Abstract
Allergic contact dermatitis is an important occupational and environmental disease caused by topical exposure to chemical allergens. An area of considerable interest and, in the context of hazard identification and characterization, an area of great importance is developing an understanding of the characteristics that confer on chemicals the ability to cause skin sensitization. For the successful acquisition of skin sensitization, it is necessary that a chemical must gain access to the viable epidermis, form stable immunogenic associations with host proteins, and provide the necessary stimuli for the activation, mobilization, and maturation of skin dendritic cells (DC). It is the last of these properties that is the subject of this article. The purpose here is to review the mechanisms through which skin sensitizers provide the triggers necessary for engagement of cutaneous DC. Of particular interest are the nature and function of danger signals elicited by skin sensitizing chemicals. Among the pathways considered here are those involving Toll-like receptors, C-type lectin receptors, neuropeptide receptors, prostanoid receptors, and the inflammasome. Collectively, danger signals in the skin provide a bridge between the innate and adaptive immune systems and are of pivotal importance for the initiation of cutaneous immune responses, including those to chemical allergens that result in skin sensitization.
Introduction
The skin is a complex and dynamic organ, functioning to provide physical and immunological defences against the external macro- and micro-environment. In fulfilling this role, the skin faces a myriad of environmental stresses and must therefore constantly adapt to maintain tissue homeostasis. The skin may be required to repair itself in response to injury, protect against pathogenic threats, or tolerate the non-pathogenic milieu (Gao et al., Citation2008). Sometimes, however, the response the skin provides is inappropriate and the balance is perturbed. One important example of this is contact hypersensitivity (CHS).
Contact hypersensitivity is an inflammatory skin condition caused by an inappropriate and unnecessary immune response to specific chemicals. These chemicals, which are collectively termed contact allergens, induce allergic sensitization that may result in contact dermatitis (ACD). This is a debilitating disease characterized by the development of redness, vesicles, papules, and eventually dry, scaly skin (Saint-Mezard et al., Citation2004; Thyssen et al., Citation2007). Significantly, a recent epidemiological study of the general population of North America and Western Europe suggests that the prevalence of contact allergy to at least one sensitizing chemical is ~ 20% (Thyssen et al., Citation2007). Thus, contact allergy is widely recognized clinically as a common and important environmental and occupational health hazard.
The requirement to test novel topical consumer products for their capacity to induce skin sensitization is evident. The standard approach to hazard identification and characterization is based upon the use of guinea pig or mouse assays. However, recent legislation seeks to limit the use of in vivo tests. Such legislation, when combined with ethical, societal, and scientific imperatives, has resulted in a significant investment in the design of alternative test methods that, it is hoped, will obviate the need for experimental animals. Clearly, success in this area will demand a detailed understanding of the biology that shapes cutaneous immune responses, and, in particular, the signals that drive the acquisition of skin sensitization.
The development of skin sensitization: A role for dendritic cells
In order for a chemical to induce skin sensitization, it must satisfy certain requirements (Kimber et al., Citation2002). First, the chemical must pass through the stratum corneum and gain access to the viable epidermis. The chemical must then bind to skin proteins, forming a hapten-protein conjugate and thus acquiring immunogenicity. The hapten-protein conjugate must be provided with the opportunity to stimulate the activation of responsive T-lymphocytes. For this to be achieved, the antigen must be internalized in the skin and transported to regional lymph nodes (LN) by cutaneous dendritic cells (DC).
Arguably, DC represent the most important initiators and regulators of not only skin sensitization but of immune responses per se. These cells play crucial roles in transferring information about the environment to the adaptive immune system. Due to the pivotal role of DC in supporting the development of skin sensitization, a number of proposed in vitro strategies for the identification and characterization of skin sensitizing chemicals have been suggested that are based upon exploiting our understanding of cultured DC or DC-like cells. Therefore, DC are important not only in the context of developing a more detailed understanding of the cell and molecular mechanisms that initiate and regulate skin sensitization, but also from the practical perspective of novel test development (Kimber et al., Citation2011).
Within mouse skin, there are at least three distinct subsets of DC; namely Langerin+ve CD103+ve dermal DC (dDC), Langerin−ve CD103−ve dDC, and Langerin+ve, CD103−ve Langerhans cells (LC) (Kaplan, Citation2010) (). Langerhans’ cells are the only DC subset to reside in the epidermis and, as such, were once considered to be wholly responsible for the presentation of chemical allergen to hapten-specific T-lymphocytes. However, more recent studies have challenged this belief, with the suggestion that all three subsets may, under certain circumstances, make distinct and opposing antigen-specific contributions to the induction and regulation of sensitization (Igyarto et al., Citation2011).
Table 1. The phenotypic markers present on the surface of mouse Langerhans’ cells, mouse Langerin-positive (+ve) dermal DC and mouse Langerin-negative (−ve) DC.
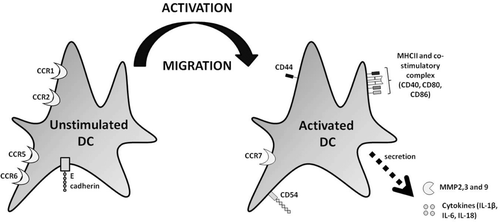
As indicated above, the development of skin sensitization requires that cutaneous DC are activated and mobilized (). To this end, it is necessary for a number of changes in DC phenotype to be induced. In general, these are changes that aid passage of the cell through the dermal layers, drive migration towards the LN, or facilitate antigen presentation (Merad et al., Citation2002). Under-pinning such cellular changes are induced changes in chemokine and cytokine expression. Of particular importance are the cytokines tumor necrosis factor (TNF)-α and interleukin (IL)-1β. After skin exposure to chemical allergens, TNFα and IL-1β are both rapidly up-regulated, with TNFα produced mainly by keratinocytes (KC) and IL-1β produced exclusively by LC (in mice) (Enk and Katz, Citation1992). In addition, recent evidence indicates that IL-18, a cytokine produced by both LC and KC, has the capacity to induce TNFα and IL-1β dependent LC mobilization (Cumberbatch et al., Citation2005). Overall, it can be concluded that IL-18, TNFα, and IL-1β are all integral to the process of LC mobilization, migration, and functional maturation.
Figure 1. Changes in DC marker expression following activation. In the resting state, DC express high levels of CCR1, CCR2, CCR5, CCR6, and E-cadherin. Upon activation, these markers are down-regulated and a number of other markers are up-regulated, including CCR7, CD40, ICAM1, MHC-II, and the co-stimulatory complex (CD40, CD80, and CD86). In addition, metalloproteinase (MMP)-2, MMP-3, MMP-9, and the cytokines IL-1β, IL-6, and IL-18 are all released. These changes facilitate mobilization, maturation, and migration of DC away from the skin and towards the LN.
Following migration into the LN, DC present allergen to allergen-specific T-lymphocytes. This causes a proliferation of Type 1 T-lymphocytes, specifically inducing the expansion of allergen-specific cytotoxic T-cell (Tc) 1 effectors and T helper (TH)-1 regulatory cells (Kehren et al., Citation1999).
The importance of Caspase-1
In order for IL-1β to induce DC activation, it must first be secreted from the cell and into the extracellular milieu. Initially, the IL-1β protein is produced as an inactive precursor called pro-IL-1β. The activation and release of this cytokine and the additional cytokine precursors pro-IL-18 and pro-IL-33 depends upon cleavage within the cytosol. This cleavage is performed by caspase-1. The importance of the caspase-1 enzyme to the development of contact sensitization was demonstrated by Antonopoulos et al. (Citation2001), who observed that, unlike the LC of wild type (WT) mice, the LC of caspase-1 deficient mice were unable to migrate in response to administration of the allergens 2,4-dinitroflurobenzene (DNFB) and oxazolone. Similarly, the capase-1 inhibitor Ac-YVAD-cmk was shown to inhibit LC migration in WT mice. The relevance of IL-1β to this process was confirmed, as local administration of exogenous IL-1β was able to restore LC migration in mice lacking caspase-1. Taken together, the evidence indicates that the development of skin sensitization is dependent on the action of the caspase-1 enzyme (Antonopoulos et al., Citation2001).
As with IL-1β, caspase-1 takes the form of an inactive precursor in the steady state and, thus, it must also be activated before it can function. This activation requires a pro-inflammatory trigger (Franchi et al., Citation2009). As caspase-1 activation is necessary for IL-1β secretion and IL-1β is required for the development of skin sensitization, it is clear that this pro-inflammatory trigger is necessary to support sensitization.
Cumberbatch et al. (Citation1993) first postulated a requirement for a pro-inflammatory trigger in the development, or optimal development, of skin sensitization. They found that the topical administration of 0.1% 2,4-dinitrochlorobenzene (DNCB), together with the non-sensitizing skin irritant sodium lauryl sulfate, enhanced DC migration and LNC proliferation when compared with responses induced by 0.1% DNCB alone. The interpretation was that by using low doses of DNCB that had only modest sensitizing potential (but little irritant activity), the vigor of cutaneous immune responses could be augmented by provision of independent pro-inflammatory signals. Grabbe et al. (Citation1996) went on to demonstrate that the topical co-administration of a sub-optimal dose of oxazalone with a sub-optimal dose of trinitrochlorobenzene (TNCB) induced the acquisition of sensitization. From these findings, it was concluded that induction of skin sensitization requires both an antigen-specific signal and an antigen non-specific pro-inflammatory signal. More recently, this antigen non-specific pro-inflammatory signal has been termed a ‘danger signal’.
The concept of danger signals was first articulated by Matzinger (Citation1994). She postulated that, instead of simply reacting to foreign antigens, the immune system has the potential to detect tissue damage. One benefit of this is that it serves to prevent the activation of adaptive immune responses to ‘harmless’ antigens, that is challenges that are not associated with tissue damage or trauma. Subsequently, such danger signals have been shown to be pivotal in bridging between the innate and adaptive immune systems. In this context, danger signals provide a mechanism through which DC can sense their environment and respond accordingly. In the remainder of this article, those danger signals that might be of particular relevance to skin DC and to the induction and orchestration of skin sensitization will be considered.
Danger signals and contact sensitization
Danger signals comprise a diverse group of molecules that are associated with the invasion of a foreign antigen. Typically, these danger signals are either a product of an invading pathogenic micro-organism, or a product of the damage that invasion creates. As such, danger signals can be divided up into two broad groups; pathogen-associated molecular patterns (PAMP) or damage-associated molecular patterns (DAMP). The former are, as their name suggests, molecules associated with the invading pathogen itself, whereas DAMP are endogenous molecules which are released in response to injury.
The integral inflammasome
One important group of PAMP receptors are the nucleotide-binding domain leucine-rich repeat containing receptors (NLR). The NLR family of proteins consists of 23 intracellular PAMP sensors including NLRP3, NLRP12, NLRC4, and NLRP1 (Franchi et al., Citation2009). Biochemical analyses have suggested that, upon activation, the NLR forms a complex with apoptosis associated speck-like protein containing a CARD (ASC). This complex is called the inflammasome. The inflammasome induces activation of the enzyme caspase-1, an enzyme that, as discussed above, is central to the development of skin sensitization. Consequently, it can be inferred that both the NLR family of pattern recognition receptors (PRR) are critical for danger signal dependent–caspase-1 activation and, moreover, that the NLR family of PRR may be integral to the development of skin sensitization.
The importance of the inflammasome to the development of skin sensitization can be demonstrated experimentally. For instance, it has been shown that ASC and NLRP3 deficient mice display an impaired response to the skin sensitizers DNFB and TNCB (Watanabe et al., Citation2007). More recently, it was found that the LC of NLRP12−/− mice exhibit defective migration to draining LN (Arthur et al., Citation2010). In this latter study, it was found also that NLRP12 deficiency attenuates the inflammatory response in two separate models of skin sensitization. It is relevant, therefore, to consider the mechanisms through which inflammasomes are activated.
NLRP3
The most comprehensively studied NLR is NLRP3. As described above, evidence suggests that this inflammasome is, in some instances, critical for the development of sensitization. The NLRP3 inflammasome is also implicated in a variety of different disease states and, thus, it is relevant to consider the role of this inflammasome in the context of skin sensitization.
The study of NLRP3 has revealed a multitude of diverse stimuli that are capable of inducing inflammasome activation. Included in this list are the crystalline compounds adenosine triphosphate (ATP), silica, asbestos and uric acid, the bacterial products listeriolysin O, potassium ionophore nigericin, aerolysin, and hemolysins, as well as the marine toxin maitotoxin (Pedra et al., Citation2009). Although many different substances have been shown to activate NLRP3, a direct ligand for this inflammasome has yet to be identified. However, a number of NLRP3 stimuli-induced cellular changes that may induce NLRP3 activation have been identified.
One theory, based upon evidence that many NLRP3 stimulants induce pore formation, suggests that pore formation itself can cause inflammasome activation. Building upon this, two additional theories have emerged. One proposes that membrane damage leads to microbes entering the cell and activating NLRP3 (Kanneganti et al., Citation2007), whereas the other suggests that membrane damage leads to the release of endogenous molecules that can then directly activate the NLRP3 (Ogura et al., Citation2006). In support of the latter theory, studies have demonstrated that endosomal membrane damage by the crystalline molecules alum, silica, and amyloid-β causes the release of cathepsin B. As cathepsin B activates the inflammasome, crystalline molecules may indeed activate NLRP3 by inducing endosomal membrane damage (Hornung et al., Citation2008). Nevertheless, other investigations suggest that this mechanism of NLRP3 activation is not common to all activating molecules. Indicative of this is evidence suggesting that ATP, unlike the crystalline compounds, induces pore formation by binding to the plasma membrane P2X7 receptor. It may be, therefore, that a single, unifying mechanism of NLRP3 inflammasome activation does not exist.
Interestingly, one study postulates that potassium efflux may be associated with toxin and crystal mediated NLRP3 activation (Petrilli et al., Citation2007). In that investigation, it was shown that inhibition of the efflux of potassium effectively inhibited NLRP3 activation. Furthermore, in vitro studies have demonstrated that inflammasome assembly and recruitment of caspase-1 occurs spontaneously at low intracellular potassium concentrations. Taken together, it was concluded that potassium efflux is a necessary mechanism for the assembly of NLRP3 (Petrilli et al., Citation2007). However, characterization of the precise mechanism of potassium efflux-induced NLRP3 assembly requires further investigation. Equally intriguing is evidence that postulates that stimulation of the P2X7 receptors by ATP may lead to the opening of a potassium-selective pore. As this will cause potassium efflux, ATP may activate the NLRP3 inflammasome via this process (Khakh and North, Citation2006). In support of this hypothesis, mice deficient in P2X7 receptor fail to mount a skin sensitization response to TNCB. However, the skin sensitization was restored when the P2X7 receptor-deficient mice were injected intra-dermally with IL-1β (Weber et al., Citation2010). Taken together, these data suggest that it is likely that P2X7-dependent IL-1β production is an important aspect of skin sensitization.
Another process commonly associated with toxin and crystal mediated NLRP3 activation is the production of reactive oxygen species (ROS) (Dostert et al., Citation2008). Importantly, many in vitro studies have revealed that ROS production is induced by contact allergens (Bruchhausen et al., Citation2003; Mehrotra et al., Citation2005; Martin et al., Citation2011). In human monocytes, the inhibition of ROS production, as achieved by administration of the NADPH oxidase inhibitor diphenyleneiodonium (DPI), inhibits ATP-mediated inflammasome activation (Hewinson et al., Citation2008). In addition, a variety of studies have reported increases in cellular ROS production after stimulation with both toxin and crystalline NLRP3 activators (Martinon, Citation2010). As mitochondria are believed to be the main source of cellular ROS, it is proposed that mitochondria may have a role to play in NLRP3 activation. Zhou et al. (Citation2011) showed in support of this that damaged, ROS-generating mitochondria are capable of stimulating activation of NLRP3. The same authors also found that depletion of the mitochondrial membrane protein voltage-dependent anion channel, a protein required for mitochondrial ROS production, impaired the activation of the NLRP3 inflammasome. Collectively, the available evidence suggests that activation of the NLRP3 inflammasome may be dependent upon ROS production, which, may in turn be driven by mitochondrial function. However, as with potassium efflux induction, the molecular mechanisms that cause ROS production are not thoroughly understood.
NLRP12
The NLRP12 inflammasome is less well-characterized. As described above, this inflammasome has also been shown to be implicated in the development of skin sensitization. However, in contrast to other NLR, NLRP12 appears to negatively regulate the production of IL-1β (Shaw et al., Citation2010). It has been shown that this inflammasome prevents phosphorylation of IL-1 receptor-associated kinase 1 (IRAK-1) and enhances degradation of the transcription factor nuclear factor κ-light-chain-enhancer of activated B-cell (NF-κB) inducing kinase (NIK). As the negative regulation of IL-1β by NLRP12 would effectively inhibit LC migration and DC accumulation, the presumption is that this mechanism would prevent the acquisition of skin sensitization. However, as NLRP12 is known to be required for the development of skin sensitization to certain allergens, it is postulated that there must be some, as yet unknown mechanism for the observed dependence upon NLRP12 (Arthur et al., Citation2010).
NLRC4 and NLRP1
Other NLR implicated in the activation of pro-inflammatory cytokines are the NLRC4 and NLRP1 inflammasomes. Although these inflammasomes have yet to be directly implicated in the development of skin sensitization, their mechanisms of action suggest that they may also have a role to play. NLRC4 activation is induced by flagellin and the inner rod component of Type III bacterial secretion systems (Zhao et al., Citation2011). A recent study demonstrated that the intracellular sensor Naip5 is required for flagellin-dependent activation of NLRC4, whereas the intracellular sensor Naip2 is involved in detecting the inner rod component of Type III secretion systems (Kofoed and Vance, Citation2011). In addition to inducing caspase-1-dependent IL-1β production, NLRC4 activation has also been implicated in the initiation of rapid cell death. However, studies using ASC-deficient macrophages indicate that the induction of cell death is dependent upon ASC. It is postulated, therefore, that pyroptosis, a form of programmed cell death, and caspase-1 activation are under separate control (Suzuki et al., Citation2007). Here again, additional investigations are required to elucidate the relevant molecular mechanisms.
The NLRP1 inflammasome is also considered to be involved in caspase-1 activation. By reconstituting the inflammasome biochemically, Faustin et al. (Citation2007) showed that NLRP1 forms oligomers when exposed to the bacterial protein muramyl dipeptide (MDP). It was proposed that MDP causes a conformational change in NLRP1, allowing it to bind ribonucleoside triphosphate, oligermize, and eventually to activate caspase-1. Furthermore, a separate study has revealed that the MDP binding NOD2 may facilitate these MDP conformational changes (Hsu et al., Citation2008). In that paper, it was shown that Bacillus anthracis infection induced IL-1β secretion is dependent upon NOD2. Thus, it appears that that the MDP-NOD2 complex may facilitate the activation of caspase-1 by the NLRP1 inflammasome.
The evidence available to date indicates that at least some NLR are vital to the development of skin sensitization. By sensing danger and consequently activating caspase-1, certain NLR facilitate the release of IL-1β and thus support the activation of DC. However, although NLR are important in this process, it appears that the stimulation of the NLR alone is insufficient to induce the activation of LC. It is probable that other PRR signaling pathways must act in concert with NLR for successful activation of LC during skin sensitization ().
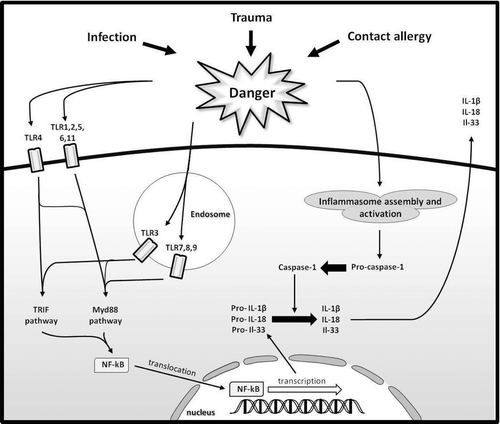
Figure 2. The interaction between the TLR and NLR signaling pathways. Upon stimulation by a variety of danger signals, different TLR activate various signaling pathways. Ligands for TLR1, -2, -5, -6, -7, -8, -9, and -10 all activate MyD88-dependent pathways, whereas ligands for TLR3 and 4 activate both MyD88 and TRIF-dependent pathways. Ultimately, activation of both the MyD88 and TRIF-dependent pathways causes the induction of NF-κB translocation. This translocation facilitates transcription of numerous pro-inflammatory proteins including pro- IL-1β, pro-IL-18, and pro-IL-33. In order for these proteins to be secreted and, thus, in order for the proteins to have physiological effect, they must be activated. This activation is achieved through cleavage by caspase-1. Caspase-1 itself must be activated, and this requires the formation of the inflammasome. The formation of the inflammasome is also induced by a number of different danger signals. Thus, it is held that activation of DC via the TLR signaling pathway requires activation of the inflammasome as well.
The additional PRR
In addition to the NLR, a whole variety of other PRR exists, including TOLL-like receptors (TLR), C-type lectin receptors (CLR), prostanoid receptors, and the neuropeptide receptors (Trinchieri and Sher, Citation2007). Many of these additional PRR are implicated in a variety of different disease processes. Significantly, many PRR have also been shown to influence the development of skin sensitization.
TOLL like receptors
As mentioned above, a family of receptors known as the TLR has been found to be capable of recognizing certain PAMP ligands. The first TLR to be identified was TLR4 (Medzhitov et al., Citation1997). In those investigations it was demonstrated that TLR4 recognizes an outer-membrane component of gram-negative bacteria; lipopolysaccharide (LPS). Subsequently, the TLR family of receptors has grown to 10 and 12 in mice and humans, respectively. As with TLR4, other TLR can be activated by a number of different PAMP ligands, including the bacterial products flagellin (TLR5) (Hayashi et al., Citation2001) and peptidoglycan (TLR2) (Schwandner et al., Citation1999). In general, the type of ligands that the TLR recognize depends upon the location of the TLR. Certain TLR (TLR1, TLR2, TLR4, TLR5, TLR6, and TLR11) are expressed on cell surfaces and, therefore, typically recognize microbial membrane products, whereas others (TLR3, TLR7, TLR8, and TLR9) are all expressed on intracellular vesicles and, therefore, typically recognize microbial nucleic acids (Kawai and Akira, Citation2010). These TLR undergo ligand induced dimerization, either forming homodimers (TLR9) or heterodimers (TLR1–TLR2, TLR2–TLR6, TLR4–TLR9, TLR3–TLR5, and TLR7–TLR9) (Ozinsky et al., Citation2000).
The importance of TLR in the development of skin sensitization was highlighted recently in an elegant study reported by Schmidt et al. (Citation2010). Here, the role of TLR4 in human nickel allergy was characterized (Schmidt et al., Citation2010). It had been appreciated for some time that humans, but not mice, were susceptible to skin sensitization to nickel without the requirement for an adjuvant. This species difference has now been resolved by the demonstration that sensitization to nickel in humans is dependent upon reaction of nickel with TLR4 (Schmidt et al., Citation2010). Specifically, histidine residues at amino acids 456 and 458 that are present in the human but not the mouse receptor are required for this interaction. Consistent with the importance of this motif, it was reported also that transgenic expression of human TLR4 in TLR4-deficient mice permitted the development of adjuvant-independent sensitization to nickel.
Stimulation of the innate immune system through activation of TLR is implicated in a variety of disease processes. Thus, an understanding of those mechanisms may serve to inform an appreciation of the influence of these receptors in skin sensitization. Involvement in such disease processes has meant that there has been a substantial investment in the characterization of TLR pathways.
The Toll IL-1 receptor (TIR) domain is found on all TLR and is integral for downstream signal transduction. Importantly, a set of TIR domain containing molecules including MyD88, TIRAP, TRAM, and TRIF can also be found intracellularly. These molecules associate with distinct TLR and subsequently activate distinct signaling pathways (Yamamoto et al., Citation2004). The TIR domain-containing adaptor MyD88 was the first member of the family to be identified. This adaptor is utilized by all TLR except TLR3 and is, thus, considered to be of pivotal importance for PAMP-dependent activation of the immune system (Medzhitov et al., Citation1998). The MyD88 dependent signaling pathway activates both NF-κB and the mitogen-activated protein kinases (MAPK). Ultimately, activation of these molecules leads to an increase in the transcription of the pro-inflammatory cytokines TNFα and pro-IL-1β amongst others (Devaraj et al., Citation2008). As discussed previously, these pro-inflammatory cytokines are important components in contact allergen-dependent activation of DC and, thus, activation of NF-κB and MAPK might therefore be important in contact allergen-dependent activation of DC. The importance of NF-κB to skin sensitization has recently been shown, again in nickel allergy. Ade et al. (Citation2007) showed that changes in the expression of activation-associated phenotypic markers were dependent upon NF-κB activation in DC exposed to NiSO4, but not in DC exposed to DNCB. This study not only provides evidence that NF-κB is important for the acquisition of sensitization to at least some contact allergens, but also supports evidence described above that indicates that nickel activates DC via interaction with the TLR4 pathway.
The pathways involved in MyD88-dependent pro-inflammatory cytokine transcription are dynamic and intricate. Upon activation of a TLR, MyD88 is recruited to the intracellular TIR domain of that TLR. MyD88 facilitates the recruitment of the IRAK1, IRAK2, IRAK4, and IRAK-M. IRAK4 is thought to be the first to be activated. The evidence available suggests that, upon activation, this kinase acts to recruit IRAK1 and IRAK2. A recent study has indicated that, because the kinase activity of IRAK1 precedes that of IRAK2 post-TLR stimulation, it is likely that IRAK1 and IRAK2 are activated sequentially (Kawagoe et al., Citation2008). Additionally, it was shown that an absence of either IRAK1 or IRAK2 has the effect of abrogating TLR-dependent cytokine production. Thus, it can be concluded that both kinases are important in the initial response to TLR activation. The IRAK-M kinase has also been investigated, with the important observation being that IRAK-M−/− mice displayed a heightened inflammatory response. This suggests that IRAK-M has an inhibitory role, possibly preventing the dissociation of IRAK1 and IRAK4 from MyD88 (Kobayashi et al., Citation2002).
Activation of the IRAK kinases ultimately results in these proteins interacting with TNF receptor associated factor (TRAF)-6 and TRAF3 (Hacker et al., Citation2006). This interaction causes TRAF6 to conjugate with the dimeric E2 ubiquitin-conjugating enzymes Uev1A and Ubc13 (Yamamoto et al., Citation2006). TRAF6 is an E3 ligase that, when conjugated to Uev1A and Ubc13, catalyses lys63 linked poly-ubiquitination (Kawai and Akira, Citation2009). By binding to the TAB1 and TAB2 components, these lys63-linked polyubiquitin chains activate the TAK1 kinase complex (Sato et al., Citation2005). Additionally, the lys63-linked polyubiquitin chains also bind to a component of the IκB kinase (IKK) complex called NEMO. On this basis it is proposed that poly-ubiquitination acts to bring the IKK and TAK1 kinase complexes together (Kawai and Akira, Citation2010). It is further proposed that the recruitment of TAK1 to IKK causes the phosphorylation and subsequent degradation of IKK. As IKK acts to inhibit the translocation of the transcription factor NF-κB in the steady state, IKK degradation will facilitate the nuclear translocation of NF-κB. After translocation into the nucleus, NF-κB promotes the transcription of a variety of genes associated with inflammation and importantly the pro-inflammatory cytokines TNFα, pro-IL-1β, IL-6, and IL-12 p40.
In addition to NF-κB, there exist a number of other transcription factors that are known to be under the control of the MyD88 pathway. One such is the activator protein 1 (AP-1) transcription factor. The AP-1 transcription factor is formed from jun-jun, jun-fos, or jun-atf dimers, and is known to control a number of cellular processes such as survival, differentiation, growth, apoptosis, cell migration, and transformation (Vesely et al., Citation2009). Although the mechanisms of AP-1 activation are not as well characterized as those of NF-κB activation, it is postulated that TAK1 is also implicated in the activation of this transcription factor. It is believed that TAK1 activates, through phosphorylation, the MAPKs that then activate AP-1 (Johnson and Lapadat, Citation2002). The evidence suggests, therefore, that the MyD88 pathway is directly responsible for the TLR dependent activation of a number of important transcription factors. In a recent study by Klekotka et al. (Citation2010), the relevance of MyD88 for skin sensitization was exemplified; that study found that mice lacking MyD88 failed to develop sensitization to DNFB. Clearly, therefore, the MyD88 pathway is required for the acquisition of skin sensitization, or at least for sensitization to DNFB.
Another important pathway induced by TLR activation is the TRIF-dependent pathway. Unlike the MyD88 pathway, this is activated only by TLR3 and TLR4 (Yamamoto et al., Citation2003). Experiments utilizing receptor-interacting protein kinase (RIP) KO mice have suggested that the adaptor molecule RIP-1 is necessary for TRIF induced NF-κB activation (Meylan et al., Citation2004). Consequently, it was hypothesized that the TRIF molecule may be recruited to the intracellular domains of TLR3 upon activation. RIP-1 may then bind to TRIF via distinct domains and thereby undergo lys63 linked poly-ubiquitination. As proposed in a recent paper, RIP-1 may bind to, and thus may be ubiquitinated by, the adaptor molecule TRADD (Pobezinskaya et al., Citation2008). This assertion is based upon the fact that, in response to TLR3 ligands, the embryonic fibroblast cells of TRADD deleted mice show reduced NF-κB activity and thus reduced TLR-dependent gene regulation.
Together with RIP-1 and TRADD, it has also been suggested that pellino-1 may have a role to play in TRIF-dependent TLR activation. Pellino-1 is a member of the Pellino family; a family of three closely-related RING-like domain-containing E3 ubiquitin ligases. In a recent study by Chang et al. (Citation2009), it was demonstrated that mice with a genetic deficiency in pellino-1 showed attenuated pro-inflammatory responses when stimulated with either TLR3 or TLR4 ligand. Interestingly, it was found that pellino-1 binds and ubiquitinates RIP-1. Thus, the proposal is that pellino-1 is an important component in the multi-protein signaling complex that binds to TRIF-1 post-TLR activation. Activation of this complex is believed to result in the activation of TAK1. As discussed above, TAK1 activation facilitates the activation of both NF-κB and MAPK signaling pathways. Taken together, it is believed that pellino-1, RIP-1, and TRADD are all integral to TRIF-dependent TLR activation.
Finally, some studies have suggested a role for TRAF6 in TRIF-dependent TLR activation. In one such study the disruption of TRAF6-binding motifs and thus the inhibition of TRAF6-TRIF binding resulted in a reduction in TRIF-induced stimulation of NF-κB (Sato et al., Citation2003). That study supports the suggestion that TRIF may bind and activate TRAF6 and, moreover, that TRIF-dependent TRAF6 activation leads to the activation of NF-κB. Using the principle of Occam’s razor, that is the explanation that makes the fewest assumptions, it is postulated that the activation of NF-κB by TRIF-dependent TRAF6 stimulation is similar to the mechanism of TRAF6 induced NF-κB activation observed in the MyD88 dependent pathway.
As well as activating NF-κB, the activation of the TRIF dependent pathway also leads to the stimulation of the regulatory factor IRF3. Although less is known about this pathway, it is thought that IRF3 activation may be induced by the IKKs TBK and IKKi (Fitzgerald et al., Citation2003). These IKKs phosphorylate IRF3 and, thus, cause the protein to translocate into the nucleus. When inside the nucleus, IRF3 binds to and activates promoters via their IRF3 binding sites. Stimulation of such promoters leads to the transcription of the cytokines RANTES and interferon (IFN)-β (Hiscott et al., Citation1999). Both of these cytokines play an important role in the immune response against viral infection. As this pathway is part of the TLR pathway and, as the TLR pathway has been implicated in certain forms of skin sensitization, one can speculate that the TRIF pathway may be important for the acquisition of sensitization. Interestingly, a recent study demonstrated that cells lacking TRAF3 fail to produce Type 1 IFN in response to TLR stimuli (Oganesyan et al., Citation2006). Moreover, this study showed that TRAF3-deficient fibroblasts were also unable to induce a Type 1 IFN response when infected directly with the vesicular stomatitis virus. Finally, the Oganesyan et al. (Citation2006) study showed that TRAF3 associates with the adaptor molecules TRIF and IRAK1 as well as with the IRF3 kinases TBK and IKKi. Taken together, these results indicate that TRAF3 has a key role to play in IRF3 activation.
It was discussed previously that induction of ROS facilitated the activation of the NLRP3 inflammasome and consequently the release of IL-1β. Intriguingly, evidence suggests that ROS may also play a role in inducing TLR-dependent pro-inflammatory gene transcription. Such studies indicate that ROS are capable of inducing the oxidative breakdown of the extracellular matrix (ECM) and, furthermore, that such breakdown of ECM releases TLR ligands (Martin et al., Citation2011). Specifically, interaction of low molecular weight hyaluronic acid (HA), a product of ECM breakdown, with TLR2 and TLR4, has recently been described. Notably, inhibition of HA using an HA inhibitor significantly abrogated the development of skin sensitization to TNCB (Martin et al., Citation2008). Additionally, there is evidence that suggests biglycan, another ECM breakdown product, is also capable of interacting with the TLR2 and TLR4 receptors (Schaefer et al., Citation2005). These findings indicate that, like HA, biglycan may have a role to play in danger signaling associated with skin sensitization. Providing further support for a role for oxidative stress in contact allergy, a recent study indicates that contact allergen-associated induction of oxidative stress, as measured by the oxidized/reduced glutathione ratio, caused the activation of certain danger signaling pathways (Mizuashi et al., Citation2005). Both DNCB and nickel induced a depletion in reduced glutathione and, furthermore, this depletion was associated with phosphorylation of MAPKs. Taken together, evidence suggests that the generation of oxidative stress plays an important role in skin sensitization.
Collectively, the evidence available currently suggests that the TLR family of PRR are capable of efficiently translating danger signals into DC activation via the induction of a range of signaling pathways. In support of this suggestion, a variety of studies have implicated TLR as a crucial PRR type in disease-associated DC activation. Pertinently, it has been shown that TLR are required for the acquisition of skin sensitization to at least some contact allergens. Specifically, it has been shown that nickel, TNCB, and DNFB all require TLR signaling pathways to induce sensitization. Moreover, this evidence indicates that TLR may also play a role in contact sensitization to other classes of contact allergen.
C-type lectin receptors
Certain C-type lectin receptors (CLR) may also be capable of modulating immune responses. Initially, these CLR were thought to bind carbohydrates in a calcium dependent manner. However, more recently, CLR that can bind carbohydrates independently of calcium have been identified. One such CLR is the PAMP receptor dectin-1; a receptor that recognizes foreign (fungal) β-1,3-glucans (Palma et al., Citation2006). Upon ligand binding, the intracellular domain of dectin-1 is phosphorylated by a src family kinase and subsequently Syk is recruited (Diebold, Citation2009). Syk acts via the signaling molecule CARD9 to activate NF-κB. In addition, Syk acts to induce the production of cytokines IL-10, IL-2, and IL-23. However, the importance of dectin-1 is still subject to debate. The results of some investigations suggest that the CLR plays a pivotal role in anti-fungal defence, whereas other papers suggest that the effect of dectin-1 is minimal (Saijo et al., Citation2007; Taylor et al., Citation2007).
In addition to dectin-1, a number of other CLR, including collectins, selectins, and the natural killer (NK) cell receptors, are implicated in the detection of danger signals. Collectins are a family of calcium-dependent, soluble CLR that have the capacity to detect pathogen-associated carbohydrates. Upon activation, these collectins initiate a variety of immune processes including agglutination, opsonization, neutralization, complement activation, and phagocytosis (Gupta and Surolia, Citation2007). Selectins also represent a group of calcium-dependent CLR involved in the detection of pathogen-associated carbohydrates. These trans-membrane molecules are believed to be intimately involved in the process of lymphocyte homing (Ley, Citation2003). Finally, the NK cell receptors have been shown to be capable of modulating the innate immune response after stimulation with the endogenous ligands MICB and MICA. As MICB and MICA are up-regulated in response to stress, it is considered that the NK cell receptors represent an important facet of the innate immune response to danger. Taken together, it appears that CLR have an important impact on danger signaling.
Although little work has been conducted to determine the role of CLR in skin sensitization, the immunological significance of these receptors is sufficient to suggest that such a role may exist. Currently, the only study that has investigated the potential role of CLR in contact allergy was by Ring et al. (Citation2009) that focussed on the roles of E- selectin and P-selectin. It was shown that administration of adenosine and regulatory T (Treg)-cells abrogated the challenge induced elicitation response to TNCB. As adenosine is known to down-regulate both E- and P-selectins, the implication is that these molecules have a role to play in the elicitation of contact allergy. However, their role in the acquisition of skin sensitization has not yet been addressed.
Prostanoids
The prostanoid receptors represent another group of PRR associated with the development of skin sensitization. The prostanoids are a group of DAMPs that include the prostaglandins (PG) D2, PGE2, PGF2α, PGI2, and thromboxane (TX) A2 (Kabashima and Miyachi, Citation2004). Production of prostanoids is complex and requires the cyclooxygenase (COX) pathway. When tissues are exposed to pathophysiological stimuli, arachidonic acid is released from the phospholipid membranes and is converted rapidly into prostanoids by the COX pathway. Once formed, these prostanoids are immediately secreted from the cell and into the extracellular milieu.
The role of PGE2 in the development of skin sensitization has been investigated recently in a study in which the PGE2 receptor EP4 was blocked using an EP4 antagonist (Yao et al., Citation2009). EP4 antagonism was shown to suppress sensitization to DNFB. Concomitantly, there was a decrease in IFNγ and IL-17 production by LN cells. The conclusion drawn was that PGE2 signaling at the EP4 receptor is important for sensitization, possibly secondary to support for the expansion of antigen-specific TH1 and TH17 cells. In support of this, the study suggested that PGE2 signaling induced production of IL-23 by DC; IL-23 is a cytokine known to be required for the expansion of TH17 cells (Yao et al., Citation2009). Given both the apparent importance of PGE2 signaling and the effect that PGE2 signaling has, it may, therefore, be worth considering the role of TH17 cells as well as TH1 cells in the development of sensitization.
Another PGE2 receptor involved in the development of skin sensitization is the EP1 receptor. Sakata et al. (Citation2008) investigated the role of this receptor by utilizing both EP1-deficient mice and an EP1 antagonist. It was found that EP1-deficient mice display a reduced TH1 cell response to DNCB. In further support of this, it was concluded that administration of an EP1 antagonist also reduced the TH1 cell activation to DNCB. In addition, DC containing inducible PGE synthase were found to increase in number in the LN. Because TH1 cell differentiation is induced in vitro by EP1 agonists, it was concluded that PGE2 secreted by DC in the LN acts to induce TH1 cell differentiation via the EP1 receptor. Another prostanoid implicated in the development of skin sensitization is PGI2. Nakajima et al. (Citation2010) examined mice lacking the PGI2 receptor (PGI2 IP) and found that DNFB failed to induce sensitization; the implication being that PGI2 IP signaling plays an important role. The same authors also demonstrated that Agonism of PGI2 IP drives the differentiation of TH1 cells. It is proposed, therefore, that PGI2 is produced by DC in the LN and serves to support the development of selective TH1 cell responses, thereby facilitating the acquisition of skin sensitization.
Neuropeptides
A number of investigations suggest that neuroimmune interactions may play important roles in the development of skin sensitization. The evidence implies that nerve fibers containing the neuropeptide calcitonin gene-related-peptide (CGRP) may act to regulate sensitization. Injection of mice with the neuropeptide depleting agent capsaicin was found to inhibit sensitization to both oxazalone and DNCB (Girolomoni and Tigelaar, Citation1990). It was subsequently demonstrated that destruction of nerve fibers prevented sensitization to DNCB (Beresford et al., Citation2004). Additionally, the administration of CGRP antagonists to mice suppressed sensitization to the experimental allergen fluorescein isothiocyanate (FITC). This suppression was associated with reduced DC migration and maturation (Maruyama et al., Citation2007).
Thus, on the basis of the evidence summarized above, neuropeptides, and in particular CGRP, are important in the development of sensitization. However, contradictory studies exist, including those in which treatment with CGRP was found to inhibit sensitization to TNCB and DNFB in mice (Asahina et al., Citation1995). In an attempt to clarify the role of CGRP in skin sensitization, Mikami et al. (Citation2011) depleted the CGRP receptor RAMP-1 in mice and examined development of sensitization to FITC and DNCB. RAMP-1 deficient mice were found to have developed stronger sensitization to FITC, but a weaker response to TNCB. In this context, it is relevant that sensitization to FITC is associated with selective TH2-type responses. One interpretation is, therefore, that CGRP serves to promote TH2 responses while inhibiting the development of TH1 responses. Consistent with this is the fact that CGRP promotes the production of IL-4 (Mikami et al., Citation2011).
It appears, therefore, that CGRP modulates cutaneous immune responses due to its influence on DC and T-cell function. Although initial experiments were reported to show that depletion of neuropeptides including CGRP attenuated sensitization to DNCB, it may be that this is not entirely attributable to CGRP. The suggestion is that depletion in the levels of other neuropeptides such as substance P may be responsible for the inhibition of skin sensitization. Consistent with this, it has been found that mice lacking neutral endopeptidase (NEP) display a stronger response to DNCB (Scholzen et al., Citation2001). As NEP degrades substance P, it is considered that the latter may be an important promoter of skin sensitization.
Nrf2-Keap1-ARE toxicity pathway
In addition to the PRR pathways, it is believed that there may be other pathways with important roles to play in the modulation of the immune response to skin sensitizers. One such pathway is the Nrf2-Keap1-ARE toxicity pathway. This pathway is responsible for detecting electrophilic stress and driving an appropriate response. In this role, stress induced by electrophilic chemicals causes the dissociation of Keap1 from Nrf2. Sequentially, Keap1 then accumulates in the nucleus and induces the transcription of ARE-dependent genes. These genes code for a variety of proteins including phase II detoxifying enzymes and the pro-inflammatory chemokine IL-8. Importantly, it has been shown that many contact allergens interact with the Nrf2-Keap1-ARE toxicity pathway (Natsch et al., Citation2008; Ade et al., Citation2007). The view is, therefore, that interaction of contact allergens with the Nrf2-Keap1-ARE pathway may be an important component of signaling and inflammation in skin sensitization.
Linking danger signals and contact allergy
Given the apparent importance of danger signal-induced DC activation in the generation of skin sensitization, very little emphasis has been placed on elucidating how contact allergens induce danger in the skin. This review postulates that different chemical allergens induce danger-like responses in DC via one or more of several different mechanisms. However, the routes by which contact allergens may elicit ‘danger’ and induce caspase-1 activation can be divided into two distinct groups ().
Figure 3. Linking danger signals and contact sensitization. The current review suggests that contact allergens induce the provision of danger signals via two routes, i.e., either directly or indirectly. An example of direct provision is observed in nickel sensitization. Here, nickel acts directly to bind TLR4 and activate the TLR4 pathway. Alternatively, an example of indirect provision is observed in sensitization to TNCB. In this, TNCB induces the production of ROS. These ROS cause the breakdown of long hyaluronic acid (HA) fragments into short HA fragments. The short HA fragments are capable of activating TLR2 and TLR4, thus it is held that TNCB activates DC via this mechanism.
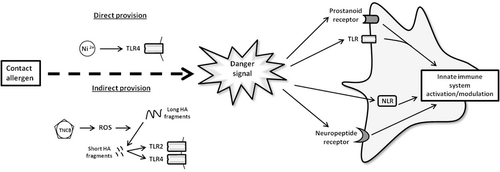
First, it is proposed that some contact allergens may evoke ‘danger’ by directly stimulating PRR. An example discussed previously is provided by nickel, as this interacts directly with TLR4 for the successful initiation of skin sensitization (Schmidt et al., Citation2010). Second, it is proposed that certain contact allergens may act indirectly; inducing cellular changes that consequently cause the elicitation of ‘danger’. An example here is the generation of ROS by TNCB and subsequent degradation of the ECM to provide relevant signals. Providing such links between the elicitation of danger signals and the acquisition of sensitization provide new insights into the factors that confer allergenic potential on certain chemicals and of the variables that influence skin sensitizing potency.
Concluding remarks
The question of what makes a chemical an allergen is a fascinating one. The importance of this conundrum is currently assuming greater importance due to the aspiration of finding suitable in vitro assays to identify and characterize skin-sensitizing chemicals. This is a daunting objective, since, although hazard identification using non-animal methods appears to be relatively tractable, assessment of skin sensitizing potency for the purposes of risk assessment remains a very substantial challenge. To meet this challenge it will be necessary to exploit a rapidly increasing understanding of the factors that influence the acquisition of skin sensitization. In this context an awareness of the various danger signals that facilitate the initiation of cutaneous immune responses to chemical allergens is of considerable importance, as is an appreciation of the ways in which the innate and adaptive immune systems communicate and interact in orchestrating the function of DC. At a pragmatic level, and with respect to the further development and refinement of methods for hazard identification and characterization, it will be appropriate to consider not only exposure to allergens, but also the role that innate signals will play in the responsiveness of DC.
Declaration of interest
The authors report no conflicts of interest. The authors alone are responsible for the content and writing of the paper.
References
- Ade, N., Antonios, D., Kerdine-Romer, S., Boisleve, F., Rousset, F., Pallardy, M. 2007. NF-κB plays a major role in the maturation of human dendritic cells induced by NiSO4 but not by DNCB. Toxicol. Sci.99:488–501.
- Antonopoulos, H., Cumberbatch, M., Dearman, R. J., Daniel, R. J., Kimber, I., Groves, R. W. 2001. Functional caspase-1 is required for Langerhans cell migration and optimal contact sensitization in mice. J. Immunol.166:3672–3677.
- Arthur, J. C., Lich, J. D., Ye, Z., Allen, I. C., Gris, D., Wilson, J. E., Schneider, M., Roney, K. E., O’Connor, B. P., Moore, C. B., Morrison, A., Sutterwala, F. S., Bertin, J., Koller, B. H., Liu, Z., Ting, J. P. 2010. Cutting Edge: NLRP12 controls dendritic and myeloid cell migration to affect contact hypersensitivity. J. Immunol. 185:4515–4519.
- Asahina, A., Hosoi, J., Beissert, S., Stratigos, A., Granstein, R. D. 1995. Inhibition of the induction of delayed-type and contact hypersensitivity by calcitonin-gene-related peptide. J. Immunol. 154:3056–3061.
- Beresford, L., Orange, O., Bell, E. B., Miyan, J. A. 2004. Nerve fibres are required to evoke a contact sensitivity response in mice. Immunology 111:118–125.
- Bruchhausen, S., Zahn, S., Valk, E., Knop, J., Becker, D. 2003. Thiol anti-oxidants block the activation of antigen-presenting cells by contact sensitizers. J. Invest. Dermatol. 121:1039–1044.
- Chang, M., Jin, W., Sun, S. C. 2009. Peli1 facilitates TRIF-dependent Toll-like receptor signaling and pro-inflammatory cytokine production. Nat. Immunol. 10:1089–1095.
- Cumberbatch, M., Clelland, K., Dearman, R. J., Kimber, I. 2005. Impact of cutaneous IL-10 on resident epidermal Langerhans’ cells and the development of polarized immune responses. J. Immunol.175:43–50.
- Cumberbatch, M., Scott, R. C., Basketter, D. A., Scholes, E. W., Hilton, J., Dearman, R. J., Kimber, I. 1993. Influence of sodium lauryl sulfate on 2,4-dinitrochlorobenzene-induced lymph-node activation. Toxicology 77:181–191.
- Devaraj, S., Dau, M. R., Rockwood, J., Winter, W., Griffen, S. C., Jialal, I. 2008. Increased toll-like receptor (TLR)-2 and TLR-4 expression in monocytes from patients with Type 1 diabetes: Further evidence of a pro-inflammatory state. J. Clin. Endocrinol. Metab. 93:578–583.
- Diebold, S. S. 2009. Activation of dendritic cells by toll-like receptors and C-type lectins. Handbook Exp. Pharmacol. 188:3–30.
- Dostert, C., Petrilli, V., van Bruggen, R., Steele, C., Mossman, B. T., Tschopp, J. 2008. Innate immune activation through Nalp3 inflammasome sensing of asbestos and silica. Science 320:674–677.
- Enk, A. H., Katz, S. I. 1992. Early molecular events in the induction phase of contact sensitivity. Proc. Natl. Acad. Sci. USA 89:1398–1402.
- Faustin, B., Lartigue, L., Bruey, J. M., Luciano, F., Sergienko, E., Bailly-Maitre, B., Volkmann, N., Hanein, D., Rouiller, I., Reed, J. C. 2007. Reconstituted NALP1 inflammasome reveals two-step mechanism of caspase-1 activation. Mol. Cell 25:713–724.
- Fitzgerald, K. A., McWhirter, S. M., Faia, K. L., Rowe, D. C., Latz, E., Golenbock, D. T., Coyle, A. J., Liao, S. M., Maniatis, T. 2003. IKKϵ and TBK1 are essential components of the IRF3 signaling pathway. Nat. Immunol. 4:491–496.
- Franchi, L., Eigenbrod, T., Munoz-Planillo, R., Nunez, G. 2009. The inflammasome: A caspase-1-activation platform that regulates immune responses and disease pathogenesis. Nat. Immunol. 10:241–247.
- Gao, Z., Tseng, C. H., Strober, B. E., Pei, Z., Blaser, M. J. 2008. Substantial alterations of the cutaneous bacterial biota in psoriatic lesions. Plos One 3(7): e2719.
- Girolomoni, G., Tigelaar, R. E. 1990. Capsaicin-sensitive primary sensory neurons are potent modulators of murine delayed-type hypersentitivity reactions. J. Immunol.145:1105–1112.
- Grabbe, S., Steinert, M., Mahnke, K., Schwarz, A., Luger, T. A., Schwarz, T. 1996. Dissection of antigenic and irritative effects of epicutaneously applied haptens in mice. Evidence that not the antigenic component but non-specific proinflammatory effects of haptens determine the concentration-dependent elicitation of allergic contact dermatitis. J. Clin. Invest. 98:1158–1164.
- Gupta, G., Surolia, A. 2007. Collectins: Sentinels of innate immunity. Bioessays 29:452–464.
- Hacker, H., Redecke, V., Blagoev, B., Kratchmarova, I., Hsu, L. C., Wang, G. G., Kamps, M. P., Raz, E., Wagner, H., Hacker, G., Mann, M., Karin, M. 2006. Specificity in Toll-like receptor signalling through distinct effector functions of TRAF3 and TRAF6. Nature 439:204–207.
- Hayashi, F., Smith, K. D., Ozinsky, A., Hawn, T. R., Yi, E. C., Goodlett, D. R., Eng, J. K., Akira, S., Underhill, D. M., Aderem, A. 2001. The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nature 410:1099–1103.
- Hewinson, J., Moore, S. F., Glover, C., Watts, A. G., Mackenzie, A. B. 2008. A key role for redox signaling in rapid P2X7 receptor-induced IL-1# processing in human monocytes. J. Immunol. 180:8410–8420.
- Hiscott, J., Pitha, P., Genin, P., Nguyen, H., Heylbroeck, C., Mamane, Y., Algarte, M., Lin, R. T. 1999. Triggering the interferon response: The role of IRF-3 transcription factor. J. Interferon Cytokine Res. 19:1–13.
- Hornung, V., Bauernfeind, F., Halle, A., Samstad, E. O., Kono, H., Rock, K. L., Fitzgerald, K. A., Latz, E. 2008. Silica crystals and aluminum salts activate the NALP3 inflammasome through phagosomal destabilization. Nat. Immunol. 9:847–856.
- Hsu, L. C., Ali, S. R., McGillivray, S., Tseng, P. H., Mariathasan, S., Humke, E. W., Eckmann, L., Powell, J. J., Nizet, V., Dixit, V. M., Karin, M. 2008. A NOD2-NALP1 complex mediates caspase-1-dependent IL-1# beta secretion in response to Bacillus anthracis infection and muramyl dipeptide. Proc. Natl. Acad. Sci. USA 105:7803–7808.
- Igyarto, B. Z., Haley, K., Ortner, D., Bobr, A., Gerami-Nejad, M., Edelson, B. T., Zurawski, S. M., Malissen, B., Zurawski, G., Berman, J., Kaplan, D. H. 2011. Skin-resident murine dendritic cell subsets promote distinct and opposing antigen-specific T-helper cell responses. Immunity 35:260–272.
- Johnson, G. L., Lapadat, R. 2002. Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases. Science 298:1911–1912.
- Kabashima, K., Miyachi, Y. 2004. Prostanoids in the cutaneous immune response. J. Dermatol. Sci. 34:177–184.
- Kanneganti, T. D. , Lamkanfi, M., Kim, Y. G., Chen, G., Park, J. H., Franchi, L., Vandenabeele, P., Nunez, G. 2007. Pannexin-1-mediated recognition of bacterial molecules activates the cryopyrin inflammasome independent of Toll-like receptor signaling. Immunity 26:433–443.
- Kaplan, D. H. 2010. In vivo function of Langerhans cells and dermal dendritic cells. Trends Immunol. 31:446–451.
- Kawagoe, T., Sato, S., Matsushita, K., Kato, H., Matsui, K., Kumagai, Y., Saitoh, T., Kawai, T., Takeuchi, O., Akira, S. 2008. Sequential control of Toll-like receptor-dependent responses by IRAK1 and IRAK2. Nat. Immunol. 9:684–691.
- Kawai, T., Akira, S. 2009. The roles of TLRs, RLRs and NLRs in pathogen recognition. Int. Immunol. 21:317–337.
- Kawai, T., Akira, S. 2010. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 11:373–384.
- Kehren, J., Desvignes, C., Krasteva, M., Ducluzeau, M. T., Assossou, O., Horand, F., Hahne, M., Kagi, D., Kaiserlian, D., Nicolas, J. F. 1999. Cytotoxicity is mandatory for CD8+ T-cell-mediated contact hypersensitivity. J. Exp. Med.189:779–786.
- Khakh, B. S., North, R. A. 2006. P2X receptors as cell-surface ATP sensors in health and disease. Nature 442:527–532.
- Kimber, I., Basketter, D. A., Gerberick, G. F., Dearman, R. J. 2002. Allergic contact dermatitis. Int. Immunopharmacol. 2:201–211.
- Kimber, I., Basketter, D. A., Gerberick, G. F., Ryan, C. A., Dearman, R. J. 2011. Chemical allergy: Translating biology into hazard characterization. Toxicol. Sci. 120(Suppl 1):S238–S268.
- Klekotka, P. A., Yang, L., Yokoyama, W. M. 2010. Contrasting roles of the IL-1 and IL-18 receptors in MyD88-dependent contact hypersensitivity. J. Invest. Dermatol. 130:184–191.
- Kobayashi, K., Hernandez, L. D., Galan, J. E., Janeway, C. A., Medzhitov, R., Flavell, R. A. 2002. IRAK-M is a negative regulator of toll-like receptor signaling. Cell 110:191–202.
- Kofoed, E. M., Vance, R. E. 2011. Innate immune recognition of bacterial ligands by NAIPs determines inflammasome specificity. Nature 477:592–595.
- Ley, K. 2003. The role of selectins in inflammation and disease. Trends Mol. Med. 9:263–268.
- Martin, S. F., Dudda, J. C., Bachtanian, E., Lembo, A., Liller, S., Duerr, C., Heimesaat, M. M., Bereswill, S., Fejer, G., Vassileva, R., Jakob, T., Freudenberg, N., Termeer, C. C., Johner, C., Galanos, C., Freudenberg, M. A. 2008. Toll-like receptor and IL-12 signaling control susceptibility to contact hypersensitivity. J. Exp. Med. 205:2151–2162.
- Martin, S. F., Esser, P. R., Weber, F. C., Jakob, T., Freudenberg, M. A., Schmidt, M., Goebeler, M. 2011. Mechanisms of chemical-induced innate immunity in allergic contact dermatitis. Allergy 66:1152–1163.
- Martinon, F. 2010. Signaling by ROS drives inflammasome activation. Eur. J. Immunol. 40:616–619
- Maruyama, T., Iizuka, H., Tobisawa, Y., Shiba, T., Matsuda, T., Kurohane, K., Imai, Y. 2007. Influence of local treatments with capsaicin or allyl isothiocyanate in the sensitization phase of a fluorescein-isothiocyanate-induced contact sensitivity model. Int. Arch. Allergy Immunol. 143:144–154.
- Matzinger, P. 1994. Tolerance, danger, and the extended family. Ann. Rev. Immunol. 12:991–1045.
- Medzhitov, R., Preston-Hurlburt, P., Kopp, E., Stadlen, A., Chen, C. Q., Ghosh, S., Janeway, C. A. 1998. MyD88 is an adaptor protein in the hToll/IL-1 receptor family signaling pathways. Mol. Cell 2:253–258.
- Medzhitov, R., Preston-Hurlburt, P., Janeway, C. A. 1997. A human homologue of the Drosophila Toll protein signals activation of adaptive immunity. Nature 388:394–397.
- Mehrotra, P., Mishra, K. P., Raman, G., Banerjee, G. 2005. Differential regulation of free radicals (reactive oxygen and nitrogen species) by contact allergens and irritants in human keratinocyte cell line. Toxicol. Mech. Meth. 15:343–350.
- Merad, M., Manz, M. G., Karsunky, H., Wagers, A., Peters, W., Charo, I., Weissman, I. L., Cyster, J. G., Engleman, E. G. 2002. Langerhans cells renew in the skin throughout life under steady-state conditions. Nat. Immunol. 3:1135–1141.
- Meylan, E., Burns, K., Hofmann, K., Blancheteau, V., Martinon, F., Kelliher, M., Tschopp, J. 2004. RIP1 is an essential mediator of Toll-like receptor 3-induced NF-κB activation. Nat. Immunol. 5:503–507.
- Mikami, N., Matsushita, H., Kato, T., Kawasaki, R., Sawazaki, T., Kishimoto, T., Ogitani, Y., Watanabe, K., Miyagi, Y., Sueda, K., Fukada, S. I., Yamamoto, H., Tsujikawa, K. 2011. Calcitonin gene-related peptide is an important regulator of cutaneous immunity: Effect on dendritic cell and T-cell functions. J. Immunol. 186:6886–6893.
- Mizuashi, M., Ohtani, T., Nakagawa, S., Aiba, S. 2005. Redox imbalance induced by contact sensitizers triggers the maturation of dendritic cells. J. Invest. Dermatol. 124:579–586.
- Nakajima, S., Honda, T., Sakata, D., Egawa, G., Tanizaki, H., Otsuka, A., Moniaga, C. S., Watanabe, T., Miyachi, Y., Narumiya, S., Kabashima, K. 2010. Prostaglandin I2-IP signaling promotes TH1 Differentiation in a mouse model of contact hypersensitivity. J. Immunol. 184:5595–5603.
- Natsch, A., Emter, R., Ellis, G. 2008. Skin sensitizers induce antioxidant response element dependent genes: Application to the in vitro testing of the sensitization potential of chemicals. Toxicol. Lett. 180:S101.
- Oganesyan, G., Saha, S. K., Guo, B. C., He, J. Q., Shahangian, A., Zarnegar, B., Pery, A., Cheng, G. H. 2006. Critical role of TRAF3 in the Toll-like receptor-dependent and -independent anti-viral response. Nature 439:208–211.
- Ogura, Y., Sutterwala, F. S., Flavell, R. A. 2006. The inflammasome: First line of the immune response to cell stress. Cell 126:659–662.
- Ozinsky, A., Underhill, D. M., Fontenot, J. D., Hajjar, A. M., Smith, K. D., Wilson, C. B., Schroeder, L., Aderem, A. 2000. The repertoire for pattern recognition of pathogens by the innate immune system is defined by cooperation between Toll-like receptors. Proc. Natl. Acad. Sci. USA 97:13766–13771.
- Palma, A. S., Feizi, T., Zhang, Y. B., Stoll, M. S., Lawson, A. M., Diaz-Rodriguez, E., Campanero-Rhodes, M. A., Costa, J., Gordon, S., Brown, G. D., Chai, W. G. 2006. Ligands for the β-glucan receptor, Dectin-1, assigned using “designer” microarrays of oligosaccharide probes (neoglycolipids) generated from glucan polysaccharides. J. Biol. Chem. 281:5771–5779.
- Pedra, J. H. F., Cassel, S. L., Sutterwala, F. S. 2009. Sensing pathogens and danger signals by the inflammasome. Curr. Opin. Immunol. 21:10–16.
- Petrilli, V., Papin, S., Dostert, C., Mayor, A., Martinon, F., Tschopp, J. 2007. Activation of the NALP3 inflammasome is triggered by low intracellular potassium concentration. Cell Death Differen. 14:1583–1589.
- Pobezinskaya, Y. L., Kim, Y. S., Choksi, S., Morgan, M. J., Li, T., Liu, C., Liu, Z. 2008. The function of TRADD in signaling through tumor necrosis factor receptor 1 and TRIF-dependent Toll-like receptors. Nat. Immunol. 9:1047–1054.
- Ring, S., Oliver, S. J., Cronstein, B. N., Enk, A. H., Mahnke, K. 2009. CD4+CD25+ regulatory T-cells suppress contact hypersensitivity reactions through a CD39, adenosine-dependent mechanism. J. Allergy Clin. Immunol. 123:1287–1296.
- Saijo, S., Fujikado, N., Furuta, T., Chung, S. H., Kotaki, H., Seki, K., Sudo, K., Akira, S., Adachi, Y., Ohno, N., Kinjo, T., Nakamura, K., Kawakami, K., Iwakura, Y. 2007. Dectin-1 is required for host defense against Pneumocystis carinii but not against Candida albicans. Nat. Immunol. 8:39–46.
- Saint-Mezard, P., Rosieres, A., Krasteva, M., Berard, F., Dubois, B., Kaiserlian, D., Nicolas, J. F. 2004. Allergic contact dermatitis. Eur. J. Dermatol. 14:284–295.
- Sakata, D., Nagamachi, M., Kabashima, K., Furuyashiki, T., Soontrapa, K., Matsuoka, T., Segi-Nishida, E., Narumiya, S. 2008. Facilitation of TH1-mediated immune response by prostaglandin E receptor EP1. J Exp Med. 204(12):2865–2874.
- Sato, S., Sanjo, H., Takeda, K., Ninomiya-Tsuji, J., Yamamoto, M., Kawai, T., matsumoto, K., Takeuchi, O., Akira, S. 2005. Essential function for the kinase TAK1 in innate and adaptive immune responses. Nat. Immunol. 6:1087–1095.
- Sato, S., Sugiyama, M., Yamamoto, M., Watanabe, Y., Kawai, T., Takeda, K., Akira, S. 2003. Toll/IL-1 receptor domain-containing adaptor inducing IFNβ(TRIF) associates with TNF receptor-associated factor-6 and TANK-Binding kinase 1, and activates two distinct transcription factors, NF-κB and IFN-regulatory factor-3, in the toll-like receptor signaling. J. Immunol. 171:4304–4310.
- Schaefer, L., Babelova, A., Kiss, E., Hausser, H. J., Baliova, M., Krzyzankova, M., Marsche, G., Young, M. F., Mihalik, D., Gotte, M., Malle, E., Schaefer, R. M., Grone, H. J. 2005. The matrix component biglycan is proinflammatory and signals through Toll-like receptors 4 and 2 in macrophages. J. Clin. Invest. 115:2223–2233.
- Schmidt, M., Raghavan, B., Mueller, V., Vogl, T., Fejer, G., Tchaptchet, S., Keck, S., Kalis, C., Nielsen, P. J., Galanos, C., Roth, J., Skerra, A., Martin, S. F., Freudenberg, M. A., Goebller, M. 2010. Crucial role for human Toll-like receptor-4 in the development of contact allergy to nickel. Nat. Immunol. 11:814–819.
- Scholzen, T. E., Steinhoff, M., Bonaccorsi, P., Klein, R., Amadesi, S., Geppetti, P., Lu, B., Gerard, N. P., Olerud, J. E., Luger, T. A., Bunnett, N. W., Grady, E. F., Armstrong, C. A., Ansel, J. C. 2001. Neutral endopeptidase terminates substance P-induced inflammation in allergic contact dermatitis. J. Immunol.166:1285–1291.
- Schwandner, R., Dziarski, R., Wesche, H., Rothe, M., Kirschning, C. J. 1999. Peptidoglycan and lipoteichoic acid-induced cell activation is mediated by toll-like receptor-2. J. Biol. Chem. 274:17406–17409.
- Shaw, P. J., Lamkanfi, M., Kanneganti, T. D. 2010. NOD-like receptor (NLR) signaling beyond the inflammasome. Eur. J. Immunol. 40:624–627.
- Suzuki, T., Franchi, L., Toma, C., Ashida, H., Ogawa, M., Yoshikawa, Y., Mimuro, H., Inohara, N., Sasakawa, C., Nunez, G. 2007. Differential regulation of caspase-1 activation, pyroptosis, and autophagy via Ipaf and ASC in Shigella-infected macrophages. Plos Pathogens 3:1082–1091.
- Taylor, P. R., Tsoni, S. V., Willment, J. A., Dennehy, K. M., Rosas, M., Findon, H., Haynes, K., Steele, C., Botto, M., Gordon, S., Brown, G. D. 2007. Dectin-1 is required for #glucan recognition and control of fungal infection. Nat. Immunol. 8:31–38.
- Thyssen, J. P., Linneberg, A., Menne, T., Johansen, J. D. 2007. The epidemiology of contact allergy in the general population – prevalence and main findings. Contact Dermatitis 57:287–299.
- Trinchieri, G., Sher, A. 2007. Cooperation of Toll-like receptor signals in innate immune defense. Nat. Rev. Immunol. 7:179–190.
- Vesely, P. W., Staber, P. B., Hoefler, G., Kenner, L. 2009. Translational regulation mechanisms of AP-1 proteins. Mutat. Res. 682:7–12.
- Watanabe, H., Gaide, O., Petrilli, V., Martinon, F., Contassot, E., Roques, S., Kummer, J. A., Tschopp, J., French, L. E. 2007. Activation of the IL-1β-processing inflammasome is involved in contact hypersensitivity. J. Invest. Dermatol. 127:1956–1963.
- Weber, F. C., Esser, P. R., Mueller, T., Ganesan, J., Pellegatti, P., Simon, M. M., Zeiser, R., Idzko, M., Jakob, T., Martin, S. F. 2010. Lack of the purinergic receptor P2X(7) results in resistance to contact hypersensitivity. J. Exp. Med. 207:2609–2619.
- Yamamoto, M., Okamoto, T., Takeda, K., Sato, S., Sanjo, H., Uematsu, S., Saitoh, T., Yamamoto, N., Sakurai, H., Ishii, K. J., Yamaoka, S., Kawai, T., Matsuura, Y., Takeuchi, O., Akira, S. 2006. Key function for the Ubc13 E2 ubiquitin-conjugating enzyme in immune receptor signaling. Nat. Immunol. 7:962–970.
- Yamamoto, M., Sato, S., Hemmi, H., Hoshino, K., Kaisho, T., Sanjo, H., Takeuchi, O., Sugiyama, M., Okabe, M., Takeda, K., Akira, S. 2003. Role of adaptor TRIF in the MyD88-independent toll-like receptor signaling pathway. Science 301:640–643.
- Yamamoto, M., Takeda, K., Akira, S. 2004. TIR domain-containing adaptors define the specificity of TLR signaling. Mol. Immunol. 40:861–868.
- Yao, C., Sakata, D., Esaki, Y., Li, Y., Matsuoka, T., Kuroiwa, K., Sugimoto, Y., Narumiya, S. 2009. Prostaglandin E2-EP4 signaling promotes immune inflammation through TH1 cell differentiation and TH17 cell expansion. Nat. Med. 15:633–640.
- Zhao, Y., Yang, J., Shi, J., Gong, Y. N., Lu, Q., Xu, H., Liu, L., Shao, F. 2011. The NLRC4 inflammasome receptors for bacterial flagellin and Type III secretion apparatus. Nature 477:596–600.
- Zhou, R., Yazdi, A. S., Menu, P., Tschopp, J. 2011. A role for mitochondria in NLRP3 inflammasome activation. Nature 469:221.