Abstract
We report the first case of severe pneumonia due to co-infection with the emerging avian influenza A (H5N1) virus subclade 2.3.2.1 and Mycoplasma pneumoniae. The patient was a returning traveller who had visited a poultry market in South China. We then review the epidemiology, virology, interspecies barrier limiting poultry-to-human transmission, clinical manifestation, laboratory diagnosis, treatment and control measures of H5N1 clades that can be transmitted to humans. The recent controversy regarding the experiments involving aerosol transmission of recombinant H5N1 virus between ferrets is discussed. We also review the relative contribution of the poor response to antiviral treatment and the virus-induced hyperinflammatory damage to the pathogenesis and the high mortality of this infection. The factors related to the host, virus or medical intervention leading to the difference in disease mortality of different countries remain unknown. Because most developing countries have difficulty in instituting effective biosecurity measures, poultry vaccination becomes an important control measure. The rapid evolution of the virus would adversely affect the efficacy of poultry vaccination unless a correctly matched vaccine was chosen, manufactured and administered in a timely manner. Vigilant surveillance must continue to allow better preparedness for another poultry or human pandemic due to new viral mutants. Emerging Microbes & Infections (2012) 1, e25; doi:10.1038/emi.2012.24
Keywords:
Case summary
A 59-year-old woman, who had no known underlying disease, first presented at the accident and emergency department on 12 November 2010, with 1 week of fever associated with haemoptysis, dyspnea, sore throat and rhinorrhea. She travelled to Shanghai, Nanjing and Hangzhou for 10 days and returned to Hong Kong on 1 November 2010. She visited a wet market while she was in Shanghai, but she denied any direct contacts with birds or poultry. Upon admission, her body temperature was 38.7 °C, with a respiratory rate of 24 breaths per minute and oxygen saturation of 96% while breathing ambient air. Chest radiograph revealed left middle zone consolidation. The clinical diagnosis was acute community-acquired pneumonia, for which she was treated as an outpatient with 1 g of oral amoxicillin–clavulanate twice daily.
Two days later, her symptoms of dyspnea and haemoptysis worsened, and she was admitted to the hospital. Chest radiograph showed progression of consolidation, involving both left middle and lower zones. She required oxygen supplementation via a nasal cannula because her oxygen saturation was 88% while breathing ambient air. Blood tests upon admission showed a total leukocyte count of 4.2×109/L, an absolute neutrophil count of 3.8×109/L and lymphopenia of 0.3×109/L. Liver enzymes were elevated, with an alkaline phosphatase level of 223 U/L and an alanine transaminase level of 71 U/L. The values for haemoglobin, platelet count and renal function test were within the normal range. Oral azithromycin at a dose of 500 mg once daily was added to treat atypical agents of pneumonia. A nasopharyngeal swab collected on the day of admission tested negative for influenza A and B by enzyme immunochromatographic assay. On day 3 of hospitalisation, she developed type I respiratory failure, required 100% oxygen supplementation via a non-rebreathing mask and was admitted to the intensive care unit. Antibiotics were switched to 2 g of intravenous ceftriaxone every 24 h and 100 mg of oral doxycycline every 12 h for enhanced coverage of bacterial pathogens. Because of her clinical deterioration, a nasopharyngeal swab sample was tested by polymerase chain reaction (PCR) and reverse transcriptase-PCR (RT-PCR) for Mycoplasma pneumoniae and influenza A virus, respectively, which were both positive. Other tests for Chlamydophila pneumoniae, Legionella and human metapneumovirus were negative. The RT-PCR for influenza A virus subtype H5 was positive. The antibody titre against influenza A by complement fixation test increased from 80 on admission (7 days after onset of symptoms) to 2560 on day 5 after admission. Microbiological investigations on the sputum sample, including Gram smear and bacterial culture, Ziehl–Neelsen staining and PCR for Mycobacterium tuberculosis, were negative. Oral oseltamivir at a dose of 150 mg every 12 h and 100 mg of oral amantadine every 12 h were started on day 3 and day 4 of hospitalisation, respectively. Her condition gradually improved. Oxygen supplementation was weaned off 10 days after admission. A total of 15 days of oseltamivir and 6 days of amantadine therapy were administered.
Computed tomography of the thorax was performed on day 24 of hospitalisation and revealed left pleural effusion with a small loculated pleural effusion over the anterior aspect of left hemithorax, subsegmental collapse consolidation with bronchiectatic changes over the left lobe, and patchy ill-defined consolidation and ground glass opacities with adjacent interstitial thickening in both lungs. Ultrasound-guided aspiration of the left pleural fluid was performed. Analysis of the pleural fluid showed a pH of 7.51, glucose level of 5.2 mol/L, total protein level of 48 g/L, lactate dehydrogenase level of 256 U/L and adenosine deaminase level of 16 U/L. The cell count was not performed because the sample was blood stained. The Gram stain and Ziehl–Neelsen stain were negative. Bacterial and mycobacterial culture results were negative. She was discharged after 29 days of hospitalisation.
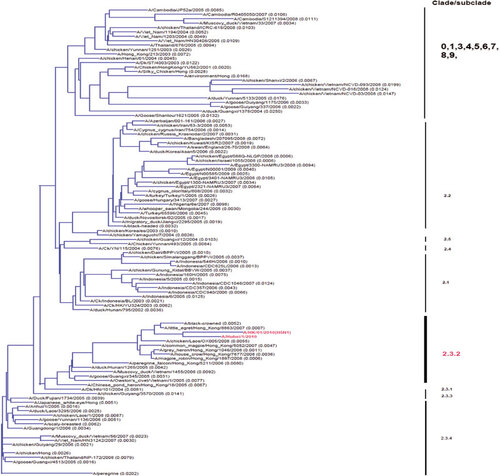
The avian influenza A (H5N1) virus was isolated using the Madin–Darby canine kidney cell line from the nasopharyngeal swab collected on the day of admission. Viral genome sequencing and analysis confirmed the features of highly pathogenic avian H5N1 virus. No unusual mutations associated with pathogenic properties, including known polymorphisms that affect receptor binding specificity, were identified. Phylogenetic analysis of haemagglutinin (HA) showed that the virus belongs to subclade 2.3.2.1 ().
Review of literature and discussion
Epidemiology
Avian influenza viruses H5N1,Citation1 H9N2,Citation2 H7N7,Citation3 H7N2,Citation4 H7N3Citation5 and H10N7Citation6 have jumped species barriers and caused human infection. Among these avian influenza viruses, H5N1 virus is the most virulent, with a crude mortality rate of 60%, which markedly surpasses the mortality rate of pandemic influenza virusesCitation7 (). The H5N1 virus was first documented to cause human infections in Hong Kong in 1997.Citation1 It reappeared again in 2003 in Hong Kong, China and Vietnam and has spread to other parts of Asia, Africa and Europe.Citation8,Citation13 Since 2009, Egypt has had the highest incidence of human H5N1 virus infection, temporally associated with a high incidence of H5N1 virus infection in its poultry population.Citation14 According to the World Health Organization, there were more than 600 confirmed human cases up to May 2012.Citation8 However, the number of human cases is likely an underestimate, and a meta-analysis suggested an overall seropositive rate of 1.2% among the exposed population.Citation15
Table 1 Table 1 Human cases of H5N1 virus infectionFootnote*
Seasonal variations in the incidence of H5N1 virus infection have been observed in some countries. In Egypt, H5N1 virus infection peaks during late winter and early spring, which are associated with low precipitation and moderate humidity and temperature.Citation16 However, the seasonal pattern is less clear for Indonesia.
H5N1 virus infection affects individuals of all ages. Similar to pandemic influenza, adolescents and young adults are disproportionately affected.Citation13,Citation17,Citation18 Our patient was 59 years of age, which is older than the median age of 18 years for patients with H5N1 virus infection. However, the age of patients with H5N1 virus infection differs widely in different countries. In Egypt, the median age is 6 years, whereas in Asian countries, the 20- to 29-year age group is most affected.Citation13 The relatively lower incidence in the elderly population may be explained by prior exposure to various types of poultry or human influenza viruses, which may lead to cross-reactive immunity. In mice, prior seasonal influenza virus infection reduced the severity of H5N1 virus infection.Citation19 Age may be associated with the severity of illness, but contrary findings have been reported. In the first outbreak of Hong Kong in 1997, all patients aged over 13 years had severe disease, while most children only had mild upper respiratory tract symptoms.Citation1 However, a study in Vietnam showed that patients 16 years of age or younger had a higher risk of death than older patients.Citation20
Although migratory birds are often infected with H5N1 virus, live domestic poultry are thought to be the main source of human infections.Citation21 Our patient did not have direct contact with live poultry, although she did visit a wet market. A multivariate analysis showed that visiting a wet poultry market and being in indirect contact within 1 m of sick or dead poultry are independent risk factors for acquiring H5N1 virus infection.Citation22 Many environmental samples from wet markets have been contaminated with the H5N1 virus,Citation23 suggesting that indirect contact can be a mode of transmission of H5N1 virus. Although human clusters occur, human-to-human transmission is still limited and usually requires a prolonged period of contact with infected individuals.Citation24 Consumption of raw duck blood is suspected for some cases of human H5N1 virus infection,Citation25 and this mode of transmission is compatible with studies in mammals that intragastric inoculation of H5N1 virus can lead to systemic dissemination via the lymphatics and venous route.Citation26 H5N1 virus can also be detected in frozen poultry and contaminated eggs, but transmission to humans via these food items has not been documented.Citation27
While H5N1 virus is highly pathogenic to human, the incidence of human infection is still low. Some identified factors may reduce the human susceptibility to this virus. First, most H5N1 viruses have a predilection to attach to the avian-like α2,3-linked sialic acid receptor (α2,3 SA) rather than the human-like α2,6-linked sialic acid receptor (α2,6 SA). All pandemic or seasonal epidemic human influenza viruses have preferential binding for α2,6 SA, which is abundant in the upper respiratory tract in human and may be a prerequisite for efficient human-to-human transmission.Citation28 Second, in addition to the host cell surface receptor requirement, the avian- and human-adapted influenza viruses also have different importin-α isoform requirements.Citation29 Importin-α is a cellular protein responsible for the entry of the viral ribonucleoprotein complex into the host cell nucleus. Efficient replication of avian-adapted influenza strains is dependent on importin-α3, while that of human-adapted strains is dependent on importin-α7. Furthermore, when mice are infected by human influenza viruses, viral dissemination is less prominent in mice lacking importin-α7 than wild-type mice. This specificity has been attributed to the D701N and N319K substitutions of the PB2 (polymerase subunit PB2) and NP (nucleoprotein) genes, respectively. Third, the H5N1 virus may be more susceptible to the human host defence system. Interferon(IFN)-induced Mx GTPases are important in the suppression of H5N1 but not the pandemic influenza A H1N1 2009 (A(H1N1)pdm09) virus due to differences in the NP, which partially explains the decreased susceptibility of human H5N1 virus infection.Citation30 Finally, mutations that are responsible for human adaptation seem to be unstable in H5N1 viruses.Citation31
Virology and virulence factors
The influenza virus is an RNA virus belonging to the family of Orthomyxoviridae. Its genome consists of eight gene segments that potentially encode 12 proteins. Influenza viruses are typed by their matrix and NPs antigenically into types A, B and C. The influenza A virus is further subtyped by its surface HA and neuraminidase (NA) into many subtypes as various combinations of 17 different HAs and 10 different NAs.Citation32 The avian H5N1 virus can be classified into genotypes or clades. Based on the sequences of internal genes, the H5N1 virus was classified into genotypes designated A, B, C, D, E, P, V, W, X0-3, Y, Z and Z+, which are seldom used in most recent studies.Citation33 Reassortments between different genotypes occur frequently in avian species.Citation34 More commonly, the H5N1 virus can be classified into clades/subclades based on the phylogeny of HA. The World Health Organization/World Organization for Animal Health/Food and Agriculture Organization H5N1 Evolution Working Group proposed a unified nomenclature system using clades.Citation35 A clade is further subdivided into second-order clades when it evolves and satisfies the criteria for discrete clades. The H5N1 progenitors closest to Gs/Guangdong/1/96 and closely related to the 1997 human isolates from Hong Kong are designated as Clade 0. Avian H5N1 viruses have evolved rapidly since 2003, when mainly clade 1 viruses were identified. Since 2004, clade 2 viruses emerged and subsequently evolved into different subclades in different geographical regions. Human infections are caused by H5N1 viruses in clades 0, 1, 2 and 7 ().
Clade 1 originated from Yunnan in 2002.Citation36 The clade has further diverged into subclades 1.1 and 1.2. Human cases due to clade 1 viruses have been found in Hong Kong, Vietnam, Thailand and Cambodia.Citation12,Citation37,Citation38,Citation39 Mutation 701N of PB2 is critical for the virulence of this clade.Citation40 The N66S mutation in PB1-F2 protein, which has been associated with virulence in mammals,Citation41 has been identified in two strains in subclade 1.1 in Cambodia in 2010 and 2011.Citation42 Clade 1 viruses are more virulent than clade 2 viruses in a ferret model with a limited number of isolates.Citation43
Viruses in clade 2 or its subclades have become the predominant clade in human infections since 2005. Subclade 2.1, with the majority of these viruses in the 2.1.3 subclade, is the only clade found in Indonesia, which has the highest cumulative number of H5N1 human cases and deaths in the world.Citation44 Subclade 2.2 virus (also called Qinghai-like virus) caused a large outbreak among wild birds around Qinghai Lake in 2005 and has since spread to other parts of Asia, Europe and Africa.Citation45 Human H5N1 cases due to subclade 2.2 viruses are mainly found in the Middle East (particularly in Egypt), South Asia and China.Citation46 This clade has diverged sufficiently such that subclades are designated. Subclade 2.2.1 viruses emerged in Egypt in 2008 and are associated with increased binding avidity to α2,6 SA, which may explain the increased incidence of human cases in the affected countries.Citation47 Another distinct feature of subclade 2.2 viruses is the E627K substitution of the PB2 protein, which is only found in some human cases infected by other subclades of H5N1 viruses.Citation48 Subclade 2.3 represents another major variant besides 2.1 and 2.2 described above. Variants of subclade 2.3.4 viruses first emerged in humans in 2005 in Anhui, became the predominant variant found in poultry in southern China and are responsible for most human infections in China.Citation49 In Vietnam, subclade 2.3.4 first appeared in humans in 2005 and has been the predominant clade since 2007.Citation50 In addition, this subclade is also found in other parts of Southeast Asia.Citation38 Two strains in the subclade 2.3.4 have increased binding affinity to α2,6 SA.Citation51 627K of PB2 in subclade 2.3.4 viruses may also contribute to its virulence.Citation40 Subclade 2.3.4.2 has also been found in Bangladesh and Myanmar.Citation11 Subclade 2.3.2 was first found in China and Vietnam in 2005.Citation52 A new subclade, 2.3.2.1, has been found in migratory birds and poultries from China, Japan and Bangladesh since 2009.Citation53,Citation54 Subsequently, the first human case caused by a 2.3.2.1 virus was found in Guangxi in 2009, and additional cases, including our presently reported case, have been infected by this new variant.Citation10,Citation55 The dead wild bird and poultry surveillance in Hong Kong showed that the dominant isolate before 2007 belonged to subclade 2.3.4, but most of the isolates found after 2007 belonged to subclade 2.3.2.1 (personal communication with Dr Thomas Sit of Agriculture Fishery and Conservation Department of Hong Kong Special Administrative Region). However, the detection of a human case in this study suggests that subclade 2.3.2.1 has emerged as the dominant variant in some regions of China.
Clade 7 viruses were first isolated from human in 2003 and subsequently found in Anhui in 2006. Viruses from this clade were also detected in chickens in Vietnam.Citation56 This clade has not been detected in humans since 2007. Data on the genetic evolution of this clade are scarce.
It is notable that among human cases, the fatality rate was 83.1% in Indonesia but only 35.9% in Egypt (). Because subclades 2.1 and 2.2 are exclusively circulating in Indonesia and Egypt, respectively, certain subclades may be more virulent than others. Sequence comparisons revealed some variations between different geographical subclades (). When comparing the known polymorphisms responsible for virulence or mammalian adaptation between the human strains from different countries, most mutations enriched in the Egypt strains may have enabled the strains to bind better to the α2,6 SA. For example, deletion of position 129 together with I151T substitution of HA is found in 40% of strains from Egypt but not from Asia (H5 numbering). This double mutation increases the binding to α2,6 SA, and the virus can attach mainly to type I and type II pneumocytes but minimally to the larynx in human.Citation47 T156A of HA is also present in most strains from Egypt but only in a few strains in China and none from Southeast Asia. This mutation results in loss of glycosylation at positions 154–156 and is associated with increased binding to α2,6 SA and increased transmission efficiency among guinea pigs.Citation60 Another major difference is the S235P mutation, which is present in most strains from Asia but only in 10% of strains from Egypt. However, this mutation does not change the affinity for α2,6 SA in direct binding assays.Citation47 Polymorphisms in PB2 are also important in mammalian adaptation, but these polymorphisms between strains from Egypt and other countries cannot be compared because the PB2 amino-acid sequences for only two Egyptian strains from humans have been deposited into GenBank. However, the 627K of PB2 is more common among isolates from Vietnam/Thailand/Cambodia/Laos (50%) than from Indonesia (11%) and China/Hong Kong (20%). Only further investigations can ascertain the relevance of these mutations to their effect on virulence in humans. Besides viral genetic polymorphisms, the difference in fatality may also be related to the availability and standard of medical care in these countries. H5N1 viruses circulating in Cambodia are similar to those in Thailand, Vietnam and China, but human cases from Cambodia exhibited the highest mortality rate (90.5%; ). Close monitoring and examination of genetic variations in different geographical areas are necessary for a better understanding of the virulence and molecular evolution of the H5N1 viruses.
Table 2 Table 2 Natural mutations in human isolates of the H5N1 virus that are associated with virulence or adaptation to humans. Gene information obtained from the NCBI Influenza Virus Reource,Citation57 sequences with duplicate strain names but different sequences were excluded from the analysis.
Clinical and laboratory features
The usual incubation period is 2 to 9 days.Citation24,Citation64 The incubation period was usually longer in patients who acquired the infection from the wet market than those patients who were exposed to sick or dead poultry.Citation65 Our patient had a typical course of the disease, which manifested with upper respiratory tract symptoms and was followed by rapid deterioration into pulmonary disease. Haemoptysis is an alarming symptom that may signify pulmonary haemorrhage. Acute respiratory distress syndrome (ARDS) is a frequent pulmonary complication.Citation66 H5N1-induced ARDS is associated with poor outcomes even with prompt medical care. In contrast to human seasonal influenza viruses, H5N1 viruses are more likely to cause severe pneumonia. This finding is consistent with the fact that H5N1 viruses preferentially bind to α2,3 SA, which is abundant on the surface of type II pneumocytes, alveolar macrophages and non-ciliated bronchiolar cells of the lower respiratory tract.Citation67 Although H5N1 virus can replicate in ex vivo upper respiratory tract cell lines,Citation68 binding to upper respiratory tract tissue is much less efficient when compared with the seasonal influenza viruses.Citation69 Some H5N1 viruses isolated from humans can recognize both α2,3 SA and α2,6 SA, such as A/Hong Kong/213/03 (H5N1) and some sublineages of the subclade 2.2.1 viruses from Egypt and subclade 2.3.4 viruses from China.Citation47,Citation51,Citation70 The dual-binding capability may indicate a wider range of cellular tropism and, therefore, more efficient spread of this virus in the respiratory tract. However, the ability to bind α2,6 SA in vitro may not equate to binding of α2,6 SA in human. In the human respiratory tract, α2,6 SAs have long oligosaccharide branches with multiple lactosamine repeats, which is different from the short α2,6 SA used in previous studies.Citation71 This study also demonstrated that influenza viruses that transmit efficiently between humans have high affinity for the long α2,6 SA, while those viruses with poor transmissibility only bind the short α2,6 SA. The difference in binding specificity between the long and short α2,6 SA may be related to their structures. The long glycans exhibit an umbrella-like topology that binds onto a wider region of the receptor-binding pocket of HA, while the short glycans that bind a smaller region of HA have a cone-like topology.
In addition to pulmonary disease, H5N1 virus infection also leads to extrapulmonary manifestations more often than infections caused by pandemic influenza viruses.Citation72 Diarrhea occurred in more than 50% of patients in a series from VietnamCitation9 but less than 10% in other series.Citation20 Liver impairment can occur, and our patient had elevated alkaline phosphatase and alanine transaminase. Renal impairment is common. Elevated creatine kinase is also common, but true rhabdomyolysis has not been described. In addition, encephalitis may occur. Many of these features are attributable to the direct invasion of the virus, and the multibasic cleavage site of HA, which allows cleavage by proteases that are ubiquitously expressed in most cells, is critical for the systemic spread of the virus.Citation73 The higher virulence of the H5N1 virus, which leads to a high pulmonary and extrapulmonary viral load, together with its intrinsic pro-inflammatory property, often causes a cytokine storm in these patients. Other common laboratory abnormalities included leucopoenia, lymphopenia, thrombocytopenia and impaired coagulation profiles.Citation1 Reactive haemophagocytic syndrome can occur and may be related to prolonged IFN-γ production.Citation74 Encephalopathy has been reported.Citation74 Reye syndrome can occur after aspirin is administered.Citation1
Viral antigens can be detected in small and large intestinal epithelial cells, bone marrow and the brain, but viral cultures from these organs are negative.Citation74,Citation75 In addition, H5N1 RNA can be detected in the spleen,Citation76 and the H5N1 virus can be cultured in the cerebrospinal fluid of infected humans.Citation77 In vitro, the H5N1 virus can replicate in cell lines originating from different anatomical sites.Citation78 In ferrets, H5N1 can cause encephalitis and non-supportive vasculitis with haemorrhage via infection of the olfactory system.Citation79 In a mouse model, H5N1 infection has also led to aggregated alpha-synuclein, prolonged microgliosis and loss of dopaminergic neurons, which is compatible with neurodegenerative diseases.Citation80
In addition to virus-induced cytolysis driving this pathogenesis, the pulmonary and systemic manifestations of H5N1 virus infection may be related to the cytokine storm. The H5N1 virus can induce strong cytokine responses in vitro and in vivo.Citation81,Citation82 Infection of pulmonary microvascular endothelial cells has been associated with a marked inflammatory response.Citation83 Nuclear factor κ-light-chain-enhancer of activated B cells (NF-κB) signalling is essential for the expression of IFN-β in H5N1 virus-infected endothelial cells.Citation84 Upregulation of genes associated with keratin filaments and keratinisation in human bronchial epithelial cells may be related to hypercytokinaemia.Citation85 Viruses that have high affinity for α2,3 SA may induce a stronger inflammatory response in human dendritic, macrophages and respiratory epithelial cells than viruses with high affinity for α2,6 SA.Citation86 In addition to the pro-inflammatory cytokine response, the H5N1 virus also inhibits a lipoxin-mediated anti-inflammatory response.Citation87
Poor prognostic factors include older age, delay in hospitalisation, diarrhoea, mucosal bleeding, chest X-ray showing extensive lung involvement, desaturation on admission, a delay in receiving antiviral therapy, neutropaenia, thrombocytopaenia, and increased levels of d-dimer, serum glucose, urea, alanine aminotransferase and lactate dehydrogenase.Citation1,Citation20,Citation65,Citation88 Secondary community-acquired bacterial infection is a well-established poor prognostic factor for influenza virus infection,Citation89 but its role has not been demonstrated conclusively for H5N1 virus infection because most of the co-infections that occurred after H5N1 infection were due to hospital-acquired bacteria or fungi.Citation1 For our patient, M. pneumoniae was detected in the nasopharyngeal swab sample collected on the day of admission and is therefore acquired from the community. Our case is the first report of H5N1 virus infection with community-acquired co-infection by M. pneumoniae. Although our patient started taking azithromycin, the progression of pneumonia may have been related to a macrolide-resistant strain of M. pneumoniae.Citation90 Therefore, it is difficult to delineate the relative contribution of M. pneumoniae in our patient's clinical presentation. Treatment with corticosteroids is associated with increased mortality.Citation20 Higher viral load in the respiratory tract is associated with poor outcome in humans.Citation91 The higher viral load may be related to the poor innate immune control of the virus. Viraemia is also associated with a poor outcome of H5N1 infection in a ferret model, although this association has not been systematically analysed in human H5N1 cases. Viral genetic polymorphisms are associated with increased virulence in animal models.Citation59 However, none of these polymorphisms has been associated with more severe outcome in human H5N1 infection, unlike the D222G substitution in A(H1N1)pdm09 virus, which is clearly correlated with more severe disease in humans.Citation92,Citation93 As expected, genetic polymorphisms such as the polymorphisms of CD55, are associated with severe A(H1N1)pdm09 infection.Citation94
Although H5N1 infection is usually associated with severe disease, some patients, especially children, only suffer from mild upper respiratory tract symptoms without pneumonia.Citation1,Citation9,Citation88 The disease severity also varies between patients from different countries. For example, Egypt has a particularly low rate of pneumonia, which is still unexplained but may be attributed to the higher predilection of the Egyptian virus for the upper airway.Citation9
Diagnosis
Currently, antigen detection by rapid immunochromatographic assays or direct immunofluorescence and nucleic acid detection by RT-PCR provides a rapid diagnosis, which guides immediate management; in contrast, viral culture and serology allow for retrospective diagnosis, which is essential for epidemiological studies. Respiratory tract specimens are the best samples for detecting the virus, although the virus can also be found in blood or rectal swabs.Citation91 Unlike human seasonal influenza viruses, a throat swab has a higher yield than a nasopharyngeal swab for the H5N1 virus due to the general predilection of the H5N1 virus for the lower respiratory tract. Although technically simple, immunochromatographic assays have a low sensitivity, and their detection limits are at least 3 log10 TCID50 per 100 µl.Citation95 Therefore, further diagnostic tests were performed for our patient even though the immunochromatographic assay showed a negative result for influenza A and B. Direct immunofluorescence assays also have poor sensitivity.Citation1 Another limitation of antigen detection assays is their failure to differentiate H5N1 from other influenza A viruses. RT-PCR is the most sensitive detection method,Citation96 but mutations in the HA gene can cause false negatives due to mismatches between the viral gene sequence and the primers or the probes.
Serology is especially useful in documenting asymptomatic infections, but is not useful for patient management because a ≥ 4-fold rise in complement fixation antibody titre in serum samples collected 2 weeks apart is necessary to make the diagnosis of influenza A infection. Another problem is that complement fixation antibody titre is not specific for H5N1 virus infection. For a more specific diagnosis of H5N1 virus infection, the viral microneutralisation assay is more specific than the haemagglutination inhibition assay. Unlike the human H1N1 or H3N2 infections, the viral microneutralisation assay, preferably with confirmation by Western blotting with a baculovirus-expressed H5 protein, is the preferred gold standard for serological assay and is presently accepted as the more specific way of serodiagnosis.Citation97 Most individuals develop a positive titre 3 weeks after the onset of disease.Citation98
Treatment
Adamantanes were used in the initial outbreak of H5N1 in Hong Kong in 1997.Citation1 However, adamantane resistance is now widespread,Citation99 and susceptible strains are limited to those strains in clade 2 from Eurasia and AfricaCitation9 and subclade 2.1 from Indonesia.Citation100 Resistance to adamantanes is due to L26I or S31N mutations of the M2 protein, and the latter mutation reduces drug binding.Citation101
The neuraminidase inhibitors oseltamivir or zanamivir are the mainstay of treatment for H5N1 infection. The survival benefit is greatest if oseltamivir is started within 2 days of symptom onset. It is doubtful if oseltamivir is useful if initiated later, although there is a report of benefit even if given within 8 days after symptom onset.Citation102 Our patient received oseltamivir >8 days after symptom onset, which may explain why our patient developed severe disease. Zanamivir is usually administered by oral inhalation. Intravenous zanamivir, which was successfully used in humans during the 2009 H1N1 pandemic, is effective against H5N1 virus infection in a macaque model.Citation103 Other neuraminidase inhibitors, including peramivir and CS-8958, are also active against the H5N1 virus.Citation104,Citation105 Strains with reduced susceptibility to oseltamivir are present in clade 2 viruses from IndonesiaCitation106 and subclade 2.3.4 viruses from Vietnam.Citation50 Studies reporting the oseltamivir susceptibility of clade 1 viruses from Cambodia have been contradictory. An earlier study showed that these strains had reduced susceptibility to oseltamivir, while a more recent study of the same strains reported full susceptibility.Citation107 Oseltamivir resistance can emerge during treatment, leading to treatment failure.Citation108 The mutations responsible for oseltamivir resistance are most commonly due to H274Y and N294S (N2 numbering) substitutions within the neuraminidase. H5N1 viruses with these mutations are still highly pathogenic.Citation109 Other mutations associated with reduced susceptibility to oseltamivir, including V116A, I222L, K150N and S246N, have also been reported.Citation110 However, oseltamivir-resistant strains are usually susceptible to zanamivir. The discrepancy between oseltamivir and zanamivir susceptibility is due to differences in how oseltamivir and zanamivir bind to the virus. The binding of oseltamivir to the viral neuraminidase requires a conformational change in the side chain of 276E, and thus, the carboxyl group of 276E is oriented away from the hydrophobic pentyloxy group of oseltamivir. In contrast, the binding of zanamivir involves hydrogen bond formation without a side chain conformational change.Citation111 True zanamivir resistance has not been reported, but a clade 1 strain from Cambodia isolated from a human patient had an half maximal inhibitory concentration (IC50) of 3.35 nM, which is eightfold higher than the mean IC50 of other strains.Citation107 Eight amino-acid substitutions were found in the NA of this virus, and the V149A mutation was postulated to be the important substitution responsible for the reduced susceptibility. A/Swan/Shanghai/10/09, an H5N1 virus belonging to the epidemic subclade 2.3.2, contains an S31N mutation in the M2 protein and an H274Y mutation in the NA protein; these mutations are responsible for resistance to adamantanes and neuraminidase inhibitors, respectively.Citation112
The clinical efficacy of oseltamivir for the much milder A(H1N1)pdm09 infection was recently questioned because its clinical benefit appears minimal even if instituted within 48 h of symptom onset.Citation113 Several antivirals in development have targeted other parts of the viral life cycle. Nucleozin, a NP inhibitor, has potent in vitro and in vivo activity against H5N1 virus in a mouse model.Citation114 Antivirals that affect viral RNA synthesis, including 2'-deoxy-2'-fluorocytidine and favipiravir (T-705), also have good in vitro activity.Citation115,Citation116 The sialidase fusion protein DAS181 can inhibit the H5N1 virus, including oseltamivir-resistant strains.Citation117,Citation118 Antiviral peptides that bind to HA, therefore preventing viral entry, are also effective in vitro.Citation119
Immunomodulatory therapy is an attractive treatment option because the currently available antivirals have only modest potency, and antiviral resistance may appear during treatment. COX-2 inhibitors, such as celecoxib, together with zanamivir, improved the survival of H5N1-infected mice.Citation120 Cytotoxic therapy has been proposed as an adjunctive treatment for H5N1, but real-life treatment experience has not been reported.Citation121 Etoposide and betamethasone were beneficial in a patient with A(H1N1)pdm09 infection complicated by haemophagocytic lymphohistiocytosis.Citation122 In guinea pigs, α-IFN treatment reduced lung viral titre,Citation123 but few would consider its use in a highly inflammatory condition such as H5N1 infection. A proteasome inhibitor that prevents NF-κB activation in infected cells reduces cytokine release.Citation124 Convalescent plasma and hyperimmune serum may be useful.Citation125 Monoclonal antibodies against HA of H5N1 appear effective in mouse models.Citation126
Prevention
Most human H5N1 infections have been acquired via direct transmission from infected poultry, and human infections are associated with poultry outbreaks.Citation127 Hence, controlling the infection in poultry is crucial to prevent human infections. Several risk factors have been identified for H5N1 outbreaks in chickens. First, the isolation of H5N1 virus from ducks and geese precedes disease outbreaks and virus isolation in chickens.Citation128 Because infected ducks and geese are often asymptomatic, the amplification and spread of the virus among these poultry often goes unnoticed.Citation129 Therefore, segregating chickens from other poultry is important. In farms, chickens should not be reared with other poultry. In the retail markets, live ducks, geese and quails are now banned in Hong Kong.Citation128 Second, human-bird segregation via a central slaughtering has been proposed in Hong Kong but yet to be implemented due to resistance from the public. Third, the spread of H5N1 virus is associated with the transport of infected chickens.Citation130 Cross-border spread may be prevented by tighter control over illegal poultry trafficking. Fourth, biosecurity measures must be in place for the transport of chickens between farms. Visiting farms is associated with increased risk of poultry outbreak; hence, biosecurity measures should include policies for visitors.Citation131 Fifth, vaccination and disinfection can decrease the risk of poultry outbreaks.Citation130 Vaccination in the poultry population is now practiced in many countries. However, a vaccine against one clade of virus may not protect against other clades or subclades due to antigenic differences.Citation132 Currently, there are four main groups of HA: clade 1 viruses are similar to clade 4, 5, 7 and 9 viruses; subclade 2.1 viruses are antigenically similar to subclade 2.4 viruses; subclade 2.2 viruses are similar to some viruses in subclade 2.3; and subclade 2.3.4 viruses represents a unique group. Further divergence of a particular clade/subclade can lead to vaccine failure, such as the recent emergence of subclade 2.3.2.1 viruses in southern China.Citation133 Due to the rapid mutation of the HA gene, poultry vaccine components must be regularly updated.Citation10
Surveillance of poultry and wild birds has allowed for the early detection of the H5N1virus and has provided important information regarding the spread of H5N1 viruses. For each farm, unvaccinated poultry are also reared because these poultry will be more symptomatic if infected and therefore serve as a sentinel of H5N1 virus infection. Wild bird surveillance allows for the understanding of the virus activity and evolution in the natural reservoir. Because migratory birds can travel long distances, they can introduce or reintroduce H5N1 virus to a disease-free area and may pose a threat to humans.Citation134 In Hong Kong, wetland parks are temporarily closed when the H5N1 virus is detected in the park. Education regarding the safe handling and disposing of wild birds is regularly provided to the public. However, even though pigs are well-known mixing vessels for influenza viruses, pig surveillance is relatively less systematic. Besides early detection of new reassortants or clades of viruses in animals, rapid genetic characterisation of the virus is important for identifying important pathogenetic or drug-resistant genomic signatures. Early detection of human cases is also important. Increased awareness among frontline clinicians, together with sensitive and specific molecular testing, will allow for prompt recognition of an infected case for early administration of antiviral treatment, infection control measures, and tracing the source of infection.
Culling of chicken has been used in Hong Kong in 1997 as a means to terminate the outbreak of human H5N1 infection. It was used again in 2003 and 2011 following the identification of H5N1 virus in poultry in the market. To minimize the expensive mass culling, precautionary measures have been put in place. Twice monthly rest days in the wet markets in Hong Kong allow for a thorough cleansing of the environment which prevents the mixing of old and new chickens and disrupts the chain of virus transmission. During outbreaks, the wholesale market is thoroughly disinfected, and live poultry from farms cannot be sold in the markets for 21 days until there are no additional cases throughout Hong Kong. When the poultry vaccine cannot catch up with the rapid evolutionary of the H5N1 virus, no live poultry is allowed to stay overnight in wet markets. They are either sold out or culled in the evening. Such severe biosecurity measure is necessary to stop transmission in the poultry market which is basically a minifarm in an overcrowded urban area.
Vaccination in human is still at the experimental stage. Human studies have shown that antibodies against HA and NA can be elicited by vero cell- and insect cell-derived vaccines.Citation135,Citation136 Using a low-dose, adjuvanted vaccine with 3.75 µg of HA has been successful.Citation137 Two doses of adjuvanted whole virus vaccine can elicit high titres of cross-neutralising antibodies that are effective against clade 0, clade 1 and clade 2 viruses.Citation138 A low-dose intradermal H5N1 vaccine has been evaluated in a phase I clinical trial, and the elicited immune response was similar to that with the regular dose of intramuscular vaccine.Citation139 Patients previously primed with an H5 vaccine developed a rapid and strong neutralising antibody response upon a second challenge with an antigenically distinct H5 vaccine.Citation140 Recently, Lyall et al.Citation141 proposed that genetically modified chickens expressing a short-hairpin RNA that prevents viral replication are resistant to H5N1 virus infection and may stop transmission of the H5N1 virus.
Controversy on the study of airborne transmission of H5N1
The current strain of H5N1 virus is highly virulent but poorly transmissible between humans.Citation24 Scientists have been examining factors that may give rise to a highly transmissible H5N1 virus and, hence, enhance the preparedness for the arrival of such a virus. Previous studies demonstrated that T156A of HA and 701N of PB2 are critical in the transmission of H5N1 virus in a mammalian host.Citation60 In that study, the virus can be transmitted via direct contact because the guinea pigs were put in the same cage. In another experiment, the K627E substitution of PB2 decreased contact transmission between guinea pigs, while 701N compensated for the lack of 627K.Citation142 While guinea pigs are similar to humans in the distribution of α2,3 SA and α2,6 SA in their respiratory tract, they do not develop the clinical signs observed in humans.Citation143 However, ferrets have a similar sialic acid distribution in the respiratory tract, susceptibility and clinical manifestations to humans; therefore, they are the preferred animal model.Citation144 A recent transmission study in ferrets demonstrated that a reassortant virus containing a mutant haemagglutinin (Q192R, Q222L and G224S), a human H3N2 neuraminidase and other genes from H5N1 virus, was transmitted to one out of two ferrets via the droplet route.Citation145
Recently, mutant viruses capable of more efficient non-contact transmission arising from the passage of the H5N1 virus in ferrets and reverse genetics were found by two groups of scientists.Citation146,Citation147 Whether the results of the research should be published has been controversial because of biosecurity concerns. In December 2011, the National Science Advisory Board for Biosecurity (NSABB) of the USA recommended that the journals not publish key details, and in February 2012, the World Health Organization also decided that the results should be withheld to allow more discussion on the impact of publishing the results.Citation148 Proponents for publishing the results suggest that air transmissibility between ferrets does not mean air transmissibility between humans. Moreover, the findings are important for understanding of the virus, which is a prerequisite for designing better treatment and prevention strategies.Citation149 Knowledge on the genomic signatures that are critical for efficient transmission identified via the ferret studies will allow us to have more targeted surveillance and research for viruses harbouring such signatures, so that early and specific preventive measures can be implemented. The proponents also raised the issue that previous studies on the rescue of the rather virulent 1918 pandemic influenza virus were allowed to be published in full. However, opponents suggested that the results will allow terrorists to replicate the experiment and produce an H5N1 virus capable of aerosol transmission that could be used in biological warfare. Furthermore, there have been suggestions that such research should be stopped because of the possibility of accidental or deliberate release of these viruses into the community. The first act of censoring scientific data may set the precedence of interfering with academic freedom in science.
Further analysis of the unpublished data suggested that the mutant virus is not as lethal as the wild type and that the mutant was not so readily air transmissible as initially perceived. Based on these findings, the NSABB has changed its position and recommended that the studies should be published in full.Citation150 In the study by Imai et al.,Citation146 reassortant influenza viruses containing the HA from an H5N1 virus and seven gene segments from an A(H1N1)pdm09 virus were created. One of the mutant viruses, which contained four mutations in HA (N154D, N220K, Q222L and T315I), was able to transmit more efficiently via the droplet route between ferrets. The mutations N154D, N220K and Q222L were thought to be responsible for enhanced binding to α2,6 SA, while T315I was thought to improve fusion between the viral envelope and the intracellular membranes. In the study by Herfst et al.,Citation147 the mutant H5N1 virus was generated by site-directed mutagenesis and serial passage in ferrets. H103Y, T156A, Q222L, G224S of HA and E627K of PB2 were present in all viruses that were isolated from ferrets that acquired the H5N1 virus via the non-contact route. It is clear from these two studies that some of the initial fears were not justified. First, many commentaries have used the term ‘airborne’.Citation151 However, both studies did not demonstrate that the viruses were transmitted via the airborne route but rather via non-contact routes (airborne or droplet) because the inoculated and naive ferrets were placed in cages that were ≤10 cm apart. Airborne transmission has been defined as transmission via droplet nuclei ≤5 µm, which may be transmitted over long distances >1 m.Citation152 Second, the mutant viruses were not highly pathogenic. No deaths occurred among the ferrets that were infected via the non-contact route.
Conclusion
In this review, we have reported a case of human infection due to H5N1 virus subclade 2.3.2.1 in South China and provided an update on the H5N1 virus related to human infection. Despite aggressive control measures in this area, sporadic human H5N1 infections still occur, highlighting the need for high vigilance, especially when encountering patients who have poultry contact or have visited a poultry market. Indonesia still has the highest number of cumulative human cases worldwide, although Egypt has the highest number of human H5N1 cases reported since 2009. Differential binding affinity to α2,3 SA and α2,6 SA by the H5N1 virus may explain the rapid spread of H5N1 virus in Egypt. Despite worldwide surveillance and aggressive strategies to eliminate the H5N1 virus, the virus continues to cause fatal outbreaks in both the avian and human population. The viruses have rapidly evolved, generating many subclades with potentially enhanced virulence or transmissibility. This evolution may have been driven by increased infection among non-chicken poultry, which often have mild manifestations,Citation153 increased mixing of viruses from different areas facilitated by migratory birds,Citation154 and mass H5N1 virus vaccination among poultries.Citation155 Tremendous progress has been made towards understanding the human H5N1 virus infection, but the mortality rate in human H5N1 cases has remained near 60% since 1997,Citation1,Citation8 although the mortality rate is as low as 30% in Egypt. Due to the lack of resources, early detection is limited in developing countries, and even in affluent countries, detection may be delayed due to the lack of awareness. Analysis of the viral genome allows the scientific community to identify virulence determinants, but such knowledge has had little clinical impact yet. Despite antiviral treatment, many patients still succumbed to viral disease. Since the introduction of neuraminidase inhibitors more than 10 years ago, no new antivirals that are active against the influenza virus have been approved. Many preventive strategies, especially vaccinations, are effective in limiting H5N1 virus transmission in poultry, but the benefit of vaccination can be impaired by the low uptake rate in backyard farms and the lack of cross-protection between different clades or subclades.
We are grateful to the generous support of Carol Yu, Richard Yu, Hui Hoy and Hui Ming for the genomic sequencing platform. This work was partly supported by the Hong Kong Special Administrative Region Research Fund for the Control of Infectious Diseases of the Health, Welfare, and Food Bureau; the Providence Foundation Limited, in memory of the late Lui Hac Minh; and the Consultancy Service for Enhancing Laboratory Surveillance of Emerging Infectious Disease for the Hong Kong Special Administrative Region Department of Health.
References
- Yuen KY, Chan PK, Peiris M et al. Clinical features and rapid viral diagnosis of human disease associated with avian influenza A H5N1 virus. Lancet1998; 351: 467–471.
- Cheng VC, Chan JF, Wen X et al. Infection of immunocompromised patients by avian H9N2 influenza A virus. J Infect2011; 62: 394–399.
- Banks J, Speidel E, Alexander DJ. Characterisation of an avian influenza A virus isolated from a human—is an intermediate host necessary for the emergence of pandemic influenza viruses? Arch Virol1998; 143: 781–787.
- Editorial team. Avian influenza A/(H7N2) outbreak in the United Kingdom. Euro Surveill2007; 12: E070531–E070532.
- Tweed SA, Skowronski DM, David ST et al. Human illness from avian influenza H7N3, British Columbia. Emerg Infect Dis2004; 10: 2196–2199.
- Arzey GG, Kirkland PD, Arzey KE et al. Influenza virus A (H10N7) in chickens and poultry abattoir workers, Australia. Emerg Infect Dis2012; 18: 814–816.
- Cheng VC, To KK, Tse H, Hung IF, Yuen KY. Two years after pandemic influenza A/2009/H1N1: what have we learned? Clin Microbiol Rev2012; 25: 223–263.
- World Health Organization. Cumulative number of confirmed human cases for avian influenza A(H5N1) reported to WHO, 2003–2012. Geneva: WHO, 2012. Available at http://www.who.int/influenza/human_animal_interface/EN_GIP_20120529CumulativeNumberH5N1cases.pdf (accessed 6 June 2012).
- Abdel-Ghafar AN, Chotpitayasunondh T, Gao Z et al. Update on avian influenza A (H5N1) virus infection in humans. N Engl J Med2008; 358: 261–273.
- World Health Organization. Antigenic and genetic characteristics of zoonotic influenza viruses and development of candidate vaccine viruses for pandemic preparedness. Geneva: WHO, 2012. Available at http://www.who.int/influenza/vaccines/virus/201202_h5_h9_vaccinevirusupdate.pdf (accessed 6 June 2012).
- World Health Organization. Antigenic and genetic characteristics of zoonotic influenza viruses and development of candidate vaccine viruses for pandemic preparedness. Geneva: WHO, 2011. Available at http://www.who.int/influenza/resources/documents/2011_09_h5_h9_vaccinevirusupdate.pdf (accessed 6 June 2012).
- World Health Organization. Updated unified nomenclature system for the highly pathogenic H5N1 avian influenza viruses. Geneva: WHO, 2011. Available at http://www.who.int/influenza/gisrs_laboratory/h5n1_nomenclature/en/index.html (accessed 6 June 2012).
- Fiebig L, Soyka J, Buda S, Buchholz U, Dehnert M, Haas W. Avian influenza A(H5N1) in humans: new insights from a line list of World Health Organization confirmed cases, September 2006 to August 2010. Eur Surveill2011; 16: pii: 19941.
- Kayali G, El-Shesheny R, Kutkat MA et al. Continuing threat of influenza (H5N1) virus circulation in Egypt. Emerg Infect Dis2011; 17: 2306–2308.
- Wang TT, Parides MK, Palese P. Seroevidence for H5N1 influenza infections in humans: meta-analysis. Science2012; 335: 1463.
- Murray EJ, Morse SS. Seasonal oscillation of human infection with influenza A/H5N1 in Egypt and Indonesia. PLoS ONE2011; 6: e24042.
- To KK, Wong SS, Li IW et al. Concurrent comparison of epidemiology, clinical presentation and outcome between adult patients suffering from the pandemic influenza A (H1N1) 2009 virus and the seasonal influenza A virus infection. Postgrad Med J2010; 86: 515–521.
- Zhang AJ, To KK, Tse H et al. High incidence of severe influenza among individuals over 50 years of age. Clin Vaccine Immunol2011; 18: 1918–1924.
- Rutigliano JA, Morris MY, Yue W et al. Protective memory responses are modulated by priming events prior to challenge. J Virol2010; 84: 1047–1056.
- Liem NT, Tung CV, Hien ND et al. Clinical features of human influenza A (H5N1) infection in Vietnam: 2004–2006. Clin Infect Dis2009; 48: 1639–1646.
- Wan XF, Dong L, Lan Y et al. Indications that live poultry markets are a major source of human H5N1 influenza virus infection in China. J Virol2011; 85: 13432–13438.
- Zhou L, Liao Q, Dong L et al. Risk factors for human illness with avian influenza A (H5N1) virus infection in China. J Infect Dis2009; 199: 1726–1734.
- Indriani R, Samaan G, Gultom A et al. Environmental sampling for avian influenza virus A (H5N1) in live-bird markets, Indonesia. Emerg Infect Dis2010; 16: 1889–1895.
- Wang H, Feng Z, Shu Y et al. Probable limited person-to-person transmission of highly pathogenic avian influenza A (H5N1) virus in China. Lancet2008; 371: 1427–1434.
- Gambotto A, Barratt-Boyes SM, de Jong MD, Neumann G, Kawaoka Y. Human infection with highly pathogenic H5N1 influenza virus. Lancet2008; 371: 1464–1475.
- Shinya K, Makino A, Tanaka H et al. Systemic dissemination of H5N1 influenza A viruses in ferrets and hamsters after direct intragastric inoculation. J Virol2011; 85: 4673–4678.
- Harder TC, Teuffert J, Starick E et al. Highly pathogenic avian influenza virus (H5N1) in frozen duck carcasses, Germany, 2007. Emerg Infect Dis2009; 15: 272–279.
- Imai M, Kawaoka Y. The role of receptor binding specificity in interspecies transmission of influenza viruses. Curr Opin Virol2012; 2: 160–167.
- Gabriel G, Klingel K, Otte A et al. Differential use of importin-alpha isoforms governs cell tropism and host adaptation of influenza virus. Nat Commun2011; 2: 156.
- Zimmermann P, Manz B, Haller O, Schwemmle M, Kochs G. The viral nucleoprotein determines Mx sensitivity of influenza A viruses. J Virol2011; 85: 8133–8140.
- Miotto O, Heiny AT, Albrecht R et al. Complete-proteome mapping of human influenza A adaptive mutations: implications for human transmissibility of zoonotic strains. PLoS ONE2010; 5: e9025.
- Tong S, Li Y, Rivailler P et al. A distinct lineage of influenza A virus from bats. Proc Natl Acad Sci USA2012; 109: 4269–4274.
- Watanabe Y, Ibrahim MS, Suzuki Y, Ikuta K. The changing nature of avian influenza A virus (H5N1). Trends Microbiol2012; 20: 11–20.
- Horimoto T, Kawaoka Y. Influenza: lessons from past pandemics, warnings from current incidents. Nat Rev Microbiol2005; 3: 591–600.
- WHO/OIE/FAO H5N1 Evolution Working Group. Continued evolution of highly pathogenic avian influenza A (H5N1): updated nomenclature. Influenza Other Respi Viruses2012; 6: 1–5.
- Buchy P, Fourment M, Mardy S et al. Molecular epidemiology of clade 1 influenza A viruses (H5N1), southern Indochina peninsula, 2004–2007. Emerg Infect Dis2009; 15: 1641–1644.
- Dung Nguyen T, Vinh Nguyen T, Vijaykrishna D et al. Multiple sublineages of influenza A virus (H5N1), Vietnam, 2005–2007. Emerg Infect Dis2008; 14: 632–636.
- Suwannakarn K, Amonsin A, Sasipreeyajan J et al. Molecular evolution of H5N1 in Thailand between 2004 and 2008. Infect Genet Evol2009; 9: 896–902.
- Buchy P, Mardy S, Vong S et al. Influenza A/H5N1 virus infection in humans in Cambodia. J Clin Virol2007; 39: 164–168.
- Le QM, Ito M, Muramoto Y et al. Pathogenicity of highly pathogenic avian H5N1 influenza A viruses isolated from humans between 2003 and 2008 in northern Vietnam. J Gen Virol2010; 91( Pt 10): 2485–2490.
- Schmolke M, Manicassamy B, Pena L et al. Differential contribution of PB1-F2 to the virulence of highly pathogenic H5N1 influenza A virus in mammalian and avian species. PLoS Pathog2011; 7: e1002186.
- Conenello GM, Zamarin D, Perrone LA, Tumpey T, Palese P. A single mutation in the PB1-F2 of H5N1 (HK/97) and 1918 influenza A viruses contributes to increased virulence. PLoS Pathog2007; 3: 1414–1421.
- Govorkova EA, Ilyushina NA, Boltz DA, Douglas A, Yilmaz N, Webster RG. Efficacy of oseltamivir therapy in ferrets inoculated with different clades of H5N1 influenza virus. Antimicrob Agents Chemother2007; 51: 1414–1424.
- Nidom CA, Yamada S, Nidom RV et al. Genetic characterization of H5N1 influenza viruses isolated from chickens in Indonesia in 2010. Virus Genes2012; 44: 459–465.
- Liu J, Xiao H, Lei F et al. Highly pathogenic H5N1 influenza virus infection in migratory birds. Science2005; 309: 1206.
- Li Y, Shi J, Zhong G et al. Continued evolution of H5N1 influenza viruses in wild birds, domestic poultry, and humans in China from 2004 to 2009. J Virol2010; 84: 8389–8397.
- Watanabe Y, Ibrahim MS, Ellakany HF et al. Acquisition of human-type receptor binding specificity by new H5N1 influenza virus sublineages during their emergence in birds in Egypt. PLoS Pathog2011; 7: e1002068.
- Chen H, Li Y, Li Z et al. Properties and dissemination of H5N1 viruses isolated during an influenza outbreak in migratory waterfowl in western China. J Virol2006; 80: 5976–5983.
- Neumann G, Chen H, Gao GF, Shu Y, Kawaoka Y. H5N1 influenza viruses: outbreaks and biological properties. Cell Res2010; 20: 51–61.
- Le MT, Wertheim HF, Nguyen HD, et al. Influenza A H5N1 clade 2.3.4 virus with a different antiviral susceptibility profile replaced clade 1 virus in humans in northern Vietnam. PLoS ONE2008; 3: e3339.
- Zhou JF, Zou SM, Li Z et al. Identification of dual receptor-binding specific strains of human H5N1 Viruses in China. Biomed Environ Sci2012; 25: 104–108.
- Chen H, Smith GJ, Li KS et al. Establishment of multiple sublineages of H5N1 influenza virus in Asia: implications for pandemic control. Proc Natl Acad Sci USA2006; 103: 2845–2850.
- Sakoda Y, Ito H, Uchida Y et al. Reintroduction of H5N1 highly pathogenic avian influenza virus by migratory water birds, causing poultry outbreaks in 2010–2011 winter season in Japan. J Gen Virol2012; 93( Pt 3): 541–550.
- Islam MR, Haque ME, Giasuddin M et al. New introduction of clade 2.3.2.1 avian influenza virus (H5N1) into Bangladesh. Transbound Emerg Dis 2011 Dec 28. doi:10.1111/j.1865–1682.2011.01297.x.
- WHO/FAO/OIE H5N1 Evolution Working Group. FAO-OIE-WHO technical update: current evolution of avian influenza H5N1 viruses. Geneva: WHO, 2012. Available at http://www.who.int/influenza/human_animal_interface/tripartite_notes_H5N1.pdf (accessed 6 June 2012).
- Davis CT, Balish AL, O'Neill E et al. Detection and characterization of clade 7 high pathogenicity avian influenza H5N1 viruses in chickens seized at ports of entry and live poultry markets in Vietnam. Avian Dis2010; 54( 1 Suppl): 307–312.
- Bao Y, Bolotov P, Dernovoy D et al. The influenza virus resource at the National Center for Biotechnology Information. J Virol2008; 82: 596–601.
- Auewarakul P, Suptawiwat O, Kongchanagul A et al. An avian influenza H5N1 virus that binds to a human-type receptor. J Virol2007; 81: 9950–9955.
- Medina RA, Garcia-Sastre A. Influenza A viruses: new research developments. Nat Rev Microbiol2011; 9: 590–603.
- Gao Y, Zhang Y, Shinya K et al. Identification of amino acids in HA and PB2 critical for the transmission of H5N1 avian influenza viruses in a mammalian host. PLoS Pathog2009; 5: e1000709.
- Stevens J, Blixt O, Chen LM, Donis RO, Paulson JC, Wilson IA. Recent avian H5N1 viruses exhibit increased propensity for acquiring human receptor specificity. J Mol Biol2008; 381: 1382–1394.
- Yamada S, Suzuki Y, Suzuki T et al. Haemagglutinin mutations responsible for the binding of H5N1 influenza A viruses to human-type receptors. Nature2006; 444: 378–382.
- Yamada S, Hatta M, Staker BL et al. Biological and structural characterization of a host-adapting amino acid in influenza virus. PLoS Pathog2010; 6: e1001034.
- Ungchusak K, Auewarakul P, Dowell SF et al. Probable person-to-person transmission of avian influenza A (H5N1). N Engl J Med2005; 352: 333–340.
- Huai Y, Xiang N, Zhou L et al. Incubation period for human cases of avian influenza A (H5N1) infection, China. Emerg Infect Dis2008; 14: 1819–1821.
- Yu H, Gao Z, Feng Z et al. Clinical characteristics of 26 human cases of highly pathogenic avian influenza A (H5N1) virus infection in China. PLoS ONE2008; 3: e2985.
- Shinya K, Ebina M, Yamada S, Ono M, Kasai N, Kawaoka Y. Avian flu: influenza virus receptors in the human airway. Nature2006; 440: 435–436.
- Nicholls JM, Chan MC, Chan WY et al. Tropism of avian influenza A (H5N1) in the upper and lower respiratory tract. Nat Med2007; 13: 147–149.
- van Riel D, den Bakker MA, Leijten LM et al. Seasonal and pandemic human influenza viruses attach better to human upper respiratory tract epithelium than avian influenza viruses. Am J Pathol2010; 176: 1614–1618.
- Shinya K, Hatta M, Yamada S et al. Characterization of a human H5N1 influenza A virus isolated in 2003. J Virol2005; 79: 9926–9932.
- Chandrasekaran A, Srinivasan A, Raman R et al. Glycan topology determines human adaptation of avian H5N1 virus hemagglutinin. Nat Biotechnol2008; 26: 107–113.
- To KK, Hung IF, Li IW et al. Delayed clearance of viral load and marked cytokine activation in severe cases of pandemic H1N1 2009 influenza virus infection. Clin Infect Dis2010; 50: 850–859.
- Schrauwen EJ, Herfst S, Leijten LM et al. The multi basic cleavage site in H5N1 virus is critical for systemic spread along the olfactory and hematogenous routes in ferrets. J Virol2012; 86: 3975–3984.
- Zhang Z, Zhang J, Huang K et al. Systemic infection of avian influenza A virus H5N1 subtype in humans. Hum Pathol2009; 40: 735–739.
- Gu J, Xie Z, Gao Z et al. H5N1 infection of the respiratory tract and beyond: a molecular pathology study. Lancet2007; 370: 1137–1145.
- Zhou JJ, Fang DY, Fu J et al. Infection and replication of avian influenza H5N1 virus in an infected human. Virus Genes2009; 39: 76–80.
- de Jong MD, Bach VC, Phan TQ et al. Fatal avian influenza A (H5N1) in a child presenting with diarrhea followed by coma. N Engl J Med2005; 352: 686–691.
- Li IW, Chan KH, To KW et al. Differential susceptibility of different cell lines to swine-origin influenza A H1N1, seasonal human influenza A H1N1, and avian influenza A H5N1 viruses. J Clin Virol2009; 46: 325–330.
- Shinya K, Makino A, Hatta M et al. Subclinical brain injury caused by H5N1 influenza virus infection. J Virol2011; 85: 5202–5207.
- Jang H, Boltz D, Sturm-Ramirez K et al. Highly pathogenic H5N1 influenza virus can enter the central nervous system and induce neuroinflammation and neurodegeneration. Proc Natl Acad Sci USA2009; 106: 14063–14068.
- Woo PC, Tung ET, Chan KH, Lau CC, Lau SK, Yuen KY. Cytokine profiles induced by the novel swine-origin influenza A/H1N1 virus: implications for treatment strategies. J Infect Dis2010; 201: 346–353.
- Cilloniz C, Shinya K, Peng X et al. Lethal influenza virus infection in macaques is associated with early dysregulation of inflammatory related genes. PLoS Pathog2009; 5: e1000604.
- Zeng H, Pappas C, Belser JA et al. Human pulmonary microvascular endothelial cells support productive replication of highly pathogenic avian influenza viruses: possible involvement in the pathogenesis of human H5N1 virus infection. J Virol2012; 86: 667–678.
- Schmolke M, Viemann D, Roth J, Ludwig S. Essential impact of NF-kappaB signaling on the H5N1 influenza A virus-induced transcriptome. J Immunol2009; 183: 5180–5189.
- Li C, Bankhead A 3rd, Eisfeld AJ et al. Host regulatory network response to infection with highly pathogenic H5N1 avian influenza virus. J Virol2011; 85: 10955–10967.
- Ramos I, Bernal-Rubio D, Durham N et al. Effects of receptor binding specificity of avian influenza virus on the human innate immune response. J Virol2011; 85: 4421–4431.
- Cilloniz C, Pantin-Jackwood MJ, Ni C et al. Lethal dissemination of H5N1 influenza virus is associated with dysregulation of inflammation and lipoxin signaling in a mouse model of infection. J Virol2010; 84: 7613–7624.
- Soepandi PZ, Burhan E, Mangunnegoro H et al. Clinical course of avian influenza A(H5N1) in patients at the Persahabatan Hospital, Jakarta, Indonesia, 2005–2008. Chest2010; 138: 665–673.
- Brundage JF. Interactions between influenza and bacterial respiratory pathogens: implications for pandemic preparedness. Lancet Infect Dis2006; 6: 303–312.
- To KK, Chan KH, Fung YF, Yuen KY, Ho PL. Azithromycin treatment failure in macrolide-resistant Mycoplasma pneumoniae pneumonia. Eur Respir J2010; 36: 969–971.
- de Jong MD, Simmons CP, Thanh TT et al. Fatal outcome of human influenza A (H5N1) is associated with high viral load and hypercytokinemia. Nat Med2006; 12: 1203–1207.
- Chen H, Wen X, To KK et al. Quasispecies of the D225G substitution in the hemagglutinin of pandemic influenza A(H1N1) 2009 virus from patients with severe disease in Hong Kong, China. J Infect Dis2010; 201: 1517–1521.
- Zheng B, Chan KH, Zhang AJ et al. D225G mutation in hemagglutinin of pandemic influenza H1N1 (2009) virus enhances virulence in mice. Exp Biol Med (Maywood)2010; 235: 981–988.
- Zhou J, To KK, Dong H et al. A functional variation in CD55 increases the severity of 2009 pandemic H1N1 influenza A virus infection. J Infect Dis2012; 206: 495–503
- Sakai-Tagawa Y, Ozawa M, Tamura D et al. Sensitivity of influenza rapid diagnostic tests to H5N1 and 2009 pandemic H1N1 viruses. J Clin Microbiol2010; 48: 2872–2877.
- Chen W, He B, Li C et al. Real-time RT-PCR for H5N1 avian influenza A virus detection. J Med Virol2007; 56( Pt 5): 603–607.
- Rowe T, Abernathy RA, Hu-Primmer J et al. Detection of antibody to avian influenza A (H5N1) virus in human serum by using a combination of serologic assays. J Clin Microbiol1999; 37: 937–943.
- Buchy P, Vong S, Chu S et al. Kinetics of neutralizing antibodies in patients naturally infected by H5N1 virus. PLoS ONE2010; 5: e10864.
- Lan Y, Zhang Y, Dong L et al. A comprehensive surveillance of adamantane resistance among human influenza A virus isolated from mainland China between 1956 and 2009. Antivir Ther2010; 15: 853–859.
- Cheung CL, Rayner JM, Smith GJ et al. Distribution of amantadine-resistant H5N1 avian influenza variants in Asia. J Infect Dis2006; 193: 1626–1629.
- Pielak RM, Schnell JR, Chou JJ. Mechanism of drug inhibition and drug resistance of influenza A M2 channel. Proc Natl Acad Sci USA2009; 106: 7379–7384.
- Adisasmito W, Chan PK, Lee N et al. Effectiveness of antiviral treatment in human influenza A(H5N1) infections: analysis of a Global Patient Registry. J Infect Dis2010; 202: 1154–1160.
- Stittelaar KJ, Tisdale M, van Amerongen G et al. Evaluation of intravenous zanamivir against experimental influenza A (H5N1) virus infection in cynomolgus macaques. Antiviral Res2008; 80: 225–228.
- Kiso M, Kubo S, Ozawa M et al. Efficacy of the new neuraminidase inhibitor CS-8958 against H5N1 influenza viruses. PLoS Pathog2010; 6: e1000786.
- Boltz DA, Ilyushina NA, Arnold CS, Babu YS, Webster RG, Govorkova EA. Intramuscularly administered neuraminidase inhibitor peramivir is effective against lethal H5N1 influenza virus in mice. Antivir Res2008; 80: 150–157.
- McKimm-Breschkin JL, Selleck PW, Usman TB, Johnson MA. Reduced sensitivity of influenza A (H5N1) to oseltamivir. Emerg Infect Dis2007; 13: 1354–1357.
- Naughtin M, Dyason JC, Mardy S, Sorn S, von Itzstein M, Buchy P. Neuraminidase inhibitor sensitivity and receptor-binding specificity of Cambodian clade 1 highly pathogenic H5N1 influenza virus. Antimicrob Agents Chemother2011; 55: 2004–2010.
- de Jong MD, Tran TT, Truong HK et al. Oseltamivir resistance during treatment of influenza A (H5N1) infection. N Engl J Med2005; 353: 2667–2672.
- Kiso M, Ozawa M, Le MT et al. Effect of an asparagine-to-serine mutation at position 294 in neuraminidase on the pathogenicity of highly pathogenic H5N1 influenza A virus. J Virol2011; 85: 4667–4672.
- Boltz DA, Douangngeun B, Phommachanh P et al. Emergence of H5N1 avian influenza viruses with reduced sensitivity to neuraminidase inhibitors and novel reassortants in Lao People's Democratic Republic. J Gen Virol2010; 91( Pt 4): 949–959.
- Collins PJ, Haire LF, Lin YP et al. Crystal structures of oseltamivir-resistant influenza virus neuraminidase mutants. Nature2008; 453: 1258–1261.
- Zhao G, Zhong L, Lu X, et al. Characterisation of a highly pathogenic H5N1 clade 2.3.2 influenza virus isolated from swans in Shanghai, China. Virus Genes2012; 44: 55–62.
- Jefferson T, Jones MA, Doshi P et al. Neuraminidase inhibitors for preventing and treating influenza in healthy adults and children. Cochrane Database Syst Rev2012; 1: CD008965.
- Kao RY, Yang D, Lau LS et al. Identification of influenza A nucleoprotein as an antiviral target. Nat Biotechnol2010; 28: 600–605.
- Kumaki Y, Day CW, Smee DF, Morrey JD, Barnard DL. In vitro and in vivo efficacy of fluorodeoxycytidine analogs against highly pathogenic avian influenza H5N1, seasonal, and pandemic H1N1 virus infections. Antiviral Res2011; 92: 329–340.
- Kiso M, Takahashi K, Sakai-Tagawa Y et al. T-705 (favipiravir) activity against lethal H5N1 influenza A viruses. Proc Natl Acad Sci USA2010; 107: 882–887.
- Triana-Baltzer GB, Gubareva LV, Klimov AI et al. Inhibition of neuraminidase inhibitor-resistant influenza virus by DAS181, a novel sialidase fusion protein. PLoS ONE2009; 4: e7838.
- Belser JA, Lu X, Szretter KJ et al. DAS181, a novel sialidase fusion protein, protects mice from lethal avian influenza H5N1 virus infection. J Infect Dis2007; 196: 1493–1499.
- Nicol MQ, Ligertwood Y, Bacon MN, Dutia BM, Nash AA. A novel family of peptides with potent activity against influenza A viruses. J Gen Virol2012; 93( Pt 5): 980–986.
- Zheng BJ, Chan KW, Lin YP et al. Delayed antiviral plus immunomodulator treatment still reduces mortality in mice infected by high inoculum of influenza A/H5N1 virus. Proc Natl Acad Sci USA2008; 105: 8091–8096.
- Henter JI, Chow CB, Leung CW, Lau YL. Cytotoxic therapy for severe avian influenza A (H5N1) infection. Lancet2006; 367: 870–873.
- Henter JI, Palmkvist-Kaijser K, Holzgraefe B, Bryceson YT, Palmer K. Cytotoxic therapy for severe swine flu A/H1N1. Lancet2010; 376: 2116.
- Van Hoeven N, Belser JA, Szretter KJ et al. Pathogenesis of 1918 pandemic and H5N1 influenza virus infections in a guinea pig model: antiviral potential of exogenous alpha interferon to reduce virus shedding. J Virol2009; 83: 2851–2861.
- Haasbach E, Pauli EK, Spranger R et al. Antiviral activity of the proteasome inhibitor VL-01 against influenza A viruses. Antiviral Res2011; 91: 304–313.
- Hung IF, To KK, Lee CK et al. Convalescent plasma treatment reduced mortality in patients with severe pandemic influenza A (H1N1) 2009 virus infection. Clin Infect Dis2011; 52: 447–456.
- Prabhu N, Prabakaran M, Ho HT et al. Monoclonal antibodies against the fusion peptide of hemagglutinin protect mice from lethal influenza A virus H5N1 infection. J Virol2009; 83: 2553–2562.
- Yupiana Y, de Vlas SJ, Adnan NM, Richardus JH. Risk factors of poultry outbreaks and human cases of H5N1 avian influenza virus infection in West Java Province, Indonesia. Int J Infect Dis2010; 14: e800–e805.
- Guan Y, Chen H, Li K et al. A model to control the epidemic of H5N1 influenza at the source. BMC Infect Dis2007; 7: 132.
- Henning J, Wibawa H, Morton J, Usman TB, Junaidi A, Meers J. Scavenging ducks and transmission of highly pathogenic avian influenza, Java, Indonesia. Emerg Infect Dis2010; 16: 1244–1250.
- Paul M, Wongnarkpet S, Gasqui P et al. Risk factors for highly pathogenic avian influenza (HPAI) H5N1 infection in backyard chicken farms, Thailand. Acta Trop2011; 118: 209–216.
- Henning KA, Henning J, Morton J, Long NT, Ha NT, Meers J. Farm- and flock-level risk factors associated with highly pathogenic avian influenza outbreaks on small holder duck and chicken farms in the Mekong Delta of Viet Nam. Prev Vet Med2009; 91: 179–188.
- Wu WL, Chen Y, Wang P et al. Antigenic profile of avian H5N1 viruses in Asia from 2002 to 2007. J Virol2008; 82: 1798–1807.
- Normile D. Avian influenza. Warning of H5N1 resurgence surprises community. Science2011; 333: 1369.
- Sakoda Y, Ito H, Uchida Y et al. Reintroduction of H5N1 highly pathogenic avian influenza virus by migratory water birds, causing poultry outbreaks in the 2010–2011 winter season in Japan. J Gen Virol2012; 93( Pt 3): 541–550.
- Ehrlich HJ, Muller M, Oh HM et al. A clinical trial of a whole-virus H5N1 vaccine derived from cell culture. N Engl J Med2008; 358: 2573–2584.
- Khurana S, Wu J, Verma N et al. H5N1 virus-like particle vaccine elicits cross-reactive neutralizing antibodies that preferentially bind to the oligomeric form of influenza virus hemagglutinin in humans. J Virol2011; 85: 10945–10954.
- Langley JM, Risi G, Caldwell M et al. Dose-sparing H5N1 A/Indonesia/05/2005 pre-pandemic influenza vaccine in adults and elderly adults: a phase III, placebo-controlled, randomized study. J Infect Dis2011; 203: 1729–1738.
- Galli G, Hancock K, Hoschler K et al. Fast rise of broadly cross-reactive antibodies after boosting long-lived human memory B cells primed by an MF59 adjuvanted prepandemic vaccine. Proc Natl Acad Sci USA2009; 106: 7962–7967.
- Patel SM, Atmar RL, El Sahly HM, Cate TR, Keitel WA. A phase I evaluation of inactivated influenza A/H5N1 vaccine administered by the intradermal or the intramuscular route. Vaccine2010; 28: 3025–3029.
- Stephenson I, Nicholson KG, Hoschler K et al. Antigenically distinct MF59-adjuvanted vaccine to boost immunity to H5N1. N Engl J Med2008; 359: 1631–1633.
- Lyall J, Irvine RM, Sherman A et al. Suppression of avian influenza transmission in genetically modified chickens. Science2011; 331: 223–226.
- Steel J, Lowen AC, Mubareka S, Palese P. Transmission of influenza virus in a mammalian host is increased by PB2 amino acids 627K or 627E/701N. PLoS Pathog2009; 5: e1000252.
- Belser JA, Katz JM, Tumpey TM. The ferret as a model organism to study influenza A virus infection. Dis Model Mech2011; 4: 575–579.
- Gustin KM, Belser JA, Wadford DA, et al. Influenza virus aerosol exposure and analytical system for ferrets. Proc Natl Acad Sci USA2011; 108: 8432–8437.
- Chen LM, Blixt O, Stevens J et al. In vitro evolution of H5N1 avian influenza virus toward human-type receptor specificity. Virology2012; 422: 105–113.
- Imai M, Watanabe T, Hatta M et al. Experimental adaptation of an influenza H5 HA confers respiratory droplet transmission to a reassortant H5 HA/H1N1 virus in ferrets. Nature2012; 486: 420–428.
- Herfst S, Schrauwen EJ, Linster M et al. Airborne transmission of influenza A/H5N1 virus between ferrets. Science2012; 336: 1534–1541.
- World Health Organization. Public health, influenza experts agree H5N1 research critical, but extend delay. Geneva: WHO, 2012. Available at: http://www.who.int/mediacentre/news/releases/2012/h5n1_research_20120217/en/index.html (accessed 6 June 2012).
- Palese P, Wang TT. H5N1 influenza viruses: Facts, not fear. Proc Natl Acad Sci USA2012; 109: 2211–2213.
- Cohen J, Malakoff D. Avian influenza. On second thought, flu papers get go-ahead. Science2012; 336: 19–20.
- Enserink M. Infectious diseases. Controversial studies give a deadly flu virus wings. Science2011; 334: 1192–1193.
- Siegel JD, Rhinehart E, Jackson M, Chiarello L, Practices HCIC. 2007 guideline for isolation precautions: preventing transmission of infectious agents in health care settings. Am J Infect Control2007; 35: S65–S164.
- Carrel M, Wan XF, Nguyen T, Emch M. Genetic Variation of highly pathogenic H5N1 avian influenza viruses in vietnam shows both species-specific and spatiotemporal associations. Avian Dis2011; 55: 659–666.
- Fusaro A, Nelson MI, Joannis T et al. Evolutionary dynamics of multiple sublineages of H5N1 influenza viruses in Nigeria from 2006 to 2008. J Virol2010; 84: 3239–3247.
- Cattoli G, Fusaro A, Monne I et al. Evidence for differing evolutionary dynamics of A/H5N1 viruses among countries applying or not applying avian influenza vaccination in poultry. Vaccine2011; 29: 9368–9375.