell Biochem. 202: 91–100. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Knekt P, Jarvinen R, Reunanen A, Maatela J. 1996. Flavonoid intake and coronary mortality in Finland: A cohort study. Br Med J. 312: 478–481
- Knekt P, Isotupa S, Rissanen H, Heliovaara M, Jarvinen R, Hakkinen S, Aromaa A, Reunanen A. 2000. Quercetin intake and the incidence of cerebrovascular disease. Eur J Clin Nutr. 54: 415–417. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Konoshima T, Takasaki M, Kozuka M, Tokuda H, Nishino H, Matsuda E, Nagai M. 1997. Anti-tumor promoting activities of isoflavonoids from Wistaria brachybotrys. Biol Pharm Bull. 20: 865–868. [PUBMED], [INFOTRIEVE], [CSA]
- Konoshima T, Takasaki M, Tokuda H, Nishino H. 2000. Cancer preventive activity of an iridoid glycoside, 8-acetylharpagide, from Ajuga decumbens. Cancer Lett. 157: 87–92. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Krishnan K, Ruffin MT, Brenner DE. 2000. Chemoprevention for colorectal cancer. Crit Rev Oncol Hematol. 33: 199–219. [PUBMED], [INFOTRIEVE], [CSA]
- Lairon D, Amiot MJ. 1999. Flavonoids in food and natural antioxidants in wine. Curr Opin Lipidol. 10: 23–28. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Lee SK, Mbwambo ZH, Chung H-S, Luyengi L, Gamez EJC, Mehta RG, Kinghorn AD, Pezzuto JM. 1998. Evaluation of the antioxidant potential of natural products. Comb Chem High Throughput Screen. 1: 35–46. [PUBMED], [INFOTRIEVE], [CSA]
- Lee SK, Luyengi L, Gerhäuser C, Mar W, Lee K, Mehta RG, Kinghorn AD, Pezzuto JM. 1999a. Inhibitory effect of munetone, an isoflavonoid, on 12-O-tetradecanoylphorbol 13-acetate-induced ornithine decarboxylase activity. Cancer Lett. 136: 59–65. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Lee SK, Song L, Mata-Greenwood E, Kelloff GJ, Steele VE, Pezzuto JM. 1999b. Modulation of in vitro biomarkers of the carcinogenic process by chemopreventive agents. Anticancer Res. 19: 35–44. [PUBMED], [INFOTRIEVE], [CSA]
- Lee D, Bhat KP, Fong HHS, Farnsworth NR, Pezzuto JM, Kinghorn AD. 2001c. Aromatase inhibitors from Broussonetia papyrifera. J Nat Prod. 64: 1286–1293. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Lee D, Cuendet M, Vigo JS, Graham JG, Cabieses F, Fong HH, Pezzuto JM, Kinghorn AD. 2001d. A novel cyclooxygenase-inhibitory stilbenolignan from the seeds of Alphanes aculeate. Org Lett. 3: 2169–2171. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Lee D, Park EJ, Cuendet M, Axelrod F, Chavez PI, Fong HHS, Pezzuto JM, Kinghorn AD. 2001e. Cyclooxygenase-inhibitory and antioxidant constituents of the aerial parts of Antirhea acutata. Bioorg Med Chem Lett. 11: 1565–1568. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Lee JM, Kwon H, Jeong H, Lee JW, Lee SY, Baek SJ, Surh YJ. 2001f. Inhibition of lipid peroxidation and oxidative DNA damage by Ganoderma lucidum. Phytother Res. 15: 245–249. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Logan RF, Little J, Hawlin PG, Hardcastle JD. 1993. Effect of aspirin and non-steroidal anti-inflammatory drugs on colorectal adenomas: Case-control study of subjects participating in the Nottingham fecal occult blood screening program. Br Med J. 307: 285–289
- Luyengi L, Suh N, Fong HHS, Pezzuto JM, Kinghorn AD. 1996. A lignan and four terpenoids from Brucea javanica that induce differentiation with cultured HL-60 promyelocytic leukemia cells. Phytochemistry. 43: 409–412. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Lynch PM. 2001. COX-2 inhibition in clinical cancer prevention. Oncology. 15: 21–26. [PUBMED], [INFOTRIEVE]
- Mata-Greenwood E, Ito A, Westenburg H, Cui B, Mehta MG, Kinghorn AD, Pezzuto JM. 2001. Discovery of novel inducers of cellular differentiation using HL-60 promyelocytic cells. Anticancer Res. 21: 1763–1770. [PUBMED], [INFOTRIEVE], [CSA]
- Mehta RG, Liu J, Constantinou A, Thomas CF, Hawthorne M, You M, Gerhäuser C, Pezzuto JM, Moon RC, Moriarty RM. 1995. Cancer chemopreventive activity of brassinin, a phytoalexin from cabbage. Carcinogenesis. 16: 399–404. [PUBMED], [INFOTRIEVE], [CSA]
- Mehta RG, Pezzuto JM. 2002. Discovery of cancer preventive agents from natural products: From plants to prevention. Curr Oncol Rep. 4: 478–486. [PUBMED], [INFOTRIEVE], [CSA]
- Middleton EJ. 1998. Effect of plant flavonoids on immune and inflammatory cell function. Adv Exp Med Biol. 439: 175–182. [PUBMED], [INFOTRIEVE], [CSA]
- Minguzzi S, Barata LES, Shin YG, Jonas PF, Chai HB, Park EJ, Pezzuto JM, Cordell GA. 2002. Cytotoxic withanolides from Acnistus arborescens. Phytochemistry. 59: 635–641. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Moreno AJ, Santos DJ, Palmeira CM. 1998. Ischemic heart disease: the role of mitochondria-carvedilol prevents lipid peroxidation of mitochondrial membranes. Rev Port Cardiol. 17(Suppl 2)II63–77. [PUBMED], [INFOTRIEVE]
- Morgan JF, Klucas RV, Grayer RJ, Abian J, Becana M. 1997. Complexes of iron with phenolic compounds from soybean nodules and other legume tissues: Prooxidant and antioxidant properties. Free Radic Biol Med. 22: 861–870. [CSA], [CROSSREF]
- O'Brien PJ. 1994. Antioxidants and cancer: Molecular mechanisms. Free radicals in diagnostic medicine. A systems approach to laboratory technology, clinical correlations, and antioxidant therapy. Armstrong D. Advances in Experimental Medicine and Biology. New York, Plenum Press. 366: pp. 215–239
- Park EJ, Pezzuto JM. 2002a. Autoxidation and antioxidants. Swarbrick J, Boylan JC. Encyclopedia of Pharmaceutical Technology, 2nd ed., New York, Marcel Dekker. pp. 97–113
- Park EJ, Pezzuto JM. 2002b. Botanicals in cancer chemoprevention. Cancer Metastasis Rev. 21: 231–255. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Pezzuto JM. 1997. Plant-derived anticancer agents. Biochem Pharmacol. 53: 121–133. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Pezzuto JM, Song LL, Lee SK, Shamon LA, Mata-Greenwood E, Jang M, Jeong HJ, Pisha E, Mehta RG, Kinghorn AD. 1998. Bioassay methods useful for activity-guided isolation of natural product cancer chemopreventive agents. Hostettmann K, Gupta MP, Marston A. Chemistry, Biological and Pharmacological Properties of Medicinal Plants from the Americas. ChurSwitzerland, Harwood Academic Publishers. pp. 81–110
- Pezzuto JM, Kosmeder II JW, Park EJ, Lee SK, Cuendet M, Gills J, Bhat K, Grubjesic S, Park HS, Mata-Greenwood E, Tan YM, Yu R, Lantvit DD, Kinghorn AD. 2004. Characterization of chemopreventive agents in natural products. Kelloff GJ, Hawk ET, Sigman CC. Strategies for Cancer Chemoprevention. Totowa, NJ, Humana Press
- Pisha E, Pezzuto JM. 1997. Cell-based assay for the determination of estrogenic and antiestrogenic activities. Meth Cell Sci. 19: 37–43. [CSA], [CROSSREF]
- Pool-Zobel BL, Bub A, Schroder N, Rechkemmer G. 1999. Anthocyanins are potent antioxidants in model systems but do not reduce endogenous oxidative DNA damage in human colon cells. Eur J Nutr. 38: 227–234. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Premalatha B, Sachdanandam P. 1999. Semecarpus anacardium L. nut extract administration induces the in vivo antioxidant defense system in aflatoxin B1 mediated hepatocellular carcinoma. J Ethnopharmacol. 66: 131–139. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Rajeshkumar NV, Kuttan R. 2001. Protective effect of picroliv, the active constituent of Picrorhiza kurroa, against chemical carcinogenesis in mice. Teratog Carcinog Mutagen. 21: 303–313. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Rasmussen SE, Breinholt VM. 2003. Non-nutritive bioactive food constituents of plants: Bioavailability of flavonoids. Int J Vitam Nutr Res. 73: 101–111. [PUBMED], [INFOTRIEVE]
- Ray PS, Maulik G, Cordis GA, Bertelli AAE, Bertelli A, Das DK. 1999. The red wine antioxidant resveratrol protects isolated rat hearts from ischemia reperfusion injury. Free Radic Biol Med. 27: 160–169. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Rimm EG, Katan MB, Ascherio A, Stampfer MJ, Willett WC. 1996. Relation between intake of flavonoids and risk for coronary heart disease in male health professionals. Ann Intern Med. 125: 384–389. [PUBMED], [INFOTRIEVE], [CSA]
- Roginsky V. 2003. Chain-breaking antioxidant activity of natural polyphenols as determined during the chain oxidation of methyl linoleate in Triton X-100 micelles. Arch Biochem Biophys. 414: 261–270. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Ross JA, Kasum CM. 2002. Dietary flavonoids: Bioavailability, metabolic effects, and safety. Ann Rev Nutr. 22: 19–34. [CSA], [CROSSREF]
- Saleem M, Alam A, Sultana S. 2001. Asafoetida inhibits early events of carcinogenesis: A chemopreventive study. Life Sci. 68: 1913–1921. [PUBMED], [INFOTRIEVE], [CROSSREF]
- Santen RJ, Harvey HA. 1999. Use of aromatase inhibitors in breast carcinoma. Endocr Relat Cancer. 6: 75–92. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Santos AC, Uyemura SA, Lopes JL, Bazon JN, Mingatto FE, Cutti C. 1998. Effect of naturally occurring flavonoids on lipid peroxidation and membrane permeability transition in mitochondria. Free Radic Biol Med. 24: 1455–1461. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Schwekendiek A, Pfeffer G, Kindl H. 1992. Pine stilbene synthase cDNA, a tool for probing environmental stress. FEBS Lett. 301: 41–44. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Sergediene E, Jonsson K, Szymusiak H, Tyrakowska B, Rietjens IM, Cenas N. 1999. Prooxidant toxicity of polyphenolic antioxidants to HL-60 cells: Description of quantitative structure-activity relationships. FEBS Lett. 462: 392–396. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Sharma S, Stutzman JD, Kelloff GJ, Steele VE. 1994. Screening of potential chemopreventive agents using biochemical markers of carcinogenesis. Cancer Res. 54: 5848–5855. [PUBMED], [INFOTRIEVE]
- She QB, Huang C, Zhang Y, Dong Z.. 2002. Involvement of c-jun NH(2)-terminal kinases in resveratrol-induced activation of p53 and apoptosis. Mol Carcinog. 33: 244–250. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Sheu S-Y, Lai C-H, Chiang H-C. 1998. Inhibition of xanthine oxidase by purpurogallin and silymarine group. Anticancer Res. 18: 263–268. [PUBMED], [INFOTRIEVE], [CSA]
- Siemann EH, Creasy LL. 1992. Concentration of the phytoalexin resveratrol in wine. Am J Enol Vitic. 43: 49–52
- Singal PK, Kapur N, Dhillon KS, Beamish RE, Dhalla NS. 1982. Role of free radicals in catecholamine-induced cardiomyopathy. Can J Physiol Pharmacol. 60: 1390–1397. [PUBMED], [INFOTRIEVE]
- Smith MA, Harris PL, Sayre LM, Perry G. 1997. Iron accumulation in Alzheimer's disease is a source of redox-generated free radicals. Proc Natl Acad Sci USA. 94: 9866–9868. [PUBMED], [INFOTRIEVE], [CSA]
- Smith MA, Perry G. 1998. The role of oxidative stress in the pathological sequelae of Alzheimer's disease. Free Radicals, Oxidative Stress, and Antioxidants. Pathological and Physiological Significance, Series A: Life Sciences. New York, Plenum Press. pp. 195–204
- Song LL, Kosmeder JW, Lee SK, Gerhäuser C, Lantvit D, Moon RC, Moriarty RM, Pezzuto JM. 1999. Cancer chemopreventive activity mediated by 4′-bromoflavone, a potential inducer of phase II detoxification enzymes. Cancer Res. 59: 578–585. [PUBMED], [INFOTRIEVE]
- Steele VE, Kelloff GJ, Balentine D, Boone CW, Mehta R, Baheri D, Sigman CC, Zhu S, Sharma S. 2000. Comparative chemopreventive mechanisms of green tea, black tea and selected polyphenol extracts measured by in vitro bioassays. Carcinogenesis. 21: 63–67. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Steinberg FM, Bearden MM, Keen CL. 2003. Cocoa and chocolate flavonoids: Implications for cardiovascular health. J Am Diet Assoc. 103: 215–223. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Stewart JR, Ward NE, Ioannides CG, O'Brian CA. 1999. Resveratrol preferentially inhibits protein kinase C-catalyzed phosphorylation of a cofactor-independent, arginine-rich protein substrate by a novel mechanism. Biochemistry. 38: 13244–13251. [PUBMED], [INFOTRIEVE], [CROSSREF]
- Su B-N, Chang LC, Park EJ, Cuendet M, Santarsiero BD, Mesecar AD, Mehta RG, Fong HHS, Pezzuto JM, Kinghorn AD. 2002a. Bioactive constituents of the seeds of Brucea javanica. Planta Med. 68: 1–4. [CROSSREF]
- Su B-N, Misico R, Park EJ, Santarsiero BD, Mesecar AD, Fong HHS, Pezzuto JM, Kinghorn AD. 2002b. Isolation and characterization of potential cancer chemopreventive principles of the leaves and stems of Physalis philadelphica. Tetradedron. 58: 3453–3466. [CROSSREF]
- Suh N, Luyengi L, Fong HHS, Kinghorn AD, Pezzuto JM. 1995. Discovery of natural products as chemopreventive agents utilizing HL-60 cell differentiation as a model. Anticancer Res. 15: 233–240. [PUBMED], [INFOTRIEVE]
- Sun AY, Simonyi A, Sun GY. 2002. The “French Paradox” and beyond: Neuroprotective effects of polyphenols. Free Radic Biol Med. 32: 314–318. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Takasaki M, Konoshima T, Kozuka M, Tokuda H. 1995. Anti-tumor-promoting activities of euglobals from Eucalyptus plants. Biol Pharm Bull. 18: 435–438. [PUBMED], [INFOTRIEVE], [CSA]
- Takasaki M, Konoshima T, Tokuda H, Masuda K, Arai Y, Shiojima K, Ageta H. 1999a. Anti-carcinogenic activity of Taraxacum plant I. Biol Pharm Bull. 22: 602–605. [PUBMED], [INFOTRIEVE], [CSA]
- Takasaki M, Tokuda H, Nishino H, Konoshima T. 1999b. Cancer chemopreventive agents (antitumor-promoters) form Ajuga decumbens. J Nat Prod. 62: 972–975. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Takasaki M, Konoshima T, Etoh H, Singh IP, Tokuda H, Nishino H. 2000. Cancer chemopreventive activity of euglobal-G1 from leaves of Eucalyptus grandis. Cancer Lett. 155: 61–65. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Talalay P, Fahey JW, Holtzclaw WD, Prestera T, Zhang Y. 1995. Chemoprevention against cancer by phase II enzyme induction. Toxicol Lett. 82: 173–179. [PUBMED], [INFOTRIEVE], [CROSSREF]
- Tapiero H, Tew KD, Ba GN, Mathe G. 2002. Polyphenols: Do they play a role in the prevention of human pathologies?. Biomed Pharmacother. 56: 200–207. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Terry P, Giovannucci E, Michels KB, Bergkvist L, Hansen H, Holmberg L, Wolk A. 2001. Fruit, vegetables, dietary fiber, and risk of colorectal cancer. J Natl Cancer Inst. 93: 525–533. [PUBMED], [INFOTRIEVE], [CROSSREF]
- Tsai SH, Lin-Shiau SY, Lin JK. 1999. Suppression of nitric oxide synthase and the down-regulation of the activation of NFκB in macrophages by resveratrol. Br J Pharmacol. 126: 673–680. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Udeani GO, Gerhäuser C, Thomas CF, Moon RC, Kosmeder JW, Kinghorn AD, Moriarty RM, Pezzuto JM. 1997. Cancer chemopreventive activity mediated by deguelin, a naturally occurring rotenoid. Cancer Res. 57: 3424–3428. [PUBMED], [INFOTRIEVE]
- Urso ML, Clarkson PM. 2003. Oxidative stress, exercise, and antioxidant supplementation. Toxicology. 189: 41–54. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Valenzuela A, Lagos C, Schmidt K, Videla LA. 1985. Silymarin protection against hepatic lipid peroxidation induced by acute ethanol intoxication in the rat. Biochem Pharmacol. 34: 2209–2212. [PUBMED], [INFOTRIEVE], [CROSSREF]
- Valenzuela A, Guerra R, Garrido A. 1987. Silybin dihemisuccinate protects rat erythrocytes against phenylhydrazine-induced lipid peroxidation and hemolysis. Planta Med. 53: 402–405. [PUBMED], [INFOTRIEVE]
- van Duyn MAS, Pivonka E. 2000. Overview of the health benefits of fruit and vegetable consumption for the dietetics professional: Selected literature. J Am Diet Assoc. 100: 1511–1521. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- von Gyuenigen VE, Hopkins MP. 2000. Alternative medicine in gynecologic oncology: A case report. Gynecol Oncol. 77: 190–192. [CSA], [CROSSREF]
- Walter MF, Mason PE, Mason RP. 1997. Alzheimer's disease amyloid β peptide 25-35 inhibits lipid peroxidation as a result of its membrane interactions. Biochem Biophys Res Commun. 233: 760–764. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Wang M, Dhingra K, Hittelman WN, Liehr JG, de Andrade M, Li D. 1996. Lipid peroxidation-induced putative malondialdehyde-DNA adducts in human breast tissues. Cancer Epidemiol Biomarkers Prev. 5: 705–710. [PUBMED], [INFOTRIEVE], [CSA]
- Westenburg HE, Lee KJ, Lee SK, Fong HHS, van Breemen RB, Pezzuto JM, Kinghorn AD. 2000. Activity-guided isolation of antioxidative constituents of Cotinus coggygria. J Nat Prod. 63: 1696–1698. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Xia J, Allenbrand B, Sun GY. 1998. Dietary supplementation of grape polyphenols and chronic ethanol administration on LDL oxidation and platelet function in rats. Life Sci. 63: 389–390. [CROSSREF]
- Yamanaka N, Oda O, Nagao S. 1997. Prooxidant activity of caffeic acid, dietary non-flavonoid phenolic acid, on Cu2+-induced low density lipoprotein oxidation. FEBS Lett. 405: 186–190. [PUBMED], [INFOTRIEVE], [CSA], [CROSSREF]
- Yochum L, Kushi LH, Meyer K, Folsom AR. 1999. Dietary flavonoids intake and risk of cardiovascular disease in postmenopausal women. Am J Epidemiol. 149: 943–949. [PUBMED], [INFOTRIEVE], [CSA]
- Yu L, Yu T, Ma R. 1995. Inhibition of the tumor promoting action of 12-O-tetradecanoylphorbol-13-acetate by tubeimoside III isolated from Bolbostemma paniculatum. Carcinogenesis. 16: 3045–3048. [PUBMED], [INFOTRIEVE], [CSA]
- Zheng GQ, Kenney PM, Zhang J, Lam LK. 1993. Chemopreventive of benzo[a]pyrene-induced forestomach cancer in mice by natural phthalides from celery seed oil. Nutr Cancer. 19: 77–86. [PUBMED], [INFOTRIEVE]