
Abstract
A small series of C-glycosides containing the methoxyaryl moieties was tested for the inhibition of the β-class carbonic anhydrases (CAs, EC 4.2.1.1) from Cryptococcus neoformans and Brucella suis. Many compounds showed activities in the micromolar or submicromolar range and excellent selectivity for pathogen CAs over human isozymes. The deprotected glycosides incorporating the 6-methoxy-2-naphthyl moiety showed the best inhibition profile and therefore represent leads for the development of novel anti-infectives with a new mechanism of action.
Introduction
Carbonic anhydrases (CAs) are a superfamily of zinc metalloenzymes that catalyze the reversible hydration of carbon dioxide to give bicarbonate and a protonCitation1. Whereas the α-CA family, mainly present in mammals has been thoroughly investigated from the drug design viewpointCitation2, only recently CAs belonging to the β- and γ-CA families, which are widespread in bacteria and fungi (the β-CAs) and Archaea (the β- and γ-CAs), respectively, started to be considered for such a purpose.
Cryptococcus neoformans is a basidiomycetes naturally found in soil contaminated with pigeon guano. Healthy individuals do not normally suffer from C. neoformans infections. However, upon inhalation into the alveoli of immunosuppressed individuals, for example AIDS patients and people undergoing chemotherapy, C. neoformans is able to colonize deep tissues and transverse into the nervous system where it can cause cryptococcal meningitisCitation3. The fungus experiences a dramatic raise in CO2 concentration during transitions from the natural environment to its mammalian host. This increase in CO2 promotes biosynthesis of a polysaccharide capsule, an important C. neoformans virulence factorCitation4. The CO2-sensing system of C. neoformans includes two prominent enzymes, the carbonic anhydrase Can2 and the fungal adenylyl cyclase Cac1. Can2, a member of β-CA family, is essential for survival of C. neoformans in its natural environment. Consistently, cultivation of C. neoformans on medium supplemented with CA inhibitors (CAIs) prevented growth of C. neoformans in low CO2 conditions typical for natural environmentsCitation5. Thus, this β-CA constitute attractive targets for development of antifungals that target horizontal transmission. Brucella sp. is a facultative intracellular Coccobacillus responsible for brucellosis, the major bacterial zoonosis worldwide, in a variety of mammals including ruminants and human. Human brucellosis may become chronic, eventually causing death. Virulence is linked to the capacity of the bacteria to replicate inside the macrophage host cell and to escape from the host immune system. The genome of the bacterial pathogen Brucella suis contains two CAs belonging to the β-class: bsCA I and bsCA IICitation6,Citation7. These two CAs were shown to be catalytically efficient, with activity for the CO2 hydration reaction similar to that of the human (h) isoform hCA II, and are inhibited by many sulfonamides/sulfamatesCitation8. Furthermore, certain sulfonamide CAIs were shown to inhibit the bacterial growth in cell culturesCitation8. Clearly parasite CAs may lead to anti-infectives with a new mechanism of action which bypasses the drug-resistance problems of clinically used agentsCitation9–11.
The use of glycomimetics in the design of CAIs has proven to be a successful approach and now constitutes one of the most attractive ways to develop new generations of effective and selective inhibitorsCitation12,Citation13. Recently our group has applied the “sugar approach” to the preparation of C-cinnamoyl phenols, where the carbohydrate moiety is tethered to a phenol CA pharmacophore through a carbon chainCitation14. These compounds have been tested as inhibitors of the Mycobacterium tuberculosis β-CAs and have shown better inhibitory activity against mtCAs than phenol. Also the antitubercular activity of the C-glycosyl phenols was investigated, allowing us to identify the first mtCAs inhibitor with antimycobacterial activityCitation15.
Very recently we developed a novel series of C-glycosides containing the methoxyaryl scaffold and investigated them as inhibitors against human isozymes of carbonic anhydrase, allowing us to identify four potent and highly selective inhibitors of hCA IX and XIICitation16. These results confirm that attaching carbohydrate moieties to CA methoxyaryl pharmacophore improves and enhances its inhibitory activity. It should be noted that only very-recently methoxyphenyl derivatives have been investigated as CAIs because it was considered that they do not bear any moiety normally associated with CA inhibition in their molecules.
Thus, in the search of non-sulfonamide CAIs belonging to different classes of compounds, we report here the synthesis of a series of C-glycosides incorporating the methoxyaryl moiety, and their inhibitory activity against the off-target hCA I and II, and C. neoformans β-CA encoded by the gene Can2, and Brucella suis β-CAs. This current study provides further grounding for the discovery of novel β-CA inhibitors which virtually can lead to new antibacterial or antifungal agents.
Materials and methods
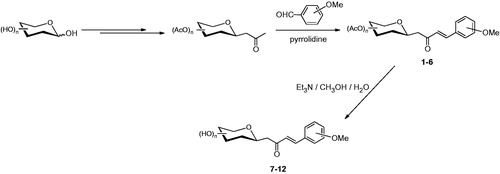
C-glycosides 1–12 were previously described, and have been prepared by aldol reaction of aryl alehydes with per-O-acetylated C-glucosyl or C-galactosyl ketones and subsequent deprotection using triethylamine in methanol/waterCitation16.
An Applied Photophysics stopped-flow instrument had been used for assaying the CA catalyzed CO2 hydration activityCitation17. Phenol red (at a concentration of 0.02 mM) had been used as indicator, working at the absorbance maximum of 557 nm, with 20 mM Hepes (pH 7.5) as buffer, and 20 mM Na2SO4 (for maintaining constant the ionic strength), following the initial rates of the CA-catalyzed CO2 hydration reaction for a period of 10–100 s. The CO2 concentrations ranged from 1.7 to 17 mM for the determination of the kinetic parameters and inhibition constants. For each inhibitor at least six traces of the initial 5–10% of the reaction have been used for determining the initial velocity. The uncatalyzed rates were determined in the same manner and subtracted from the total observed rates. Stock solutions of inhibitor (0.1 mM) were prepared in distilled-deionized water and dilutions upto 0.01 nM was done thereafter with distilled-deionized water. Inhibitor and enzyme solutions were preincubated together for 15 min at room temperature prior to assay, in order to allow for the formation of the E-I complex. The inhibition constants were obtained by non-linear least-squares methods using PRISM 3, and the Cheng-Prussoff equationCitation18 as reported earlier and represent the mean from at least three different determinations.
Results and discussion
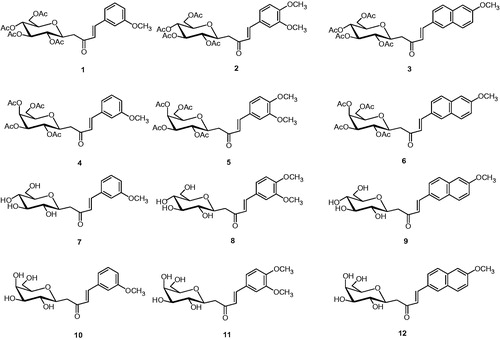
A set of C-cinnamoyl glycosides () was synthesized as outlined in Scheme 1 and described previously by usCitation14. C-cinnamoyl glycosides 1–6 have been prepared by aldol condensation of β-C-glucosyl and β-C-galactosyl ketones with 3-methoxybenzaldehyde, veratraldehyde or 6-methoxy-2-naphtaldehyde at room temperature in the presence of pyrrolidine as catalyst. The O-acetate protecting groups of the carbohydrate moiety were next removed using triethylamine in methanol/water to afford the deprotected C-glycosides 7–12.
Figure 1. Per-O-acetylated C-glycosides (1–6) and fully deprotected derivatives (7–12).
Scheme 1. Preparation of C-cinnamoyl glycosides 1–12.
The inhibitory activity of the C-glycosides 1–12 against human CA I and CA II and the purified pathogen β-CAs, Can2, bsCA I and bsCA II are presented in . These inhibition data were acquired using a stopped flow assay that monitors the physiological reaction, namely, the CA catalyzed hydration of CO2.
Table 1. Inhibition of mammalian α-CAs and pathogen β-CAs with the C-glycosides 1–12.a
A number of structure-activity relationships (SARs) were identified in this study and are summarized as follows.
(i) The cytosolic isoform hCA I was inhibited by most of the C-glycosides investigated here (i.e. 1–2, 4–8 and 10–12) in the micromolar range, with Ki values in the range of 4.67–8.69 µM. 6-Methoxy-2-naphthyl glucoside 3 and its deprotected derivative 9 are ineffective as hCA I inhibitors.
(ii) The C-glycosides showed a very interesting inhibition profile against hCA II. It should be noted that all glycosyl derivatives of veratraldehyde (i.e. 2, 5, 8 and 11) showed to be good inhibitors in the low micromolar range with Ki values in the range of 0.44–0.63 µM. Also compounds 1 and 3 are good hCA II inhibitors in the micromolar range. However being a ubiquitous, housekeeping isoform, this may not be a valuable property in another context if compounds targeting other isoforms should also possess activity against hCA II. Thus it is interesting to note that some of the novel compounds showed poor inhibition against hCA II while retaining good inhibition against pathogen CAs. C-galactosyl derivatives 4 and 6 and their deprotected analogues 10 and 12 were ineffective as inhibitors of hCA II. Also C-glucosides 7 and 9 showed a highly reduced activity against this isozyme.
(iii) The activity of the C-glycosides against C. neoformans β-CA (Can2) comprised compounds with three distinct inhibition profiles. The first group includes the compounds containing the veratrole moiety (i.e. 2, 5, 8 and 11) and exhibited very good Can2 inhibitory activity with inhibition constants in the range 0.22–1.59 µM. The second group (compounds 1, 3, 6, 7, 9 and 12) showed weaker Can2 inhibitory activity, with Kis in the range 2.37–8.10 µM. The third group includes the galactosyl derivatives of 3-methoxybenzaldehyde (4 and 10) which showed to be ineffective as inhibitors of Can2.
(iv) Brucella suis enzyme, bsCA I was inhibited by most of the C-glycosides investigated here (i.e. 1–3, 5, 7–9, 11 and 12) in micromolar or submicromolar range, with Ki values in the range of 4.56–0.42 µM. The per-O-acetylated C-galactosides 4 and 6, and the deprotected derivative 10 were ineffective as inhibitors of this pathogen isozyme. It is significant to note that the deprotected C-glycosides containing the 6-methoxy-2-naphthyl moiety (9 and 12) showed to be the most efficient inhibitors of bsCA I.
(v) The inhibition profile for bsCA II lays in two distinct groups. The first group includes compounds 1–3, 5, 8, 9, 12 with Kis of 0.31–0.70 µM while glycosides 4, 6, 7 and 10 (second group) were less effective bsCA II inhibitors with Kis in the range 4.02-8.69 µM. The deprotected galactoside containing the veratrole moiety 11 showed to be ineffective as inhibitor of bsCA II. As for bsCA I, the glycosides 9 and 12 were the best inhibitors of the Brucella suis isozyme II.
Selectivity for β-CAs against human CAs is an important consideration for downstream use of these C-glycosides novel probes that may help in the investigation and control of infectious diseases. The selectivity ratios of β-CA inhibition versus hCA I and hCA II inhibition are presented in . The clinically used CA inhibitor acetazolamide (AAZ) was less effective inhibitor for the bacterial and fungal β-CA enzymes. The reverse trend was observed for several of the glycosides reported here, with some compounds exhibiting a marked selectivity for the inhibition of the β-class CAs over the α-CAs. The selectivity ratios listed in show that C-glycosides 9 and 12 were up to several hundred-fold selective for pathogen CAs over human CA I and CA II and thus may represent leads for better discriminating the inhibition of β-CAs from pathogens. This observation provides a compelling opportunity to explore the 6-methoxy-2-naphthyl moiety scaffold in the development of potent and selective glycosyl inhibitors for the β-family of CAs. Clearly, the per-O-acetylated glycosides containing this scaffold (3 and 6) were less effective in this respect. C-glycoside 6 showed a maximum of 6-fold selectivity for pathogen CAs over human CA II and no selectivity for β-CAs over human CA I. On the other hand, C-glycoside 3 showed an excellent selectivity for the inhibition of β-CA enzymes over hCA I, but no selectivity for pathogen CAs over hCA II.
Table 2. Selectivity ratios of Ki for β-CAs compared to human α-CA isozymes I and II for the C-glycosides 1–12.a
In conclusion, we have investigated the enzyme inhibition profile of a series of C-glycosides incorporating the methoxyaryl moiety (compounds 1–12) against a panel of CAs encompassing the human α-CAs I and II, the fungal C. neoformans β-CAS and the pathogenic Brucella suis enzymes. The two best performing compounds, 9 and 12, preferentially inhibited pathogen CAs over human CAs and therefore represent leads for the development of novel anti-infectives with a new mechanism of action.
Declaration of interest
The authors report no declarations of interest. This work was financed in part by an EU grant (Dynano) to CTS, and by UNLP and CONICET (Argentina). P.A.C is member of the Scientific Research Career of CONICET.
References
- Supuran CT. Carbonic anhydrases: novel therapeutic applications for inhibitors and activators. Nat Rev Drug Discov 2008;7:168–81
- Supuran CT. Carbonic anhydrase inhibitors. Bioorg Med Chem Lett 2010;20:3467–74
- Supuran CT, Winum JY, Editors. Drug design of zinc-enzyme inhibitors: functional, structural, and disease applications. Hoboken: Wiley; 2009;1–1022
- Granger DL, Perfect JR, Durack DT. Virulence of Cryptococcus neoformans: regulation of capsule synthesis by carbon dioxide. J Clin Invest 1985;76:508–16
- Schlicker C, Hall RA, Vullo D, et al. Structure and Inhibition of the CO2-Sensing carbonic anhydrase Can2 from the pathogenic fungus Cryptococcus neoformans. J Mol Biol 2009;385:1207–20
- Joseph P, Turtaut F, Ouahrani-Bettache S, et al. Cloning, characterization, and inhibition studies of a β-carbonic anhydrase from brucella suis. J Med Chem 2010;5:2277–85
- Joseph P, Ouahrani-Bettache S, Montero JL, et al. A new β-carbonic anhydrase from Brucella suis, its cloning, characterization, and inhibition with sulfonamides and sulfamates, leading to impaired pathogen growth. Bioorg Med Chem 2011;19:1172–8
- Lopez M, Köhler S, Winum JY. Zinc metalloenzymes as new targets against the bacterial pathogen Brucella. J Inorg Chem 2012;111:138–45
- Capasso C, Supuran CT. Anti-infective carbonic anhydrase inhibitors: a patent and literature review. Expert Opin Ther Pat 2013;23:693–704
- (a) Maresca A, Scozzafava A, Vullo D, Supuran CT. Dihalogenated sulfanilamides and benzolamides are effective inhibitors of the three β-class carbonic anhydrases from Mycobacterium tuberculosis. J Enzyme Inhib Med Chem 2013;28:384–7. (b) Maresca A, Carta F, Vullo D, Supuran CT. Dithiocarbamates strongly inhibit the β-class carbonic anhydrases from Mycobacterium tuberculosis. J Enzyme Inhib Med Chem. 2013;28:407–11. (c) Singh S, Supuran CT. 3D-QSAR CoMFA studies on sulfonamide inhibitors of the Rv3588c β-carbonic anhydrase from Mycobacterium tuberculosis and design of not yet synthesized new molecules. J Enzyme Inhib Med Chem 2014;29:449–55
- Maresca A, Vullo D, Scozzafava A, Supuran CT. Inhibition of the alpha- and beta-carbonic anhydrases from the gastric pathogen Helycobacter pylori with anions. J Enzyme Inhib Med Chem 2013;28:388–91. (b) Del Prete S, De Luca V, Scozzafava A, et al. Biochemical properties of a new α-carbonic anhydrase from the human pathogenic bacterium Vibrio cholerae. J Enzyme Inhib Med Chem 2014;29:23–7. (c) Monti SM, Maresca A, Viparelli F, et al. Dithiocarbamates strongly inhibit the beta-class fungal carbonic anhydrases from Cryptococcus neoformans, Candida albicans and Candida glabrata. Bioorg Med Chem Lett 2012;22:859–62
- (a) Colinas PA. Novel glycomimetics: anomeric and N-glycosyl sulfonamides. Curr Org Chem 2012;16:1670–6. (b) Colinas PA, Nuñez NA, Bravo RD. Sulfonamidoglycosylation of methyl glycosides employing perchloric acid supported on silica gel. J Carbohydr Chem 2008;27:141–7. (c) Colinas PA, Bravo RD. Sulfonamidoglycosylation of methyl ribofuranosides: a novel approach to furanosylsulfonamides. Tetrahedron Lett 2005;46:1687–9
- Winum JY, Poulsen SA, Supuran CT. Therapeutic applications of glycosidic carbonic anhydrase inhibitors. Med Res Rev 2009;29:419–35
- Riafrecha LE, Rodríguez OM, Vullo D, et al. Synthesis of C-cinnamoyl glycosides and their inhibitory activity against mammalian carbonic anhydrases. Bioorg Med Chem 2013;21:1489–94
- Buchieri MV, Riafrecha LE, Rodríguez OM, et al. Inhibition of the β-carbonic anhydrases from Mycobacterium tuberculosis with C-cinnamoyl glycosides: Identification of the first inhibitor with anti-mycobacterial activity. Bioorg Med Chem Lett 2013;23:740–3
- Riafrecha LE, Rodríguez OM, Vullo D, et al. Attachment of carbohydrates to methoxyaryl moieties leads to highly selective inhibitors of the cancer associated carbonic anhydrase isoforms IX and XII. Bioorg Med Chem 2014;22:5308–14
- Khalifah RG. The carbon dioxide hydration activity of carbonic anhydrase. I. Stop-flow kinetic studies on the native human isoenzymes B and C. J Biol Chem 1971;246:2561–73
- Yung-Chi C, Prusoff WH. Relationship between the inhibition constant (KI) and the concentration of inhibitor which causes 50 percent inhibition (I50) of an enzymatic reaction. Biochem Pharmacol 1973;22:3099–108