Abstract
A series of sulfonamide derivatives incorporating substituted 3-formylchromone moieties were investigated for the inhibition of three human carbonic anhydrase (hCA, EC 4.2.1.1) isoforms, hCA I, II, and VI. All these compounds, together with the clinically used sulfonamide acetazolamide, were investigated as inhibitors of the physiologically relevant isozymes I, II (cytosolic), and VI (secreted isoform). These sulfonamides showed effective inhibition against all these isoforms with KI’s in the range of 0.228 to 118 µM. Such molecules can be used as leads for discovery of novel effective CA inhibitors against other isoforms with medicinal chemistry applications.
Introduction
The involvement of the metalloenzyme carbonic anhydrase (CA; EC 4.2.1.1) in various physiological processes has been recognized for a long period, being shown that the deregulated expression and/or abnormal performance of the 16 presently known isozymes may have important pathological consequencesCitation1,Citation2. In fact, there are several human diseases whose pathopysiological characteristics include disbalance in the conversion between carbon dioxide and bicarbonate (the two substrates of these enzymes), resulting in perturbed ion transport, shift in pH, abnormal fluid secretion, etc.Citation1–4. Therefore, it seems plausible that modulation of CA activity to normal levels either by inhibition or activation offers interesting therapeutic optionsCitation1,Citation3. Clinical testing and/or use of the first CA inhibitors (CAIs) dates back several decades ago, to a period before the recognition of the diversity of isoforms within the CA family and their differential distribution in various human tissues and organsCitation1–4. Traditionally, the use of inhibitors was based on histochemical, biochemical, and functional evidences for the presence of active CAs, potentially contributing to disease, and consequences of the treatment were evaluated mainly symptomaticallyCitation1–4. Because of their favourable outcomes, sulfonamides became widely accepted drugs in the treatment of several CA-based diseases, especially as antiglaucoma agents, diuretics, and antiulcer agents among othersCitation2–4. However, systemic and even topically administered CA inhibitors regularly showed serious side effectsCitation2,Citation4–9. It is now understood that these undesired effects are due to the existence of at least 12 different active CA isoforms (together with the three inactive isozymes, the CA related proteins, CARPs VIII, X, and XI)Citation2,Citation4–10, that are indiscriminately inhibited irrespective of whether they play a real role in disease or are just coexpressed in the same tissue and elsewhere in the body. Moreover, certain drugs directed primarily against different CA unrelated targets may also inhibit activity of CAs. This may be exemplified by the antiinflammatory cyclooxygenase-2-selective drugs celecoxib and valdecoxib that show nanomolar affinity to several CA isoforms, but are generally well tolerated and give clinical responses in several disorders1–5. Thus, it is critically important to thoroughly characterize the affinity of different isozymes for sulfonamide CAIs, due to the wide range of applications of such drugs, and also to better understand the side effects due to inhibition of isozymes, which do not constitute the main target for a certain disease/applicationCitation1–10.
Our groups recently investigated the interaction of CA I and II isozymes with several types of phenols, pyrrole derivatized sulfonamides, dopaminergic bromophenolic compounds, antioxidant bisphenols and several of its substituted derivatives, e.g., salicyclates and some of their derivativesCitation8,Citation9. Here we extend these earlier investigations to series of sulfonamides, some of which are widely used as prodrug or as drugs. Sulfonamides possess many types of biological activities, and representatives of this class of pharmacological agents are widely used in clinic as antibacterial, hypoglycemic, diuretic, anti-hypertensive and antiviral drugs among othersCitation1–5. Recently, a host of structurally novel sulfonamide derivatives have been reported to show substantial antitumor activity in vitro and/or in vivoCitation5–9.
In the present study, we have purified human CA I, II, and VI (hCA I, hCA II, and hCA VI) isoenzymes and examined the in vitro inhibition effects of some sulfonamide compounds mentioned above on these enzymes, using the CO2 hydration activity of hCA I, hCA II, and hCA VI.
Materials and methods
CNBr-activated Sepharose 4B, protein assay reagents, p-aminobenzene sulfonamide L-tyrosine, 4-nitrophenylacetate and chemicals for electrophoresis were purchased from Sigma-Aldrich Co. All other chemicals were of analytical grade and obtained from either Sigma or Merck.
General method for the synthesis of compounds
A solution containing 0.001 mol 6-(un)substituted 3-formylchromone in 5–7 mL ethanol was stirred with heating until dissolved. Then catalytic amount of p-toluenesulfonic acid (p-TsOH) was added followed by the addition of 0.001 mol of 3- and 4-aminobenzene sulfonamide in equal volumes of ethanol. Reaction mixture was refluxed for 3.5 h and kept overnight. Solid product was obtained by filtration and purified by recrystallization from a mixture of hot ethanol and acetone (1:1)Citation11.
Kinetic studies
Purification of hCA I and hCA II were previously describedCitation8,Citation9. The methods proposed by Kivela et al. were modified and used for purification of CA VI from human serumCitation12–14. The activity assay was carried out according to Verpoorte et al.Citation15.
hCA-VI purification assay
Serum was obtained from fresh human blood at the Blood Centre of the Research Hospital at Atatürk University. The blood samples were centrifuged at 5000 rpm for 15 min, the precipitant was removed and serum was isolated. The pH was adjusted to 8.7 with solid Tris. Sepharose-4B-aniline-sulfanylamide affinity column was equilibrated with 25mM Tris-HCl/0.1M Na2SO4 (pH 8.7). The affinity gel was washed with 25mM Tris-HCl/22mM Na2SO4 (pH 8.7). The human carbonic anhydrase (hCA-VI) isozyme was eluted with 0.25 M H2NSO3H/25 mM Na2HPO4 (pH = 6,7). All procedures were performed at 4°CCitation12–14.
CA activity assay
Carbonic anhydrase activity was assayed by following the change in absorbance at 348 nm of 4-nitrophenylacetate (NPA) to 4-nitrophenylate ion over a period of 3 min at 25°C using a spectrophotometer (Shimadzu UV-VIS) according to the method described by Verpoorte et al.Citation15. The enzymatic reaction, in a total volume of 3.0 mL, contained 1.4 mL 0.05M Tris-SO4 buffer (pH 7.4), 1mL 3 mM 4-nitrophenylacetate, 0.5 mL H2O and 0.1mL enzyme solution. A reference measurement was obtained by preparing the same cuvette without enzyme solution. The inhibitory effects of the sulphonamide derivatives were examined. All compounds were tested in triplicate at each concentration used. Different concentrations of the compounds were used. Control cuvette activity in the absence of inhibitor was taken as 100%. For the compounds, Activity (%)-[Inhibitor] graphs were drawn. In these experiments, 4-nitrophenylacetate was used as substrate at five different concentrations (0.15–0.75 mM).
Protein determination
Protein quantity was determined spectrophotometrically at 595 nm according to the Bradford method during the purification steps, using bovine serum albumin as the standardCitation16.
SDS polyacrylamide gel electrophoresis
SDS polyacrylamide gel electrophoresis was performed after purification of the enzymes. It was carried out in 10 and 3% acrylamide for the running and the stacking gel, respectively, containing 0.1% SDS according to Laemmli procedureCitation17.
Results and discussion
Sulfonamide type inhibitors bind to CAs, with coordination to the Zn(II) ion from the enzyme active site by substituting the fourth, non-protein ligand, a water molecule or hydroxide ion, such as for example acetazolamide (AZA), a clinically used compound since 1954Citation6–10. The X-ray crystal structure has been extensively used for understanding the inhibition mechanism of CAIs. For example, for the adduct of hCA II with sulfamideCitation9,Citation10, it has been observed that the compound binds to CA by anchoring its NH moiety to the zinc ion of the enzyme active site, through a hydrogen bond, as well as through a second hydrogen bond to the NH amide of Thr199, an amino acid conserved in all α-CAs and critically important for the catalytic cycle of these enzymesCitation2,Citation4–10.
Our groups recently investigated the interactions of sulfonamides and some of their substituted derivatives with all mammalian carbonic anhydrase enzymesCitation8,Citation18–24, demonstrating some low micromolar/submicromolar inhibitors, as well as the possibility to design isozyme selective CAIs. The inhibition profile of various CA isozymes with this class of agents is very variable, with inhibition constants ranging from the millimolar to the submicromolar rangeCitation10. Thus, it seemed reasonable to us to extend the previous studiesCitation8,Citation10,Citation18–29, including in this investigation a series of Schiff’s bases obtained by condensing formylchromone with aminosulfonamidesCitation11.
The purification of the three CA isozymes was performed with a simple one step method by a Sepharose-4B-aniline-sulfanilamide affinity column chromatoghrapyCitation9,Citation18. The hCA I was purified, 102.6-fold with a specific activity of 875.12 EUmg−1 and overall yield of 52.42 %, hCA II was purified, 867.3-fold with a specific activity of 6970 EUmg−1 and overall yield of 63.8 % and hCA VI was purified, 86.9-fold with a specific activity of 247 EUmg−1 and overall yield of 38.5 %Citation5,Citation12–14. Inhibitory effects of these sulfonamides 1–10 on enzyme activities were tested under in vitro conditions.
We report here the inhibitory effects of sulfonamides 1–9 on the hydratase activity of hCA I, II and VI. The sulfonamide CAI acetazolamide AZACitation1–4 has been used as a negative control in our experiments, and for comparison reasons. The previous reports by al-Rashida et al.Citation11 investigated compounds 1–5 with bovine carbonic anhydrase (bCA). Data of show the following regarding inhibition of hCA I, II and VI with compounds 1–9 and AZA (as standard), by an hydratase assay, with CO2 as substrateCitation15:
Table 1. hCA I, II and VI inhibition data with compounds 1–10, by an esterase assay with 4-nitrophenylacetate as substrate.
(i) Against the slow cytosolic isozyme hCA I, compounds 1–9 behave as quite weak inhibitors, with Ki values in the range of 13.724–118.47 μM. Therefore, the chromone moiety does not strongly influence hCA I inhibitory activity. It is also interesting to note that derivative 1 was a better hCA I inhibitor, compared to compounds 2–4. This might indicate that hydrophobicity in the chromone moiety is favourable for the inhibition of hCA I. Acetazolamide AZA is also a medium hCA I inhibitor with this assay and Ki of 1.213 μM), whereas compounds 1 and 5 were more powerful inhibitors than AZA ().
Figure1. Structure of the tested compounds.
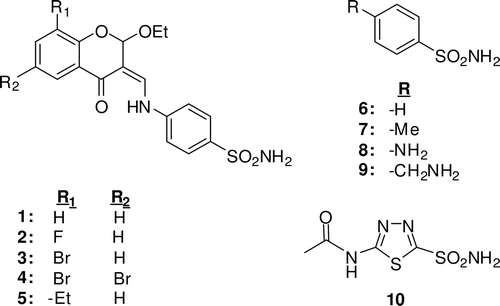
(ii) A better inhibitory activity has been observed with compounds 1–5 investigated here for the inhibition of the rapid cytosolic isozyme hCA II (). Two derivatives, i.e. 3–4, showed moderate hCA II inhibitory activity with Ki-s in the range of 1.134–1.863 μM, ), whereas the remaining derivatives were quite effective hCA II inhibitors (). The best hCA II inhibitor in this series of derivatives was the bulky, chromene derivative 1, which with a Ki of 0.228 μM, is a better inhibitor than AZA, a clinically used sulfonamide. Compounds 6–9, lacking the chromene moiety, were also moderate inhibitors for hCA II showing that the chromene moiety does not strongly influence the activity as in hCA I.
(iii) Chromene derivatives were also strong inhibitors of the secreted isozyme hCA VI, with Ki of 0.943–2.237 μM, except for 4, which had relatively weaker effect (Ki: 4.281). In general, the chromene derivatives showed a higher affinity for this isozyme, as compared with chromene lacking benzenesulfonamides 6 and 7 ().
(iv) All the tested compounds were much better inhibitors for human CA isoforms as compared with bovine CA reported by al-Rashida et al.Citation11.
Although there are several studies regarding the interactions of sulfonamide derivatives with carbonic anhydrase isoenzymes, it is critically important to explore further classes of potent CAIs in order to detect compounds with a different inhibition profile to find novel applications for the inhibitors of these widespread enzymes.
Especially, 4-{[(2-Ethoxy-4-oxo-2H-chromen-3(4H)-ylidene)methyl] amino}benzenesulfonamide 1 and 3-{[(6-Ethyl-2-ethoxy-4-oxo-2H-chromen-3(4H)- ylidene)methyl]amino}benzenesulfonamide 5 showed good activity against these hCA isozymes, more effective than the simple benzenesulfonamides 6–9. Findings of our study indicate another class of possible CAIs of interest, in addition to the well-known sulfonamides/sulfamates/sulfamides, the phenols/biphenyl diphenols bearing bulky ortho moieties in their molecules.
Declaration of interest
This study was financed by the Turkish Republic Prime Ministry State Planning Organization (DPT), (Project no: 2010K120440) and Agri Ibrahim Cecen University Scientific Research Council, (Project no: Agri BAP-2010/K-10) for (MS) and by an FP7 EU grant (Metoxia) to CTS. The authors report no conflicts of interest
References
- Supuran CT. Carbonic anhydrases: Novel therapeutic applications for inhibitors and activators. Nat Rev Drug Discov 2008;7:168–181.
- Weber A, Casini A, Heine A, Kuhn D, Supuran CT, Scozzafava A et al. Unexpected nanomolar inhibition of carbonic anhydrase by COX-2-selective celecoxib: New pharmacological opportunities due to related binding site recognition. J Med Chem 2004;47:550–557.
- Supuran CT. Carbonic anhydrase inhibitors. Bioorg Med Chem Lett 2010;20:3467–3474.
- Supuran CT. Carbonic Anhydrases: Catalytic Mechanism, Distribution and Physiological Roles. In Carbonic Anhydrase-Its Inhibitors and Activators. Supuran CT, Scozzafava A, Conway J, (eds.); CRC: Boca Raton (FL), USA, 2004; pp 1–24.
- Sly WS, Hu PY. Human carbonic anhydrases and carbonic anhydrase deficiencies. Annu Rev Biochem 1995;64:375–401.
- Parkkila S, Parkkila AK. Carbonic anhydrase in the alimentary tract. Roles of the different isozymes and salivary factors in the maintenance of optimal conditions in the gastrointestinal canal. Scand J Gastroenterol 1996;31:305–317.
- Pastorekova S, Parkkila S, Pastorek J, Supuran CT. Carbonic anhydrases: Current state of the art, therapeutic applications and future prospects. J Enzyme Inhib Med Chem 2004;19:199–229.
- Sentürk M, Gülçin I, Beydemir S, Küfrevioglu OI, Supuran CT. In Vitro Inhibition of Human Carbonic Anhydrase I and II Isozymes with Natural Phenolic Compounds. Chem Biol Drug Des 2011;77:494–499.
- Durdagi S, Sentürk M, Ekinci D, Balaydin HT, Göksu S, Küfrevioglu ÖI et al. Kinetic and docking studies of phenol-based inhibitors of carbonic anhydrase isoforms I, II, IX and XII evidence a new binding mode within the enzyme active site. Bioorg Med Chem 2011;19:1381–1389.
- Alterio V, Vitale RM, Monti SM, Pedone C, Scozzafava A, Cecchi A et al. Carbonic anhydrase inhibitors: X-ray and molecular modeling study for the interaction of a fluorescent antitumor sulfonamide with isozyme II and IX. J Am Chem Soc 2006;128:8329–8335.
- al-Rashida M, Ashraf M, Hussain B, Nagra SA, Abbas G. Discovery of new chromone containing sulfonamides as potent inhibitors of bovine cytosolic carbonic anhydrase. Bioorg Med Chem 2011;19:3367–3371.
- Kivelä J, Parkkila S, Waheed A, Parkkila AK, Sly WS, Rajaniemi H. Secretory carbonic anhydrase isoenzyme (CA VI) in human serum. Clin Chem 1997;43:2318–2322.
- Oztürk Sarikaya SB, Topal F, Sentürk M, Gülçin I, Supuran CT. In vitro inhibition of α-carbonic anhydrase isozymes by some phenolic compounds. Bioorg Med Chem Lett 2011;21:4259–4262.
- Senturk M, Ekinci D, Goksu S, Supuran CT. Effects of dopaminergic compounds on carbonic anhydrase isozymes I, II, and VI. J Enzyme Inhib Med Chem 2011; Doi:10.3109/14756366.2011.591290.
- Verpoorte JA, Mehta S, Edsall JT. Esterase activities of human carbonic anhydrases B and C. J Biol Chem 1967;242:4221–4229.
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 1976;72:248–254.
- Laemmli DK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970;227:680–685.
- Scozzafava A, Mastrolorenzo A, Supuran CT. Carbonic anhydrase inhibitors and activators and their use in therapy. Expert Opin Ther Pat 2006;16:1627–1644.
- Thiry A, Dogné JM, Supuran CT, Masereel B. Carbonic anhydrase inhibitors as anticonvulsant agents. Curr Top Med Chem 2007;7:855–864.
- Ekinci D, Ceyhun SB, Sentürk M, Erdem D, Küfrevioglu OI, Supuran CT. Characterization and anions inhibition studies of an a-carbonic anhydrase from the teleost fish Dicentrarchus labrax. Bioorg Med Chem 2011;19:744–748.
- Alp C, Ekinci D, Gültekin MS, Sentürk M, Sahin E, Küfrevioglu OI. A novel and one-pot synthesis of new 1-tosyl pyrrol-2-one derivatives and analysis of carbonic anhydrase inhibitory potencies. Bioorg Med Chem 2010;18:4468–4474.
- Ekinci D, Cavdar H, Talaz O, Sentürk M, Supuran CT. NO-releasing esters show carbonic anhydrase inhibitory action against human isoforms I and II. Bioorg Med Chem 2010;18:3559–3563.
- Ceyhun SB, Sentürk M, Ekinci D, Erdogan O, Ciltas A, Kocaman EM. Deltamethrin attenuates antioxidant defense system and induces the expression of heat shock protein 70 in rainbow trout. Comp Biochem Physiol C Toxicol Pharmacol 2010;152:215–223.
- Ceyhun SB, Sentürk M, Yerlikaya E, Erdogan O, Küfrevioglu OI, Ekinci D. Purification and characterization of carbonic anhydrase from the teleost fish Dicentrarchus labrax (European seabass) liver and toxicological effects of metals on enzyme activity. Environ Toxicol Pharmacol 2011;32:69–74.
- Supuran CT. Carbonic anhydrases–an overview. Curr Pharm Des 2008;14:603–614.
- Winum JY, Scozzafava A, Montero JL, Supuran CT. Design of zinc binding functions for carbonic anhydrase inhibitors. Curr Pharm Des 2008;14:615–621.
- Supuran CT, Mincione F, Scozzafava A, Briganti F, Mincione G, Ilies MA. Carbonic anhydrase inhibitors. Part 52. Metal complexes of heterocyclic sulfonamides: A new class of strong topical intraocular pressurelowering agents with potential use as antiglaucoma drugs. Eur J Med Chem 1998;33:247–254.
- Abbate F, Winum JY, Potter BV, Casini A, Montero JL, Scozzafava A et al. Carbonic anhydrase inhibitors: X-ray crystallographic structure of the adduct of human isozyme II with EMATE, a dual inhibitor of carbonic anhydrases and steroid sulfatase. Bioorg Med Chem Lett 2004;14:231–234.
- Renzi G, Scozzafava A, Supuran CT. Carbonic anhydrase inhibitors: Topical sulfonamide antiglaucoma agents incorporating secondary amine moieties. Bioorg Med Chem Lett 2000;10:673–676.