
Abstract
The present study is focused on evaluating the antimutagenic properties of Schiff bases and Mn(III) complexes with L-Threonine, L-Serine and L-Tyrosine, which have antimicrobial activity. These six compounds were investigated for antimutagenic properties against Aflatoxin Bı (AFBı) by the micronucleus (MN) assay in a human lymphocyte cell culture in vitro. The protective role of these compounds against AFBı-induced MN is probably related to its doses. A mechanism has been proposed to reduce the effect of AFBı.
Introduction
Many amino acid derivatives possess a wide variety of pharmacological activities (CitationKuhl et al. 2005). Amino acids hold promise as ideal targets for tumor imaging. They are necessary for the continuous, sustained, and uncontrolled growth of tumor cells. Studies have demonstrated that malignant tumors can be detected with high sensitivity and specificity, by imaging their increased metabolic rates for amino acids. Therefore, many natural and artificial amino acids have been radiolabeled for positron emission tomography (PET) imaging of tumors (CitationMcConathy et al. 2002).
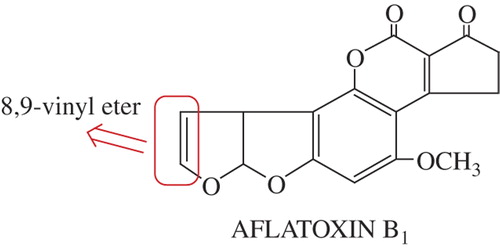
Recently, several studies have demonstrated that certain amino acids (such as cysteine, glycine, tryptophan, lysine, arginine, glutamine, and alanine) show an antimutagenic effect, tested by different systems (CitationRoy et al. 2002, CitationTavares et al. 1998, and CitationHandique and Aprem 1997). However, no report so far has demonstrated the protective effect of these compounds against AFBı genotoxicity. AFBı is the most potent of the naturally occurring mycotoxins. The activation of AFB1 on organisms, initiated by the oxidation of the 8,9-vinyl ether bond, is shown in . AFB1 in itself is not dangerous, but is metabolized by the body, ultimately producing a carcinogenic metabolite. Oxidized AFB1 may bind to nuclear DNA, resulting in nuclear damage due to electrophilic attack on the N7 position of guanine of DNA and RNA (CitationMclean and Dutton 1995).
Molecules attached with amino acids have wide applications for the synthesis of drugs as antimutagenic and antibacterial agents. Therefore, there is considerable interest in the synthesis and characterization of these compounds. However, drug resistance against antimutagenic/antibacterial agents may pose a problem in their use for medical purposes. The problem could be overcome by the preparation of metal complexes by a process of chelation, with the coordination of transition metal ions. It is well known that N and O atoms play a key role in the coordination of metals. CitationLiping et al. have studied the inhibitory activity of oxovanadium (IV) complexes with amino acid-Schiff base, against human tyrosine phosphatase 1B in vitro (2011). Zasukhina et al., have studied the antimutagenic activity of compounds including nitrogen (2003). Many research studies have demonstrated that the genotoxicity of some metal salts (e.g. Cd(II), Ni(II), Pt(II), etc.) might depend on the phase of the cell cycle in human lymphocytes (CitationHartmann and Hartwig 1998, CitationSnow 1992, CitationColuccia et al. 1984).
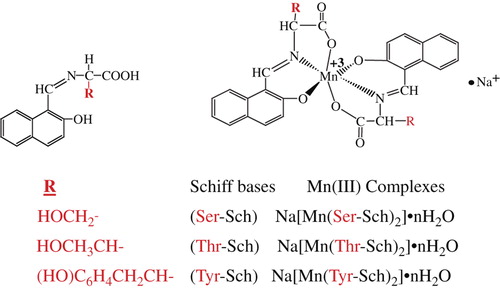
We know that one of the major challenges in medical and drug delivery is the development of new antimutagens, because they have good prospects of practical use in preventing delayed negative effects of mutagens induced in humans, the most important of which are the high prevalence of hereditary diseases and cancer (CitationZasukhina et al. 2003). Therefore, the purpose of this study was to investigate the antimutagenic effects of Schiff bases attached with L-Serine and L-Threonine, and L-Tyrosine, and their Mn(III) complexes, on human peripheral blood in vitro, by the micronucleus assay. Furthermore, antimicrobial activity was tested in the ligands and complexes by the well-diffusion method. First of all, amino acid-Schiff bases were synthesized by using the condensation methods. Then, their Mn(III) complexes were synthesized by means of a template method ().
Materials and methods
Chemicals and physical measurements
All chemicals investigated in the study were of reagent grade and were purified when necessary. All organic solvents used in this study were purified according to standard methods. The amino acids (L-Threonine, L-Serine and L-Tyrosine), 2-hydroxy-1-naphthaldehyde, methanol, and n-heptane were purchased from Sigma-Aldrich. Manganese (III) acetate was prepared by the method used by Gunduz et al. (CitationLandini et al. 2004). Elemental analyses were performed with a LECO-CHNS-9320 instrument. The metal contents were determined using a Philips PU 9285 atomic absorption instrument. 1H-NMR spectra were recorded with a Bruker DPX-300 MHz and 100 MHz, using TMS as an internal standard and CDCl3 as solvent. The electronic spectra were recorded on a UV-1800 ENG240V spectrophotometer in ethanol. The IR spectra were recorded on a Mattson-5000 FTIR instrument in KBr pellets. The melting points were determined with a Barnstead-Electrothermal-9200 melting point apparatus. Magnetic measurements were performed with a Sherwood Scientific Magnetic Susceptibility Balance (Model No: MK 1) at 21 °C with Hg[Co(NCS)4] as a calibration.
Test microorganisms and medium
The bacterial subcultures chosen were Listeria monocytogenes 4b ATCC19115, Staphylococcus aureus ATCC25923, Escherichia coli ATCC1280, Salmonella typhi H NCTC901.8394, Brucella abortus RSKK03026, Staphylococcus epidermis sp., Micrococcus luteus ATCC9341, Shigella dysenteriae type 7 NCTC 9363, Pseudomonas putida sp., and Bacillus cereus RSKK863. An antifungal susceptibility test was performed using Candida albicans Y-1200-NIH, Tokyo.
Synthesis
Schiff Bases: A solution of 2-hydroxy-1-naphthaldehyde (5 mmol, 0.61 g) in methanol (50 ml) was added to an amino acid (L-Threonine, L-Serine and L-Tyrosine, 5 mmol)- MeOH solution (50 ml), and the synthesis was carried out according to the method we had detailed in an earlier article (CitationGunduz et al. 1994). Mn(III) complexes (template method): Mn(III) complexes were prepared by the template method. Firstly, Mn(CH3COO)3.2H2O was synthesized according to the method explained in our earlier article. Then, the amino acids (L-Glutamine and L-Aspartic acid), 2-hydroxy-1-naphthaldehyde, and Mn(CH3COO)3 were dissolved in methanol, and the synthesis was carried out according to the procedure (CitationGunduz et al. 1994).
Detection of antimicrobial activity
The ligands and complexes were tested for their antimicrobial activity by the well-diffusion method. Each ligand and complex was kept dry at room temperature and dissolved (10 − 3 M) in DMF. DMF was used as a solvent and also for control. It was found to have no antimicrobial activity against any of the organisms tested. For the study, 1% (v/v) of a 24-h broth culture containing 106 CFU/ml was placed in sterile Petri dishes. Mueller- Hinton Agar (MHA) (15 ml) kept at 45°C was then poured into the Petri dishes and allowed to solidify. Then, wells measuring 6 mm in diameter were punched carefully using a sterile cork borer and were entirely filled with the test solutions. The plates were incubated for 24 h at 37°C. On completion of the incubation period, the mean value obtained for the two holes was used to calculate the zone of growth inhibition of each sample. Bacterial subcultures and yeast were tested for resistance to five antibiotics (produced by Oxoid Ltd., Basingstoke, UK): ampicillin (preventing the growth of gram-negative bacteria), nystatin (binding to sterols in the fungal cellular membrane, altering the permeability, and allowing leakage of the cellular contents), kanamycin (used in molecular biology as an agent to isolate bacteria), sulphamethoxazole (a bacteriostatic antibacterial agent that interferes with folic acid synthesis in susceptible bacteria), amoxicillin (a b-lactam antibiotic used to treat bacterial infections caused by susceptible microorganisms).
Cytogenetic analysis
Peripheral blood lymphocytes were taken from four non-smoking healthy individuals (two men and two women). Lymphocyte cultures were set up by adding 0.5 mL of heparinized whole blood to RPMI-1640 chromosome medium supplemented with 15% heat-inactivated fetal calf serum, 100 IU/mL of streptomycin, 100 IU/mL of penicillin, and 1% L-glutamine. Lymphocytes were stimulated to divide by 1% phytohemagglutinin.
The experiments were performed on 26 groups as follows:
Group 1: Control
Group 2: 5 μM AFB1; 0
Group 3: Na[Mn(Ser-Sch)2] 40 μM
Group 4: 5 μM AFB1 + Na[Mn(Ser-Sch)2] (5 μg/ml)
Group 5: 5 μM AFB1 + Na[Mn(Ser-Sch)2] (10 μg/ml)
Group 6: 5 μM AFB1 + Na[Mn(Ser-Sch)2] (20 μg/ml)
Group 7: (Ser-Sch)
Group 8: 5 μM AFB1 + (Ser-Sch) (5 μg/ml)
Group 9: 5 μM AFB1 + (Ser-Sch) (10 μg/ml)
Group 10: 5 μM AFB1 + (Ser-Sch) (20 μg/ml)
Group 11: [Mn(Thr-Sch)2] 40 μM
Group 12: 5 μM AFB1 + [Mn(Thr-Sch)2] (5 μg/ml)
Group 13: 5 μM AFB1 + [Mn(Thr-Sch)2] (10 μg/ml)
Group 14: 5 μM AFB1 + [Mn(Thr-Sch)2] (20 μg/ml)
Group 15: (Thr-Sch) 40 μM
Group 16: 5 μM AFB1 + (Thr-Sch) (5 μg/ml)
Group 17: 5 μM AFB1 + (Thr-Sch) (10 μg/ml)
Group 18: 5 μM AFB1 + (Thr-Sch) (20 μg/ml)
Group 19: Na[Mn(Tyr-Sch)2] (40 μM)
Group 20: 5 μM AFB1 + Na[Mn(Tyr-Sch)2] (5 μg/ml)
Group 21: 5 μM AFB1 + Na[Mn(Tyr-Sch)2] (10 μg/ml)
Group 22: 5 μM AFB1 + Na[Mn(Tyr-Sch)2] (20 μg/ml)
Group 23: (Tyr-Sch) (40 μM)
Group 24: 5 μM AFB1 + (Tyr-Sch) (5 μg/ml)
Group 25: 5 μM AFB1 + (Tyr-Sch) (10 μg/ml)
Group 26: 5 μM AFB1 + (Tyr-Sch) (20 μg/ml)
For the micronucleus (MN) analysis, Cytochalasin-B was added 44 h after phytohemagglutinin (PHA) stimulation, to a final concentration of 3 g/ml. Twenty-eight hours later (after 72 h of cultivation), the cells were harvested by centrifugation (1000 g × 10 min). The supernatant was removed, the cells were mixed thoroughly, and 5 ml of cold hypotonic solution (0.05 M KCl) was added. The cells were subsequently incubated at 37°C for 20 min and centrifuged again (1000 g × 10 min). The pellet was mixed thoroughly, and 5 ml of fresh fixative (in a ratio of 1/3, acetic acid/methanol) was added drop-wise. This fixation procedure was repeated three times, and the tube was centrifuged again. The cell pellet was then re-suspended in 1 ml of fresh fixative, dropped onto a clean microscope slide, incubated at 37°C or at room temperature overnight, and stained with Giemsa dye. Coded slides were scored blind by two independent individuals. Only binucleated cells were scored for MN analysis. For each subject, at least 2000 binucleated cells were analyzed for the presence of MN. For the MN scoring, the criteria for micronucleus described by Countryman and Heddle, were followed: a diameter of less than 1/3 of the main nucleus, non-refractility, not touching, and of the same color as the nucleus, or lighter.
The MN frequencies were statistically analyzed by the chi-square test. A value of P less than 0.05 was accepted as being statistically significant. For these procedures, SPSS 11.5 for Windows (SPSS Inc., Chicago, Illinois, USA) was used.
Results and discussion
IR, UV-visible, and NMR spectra of Schiff bases and their Mn(III) complexes
The analytical data and some of the physical properties of the Schiff bases and their complexes are summarized in . The complexes are only soluble in DMF and DMSO, but insoluble in organic solvents like C2H5OH, CCl4, and benzene. summarizes the main IR and UV-visible bands of the azomethines (Schiff bases) and their Mn(III) complexes. In the IR spectra of the Schiff bases, the most characteristic bands appear at 1714–1613 cm− 1, which are attributable to v(C = O) and v(C = N) stretching of the keto and imine forms (CitationGunduz et al. 1994). These bands are shifted in the complexes, which means that the imino nitrogen and phenolic oxygen of the ligands are coordinated to the Mn(III) ion. The 1H-NMR spectrum of the amino acid-Schiff bases, recorded in DMSO-d6, showed the following signals: phenolic –OH proton at 14.09–14.02 ppm (1H), –COOH protons at 10.92–9.25 ppm (1H), aromatic–H proton at 6.60–8.05 ppm (6H), phenyl as a multiplet, and –CH = N– at 9.06–8.75 ppm (1H). The three signals are at 4.70–4.44 ppm and 4.00–3.20 ppm of –CH and –CH2 protons. In the spectra of Schiff bases, two medium intensity bands appear at ∼ 300 nm and ∼ 400–488 nm. It follows from the literature that these bands can be assigned to the phenol-imine and keto-amine forms. They may be attributed to n→ π*, and π→π* type transitions, respectively. Although the bands at ∼ 300 nm exist in the spectra of all complexes, the other bands at ∼ 400 and 420 nm are shifted to shoulder bands. This means that the complex Schiff bases exist only in the phenol-imine form with the Mn(III) ion, coordinated with phenolic oxygen and imine nitrogen. For manganese (III) complexes, a rather broad band appears at 475–510 nm in the visible region, which may be attributed to the 5Eg → 5T2g transition in the octahedral complexes (CitationGunduz et al. 1994).
Table I. Analytical data, important IR vibration frequencies (cm− 1), UV-visible spectra values (nm) of all synthesized molecules and 1H-NMR spectral data of Schiff bases attached L-Threonine, L- Serine and L-Tyrosine.
The biological and anti-mutagenic activities of amino acid-Schiff bases and their Mn(III) complexes
Amino acid-Schiff bases and their Mn(III) complexes were screened for antimicrobial activity in DMF solvent as a control substance. The compounds were tested with the same concentrations in DMF solution (0.25 μg/ml). All the synthesized compounds and antibiotics exhibited varying degrees of inhibitory effect on the growth of different strains tested (, ).
Figure 3. Imaging of antimicrobial affectivities of [Mn(Tyr-Sch)2], (Tyr-Sch) [Mn(Thr-Sch)2], and against B. abartuss, M. luteus.
![Figure 3. Imaging of antimicrobial affectivities of [Mn(Tyr-Sch)2], (Tyr-Sch) [Mn(Thr-Sch)2], and against B. abartuss, M. luteus.](/cms/asset/d924c31a-1ba1-45fe-8e7a-a033db014273/ianb_a_991792_f0003_oc.jpg)
Table II. Antimicrobiyal activity of studied compounds (0.25 μg/ml) and standard reagents (diameter of zone inhibition (mm)).
(Thr-Sch) and (Ser-Sch) bases and their Mn(III) complexes were active against B.abortus. (Tyr-Sch) base and its Mn(III) complexes were more active against P. putida than the other molecules studied. In general, Mn(III) complexes are more potent bactericides than the Schiff bases. This enhancement in activity may be explained on the basis of the chelation theory (CitationSakiyan 2007). Chelation reduces the polarity of the metal ion. Hence, a complex has a lipophilic character, favoring increased interaction between the metal ion and the lipid. This leads to the breakdown of the permeability barrier of the cell wall, resulting in interference with the normal cellular processes. In addition, the antibacterial activity of these compounds was also compared with that of five commercial antibiotics, namely kanamycin, sulfamethoxazole, ampicillin, amoxicillin, and nystatin. It was seen that the synthesized compounds were as effective as the antibiotics mentioned.
Antimutagenic activity
MN frequencies (as mean + SD) of the experimental groups are given in . AFBı caused an increase in MN frequency (). This increase was statistically significant (p < 0.005). However, the MN frequency decreased significantly after treatment with AFB1 and [Mn(Ser-Sch)2], (Ser-Sch), [Mn(Thr-Sch)2], (Thr-Sch), [Mn(Tyr-Sch)2], and (Tyr-Sch). Such a decrease was found to be statistically significant (P < 0.001 and < 0.05).
Table III. Comparison the effects on the number of MN different concentrations of (Thr-Sch).
We know that a micronucleus (MN) is formed during the metaphase/anaphase transition of mitosis (cell division), and is a biomarker of genotoxic events and chromosomal instability. A MN can originate during anaphase from a lagging acentric chromosome or chromatid fragments caused by disrepair of DNA breaks or unrepaired DNA breaks.
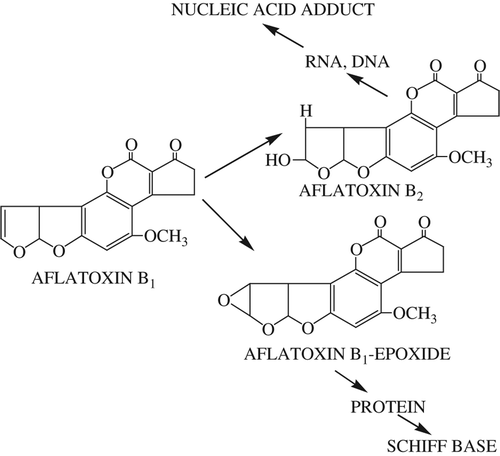
Aflatoxin B2 and Aflatoxin B1-epoxide are formed by oxidation of the 8,9-vinyl ether bond in AFB1 ( and ). The AFB1-epoxide may bind to nuclear DNA, resulting in nuclear damage due to electrophilic attack on the N7 position of guanine of DNA and RNA. As a result, adduct formation in vivo may occur, leading to the transformation of cells, or even cell death, depending on the severity of impairment of template activity (CitationSarı et al. 2013, CitationJacobsen et al. 1991, CitationIrie et al. 1991).Therefore, AFB1-epoxide is not formed or must be neutralized. In this study, AFB1 or AFB1-epoxide may be neutralized due to the type of hydrogen bond or the coordination structure ( and ).
Figure 4. Some products formed by oxidation of AFB1.
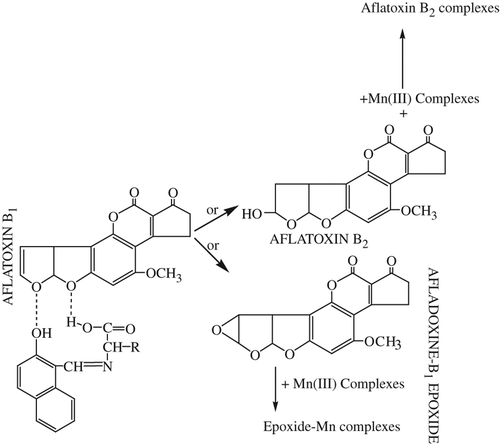
The presence of electronegative groups like oxygen, nitrogen, and sulfur in molecules may be beneficial in reducing the effect of AFB1 by the hydrogen bond (CitationHandique and Aprem 1997). Furthermore, a coordinate bond may form between AFB1-epoxide and Mn(III) complexes, similar to the mechanism of the Jacobsen-Katsuki epoxidation (CitationSarı et al. 2013, CitationJacobsen et al. 1991, CitationIrie et al. 1991). Thus, the Mn(III) complexes synthesized are more effective than their amino acid-Schiff bases.
In addition, it should be noted that the antimutagenic effects of these compounds may be related to their action on the enzymatic activation system. This effect shown by the compounds studied may be attributed primarily to their antioxidant action, or to their function as cofactors for enzyme systems, which is known to protect DNA and other cellular components from damage by oxygen radicals. Consequently, in the present study, it has been revealed that these compounds are active inhibitors of the mutagenic action of AFBı.
In summary, compounds attached to amino acids were prepared for preliminary screening as antimicrobial and antimutagenic agents. The material exhibited very good antimicrobial activity against a wide range of microorganisms. The results from this study show that antimutagenic activity and antimicrobial affectivity are compatible with each other.
Declaration of interest
The authors report no declarations of interest. The authors alone are responsible for the content and writing of the paper.
Funding
This research was supported by the Gazi University Research Fund (Project number: 05/2010-03).
References
- Coluccia M, Correale M, Fanizzi FP, Giordano D, Maresca L, Mariggiò MA, et al. 1984. Mutagenic activity of some platinum complexes: chemical properties and biological activity. Toxicol Environ Chem. 8:1–8.
- Gunduz T, Gunduz N, Sakiyan I. 1994. A new method for synthesis of manganese(III) acetate dihydrate. Synth React Inorg Met-Org Chem. 24:519–524
- Handique AK, Aprem H. 1997. Antimutagenic activity of tryptophan and alanine. Curr Sci. 72:578–580.
- Hartmann M, Hartwig A. 1998. Disturbance of DNA damage recognition after UV-irradiation by nickel(II) and cadmium(II) in mammalian cells. Carcinogenesis. 19:617–621.
- Irie R, Noda K, Ito Y, Matsumoto N, Katsuki T. 1991. Catalytic asymmetric epoxidation of unfunctionalized olefins using chiral (salen) manganese(III) complexes. Tetrahedron Asymmetry. 2:481–494.
- Jacobsen EN, Zhang W, Muci AR, Ecker JR, Deng L. 1991. Highly enantioselective epoxidation catalysts derived from 1,2-diaminocyclohexane. J Am Chem Soc. 113:7063–7064.
- Kuhl A, Hahn MG, Dumić M, Mittendorf J. 2005. Alicyclic beta-amino acids in medicinal chemistry. Amino Acids. 29:89–100.
- Landini D, Maia A, Pina C. 2004. Ring opening reactions of 1,2-epoxides catalyzed by complexes of polyether ligands with metal halides in low polarity solvents. Comparison with heterogeneous systems. Arkivoc 3:202–212.
- Liping L, Jinjun Y, Caixia Y, Miaoli Z, Hong H, Zhiwei L, Maolin G. 2011. Ternary oxovanadium (IV) complexes with amino acid- Schiff base and polypyridyl derivatives: Synthesis, characterization, and protein tyrosine phosphatase 1B inhibition. J Inorg Biochem.105:1323–1328.
- McConathy J, Martarello L, Malveaux EJ, Camp VM, Simpson NE, Simpson CP, et al. 2002. Radiolabeled amino acids for tumor imaging with PET: Radiosynthesis and biological evaluation of 2-amino-3-[18F]fluoro-2-methylpropanoic acid and 3-[18F]Fluoro-2-methyl-2-(methylamino)propanoic acid. J Med Chem. 23: 2240–2249.
- McLean M, Dutton MF.1995. Cellular interactions and metabolism of aflatoxin: an update. Pharmacol Ther. 65:163–192.
- Roy MK, Kuwabara Y, Hara K, Watanabe Y, Tamai Y. 2002. Antimutagenic effect of amino acids on the mutagenicity of N-methyl- N’-nitro-N-nitrosoguanidine (MNNG). Biosci Biotech Biochem. 66: 1400–1402.
- Sakiyan I. 2007. Synthesis and characterization of four new manganese(III) complexes and amino acid (L-Aspartic Acid, L-Asparagine, L-Glutamic Acid, L-Glutamine) Schiff bases. Trans Met Chem. 32:131–135.
- Sarı N, PiŞkin N, Öğütcü H, Yetim KN. 2013. Spectroscopic characterization of novel D-aminoacid-Schiff bases and their Cr(III) and Ni(II) complexes as antimicrobial agents. Med Chem Res. 22: 580–587.
- Snow ET. 1992. Mechanistic implications. Pharmacol Ther. 53:31–65.
- Tavares DC, Cecchi AO, Antunes LM, Takahash CS. 1998. Protective effects of the amino acid glutamine and of ascorbic acid against chromosomal damage induced by doxorubicin in mammalian cells. Teratog Carcinog Mutagen. 18:153–161.
- Zasukhina GD, Vasi'eva IM, Mikhal'chik ES, Durnev AD, Gromov SP, Fedorova OA, Alfimov MV. 2003. Antimutagenic and antioxidant activities of crown compounds in comparison with the effects of garlic extract. Bull Exp Biol Med. 135:261–264.