
Dear Editor,
Emerging and reemerging viruses that cause respiratory infectious diseases, such as severe acute respiratory syndromes coronavirus (SARS-CoV), Middle East respiratory syndrome coronavirus (MERS-CoV), and influenza viruses, are a significant threat to public health worldwide. Although vaccines are the most effective strategy to prevent viral infection, vaccine development is a long process and may be effective only against the corresponding virus. Therefore, it is essential to develop antiviral agents for intranasal administration as nonspecific prophylaxis against infection by emerging or reemerging respiratory viruses with epidemic or pandemic potential.
Cholera toxin (CT), which acts as a mucosal adjuvant to stimulate mucosal and systemic immune responses, has the potential for intranasal application as an immunotherapeutic against infection by respiratory viruses.Citation1 However, CT can exacerbate lung pathology after influenza virus infection through the induction of IL-17, a potent proinflammatory cytokine, because co-administration of an IL-17RA neutralizing antibody with CT attenuates lung pathology and increases protection against influenza virus infection.Citation2 Therefore, we hypothesized that intranasal application of a mucosal stimulant that induces antiviral cytokines, except IL-17, may be effective against influenza virus infection.
In this regard, polyethyleneimine (PEI), a mucosal adjuvant, exhibits stronger mucosal adjuvanticity but induces much lower IL-17 expression than cholera toxin subunit B (CTB).Citation3 Therefore, for the first time, we tested PEI for its potential protective effects against influenza virus infection. Ten-week-old specific-pathogen-free (SPF) female Balb/c mice were anesthetized with pelltobarbitalum natricum. Then, 50 μL of PBS containing 20 μg of 25 kD linear PEI (Polysciences, Warrington, PA) or 2.5 μg of CTB (Sigma-Aldrich, St Louis, MO, USA) or PBS alone as a control was intranasally administered twice at 24 and 48 h before challenge with 5 LD50 influenza virus H1N1 (A/PR/8/34). Mouse body weights were monitored every day, and those with greater than 25% loss of their initial body weight were euthanized as described.Citation4
As shown in , the body weight of the mice in the PEI-pretreated group remained stable from day 1 to day 5 after H1N1 challenge and then gradually decreased until day 11, for a total loss of 18%, before recovering. By contrast, the body weights of the mice in the CTB- and PBS-pretreated groups decreased significantly beginning on day 2 and reached losses of more than 25% by day 8 and 10, respectively, after H1N1 challenge. The final survival rate of the mice in the PEI-pretreated group was 60%, whereas that of the mice in the CTB- and PBS-pretreated groups was 0% (). We then examined the viral titers in mouse lungs as previously describedCitation5 and found that PEI significantly reduced lung viral titers on day 2 after H1N1 challenge, whereas the viral titers in the lungs of mice in the CTB- and PBS-pretreated groups showed no significant differences (). Subsequently, we examined lung sections stained with hematoxylin and eosin as previously described.Citation2 On day 2 after H1N1 infection, the pulmonary alveoli were relatively intact, and only a few inflammatory cells were observed in the lung tissues of mice in the PEI-pretreated group. However, the lungs of mice in the CTB- and PBS-pretreated groups were filled with abundant inflammatory cells (). These results suggest that intranasal application of PEI has efficacy in protecting mice from challenge by influenza virus H1N1.
Figure 1 Protective effects of PEI against H1N1 and H3N2 challenge. (A) Body weight loss after 5LD50 H1N1 challenge. (B) Survival rate of mice in the PEI-pretreated group (n=10), CTB-pretreated group (n=7), and PBS-pretreated group (n=8) after 5 LD50 H1N1 challenge. (C) Lung virus titer on day 2 after 5 LD50 H1N1 challenge. For panels A to C, *a significant difference (*P<0.05) was observed between the PEI-pretreated group and the CTB- as well as PBS-pretreated groups. (D) Examination of lung pathology in lung tissue sections stained with hematoxylin and eosin. Original magnification: × 200. (E) Body weight loss of mice after 5 LD50 H3N2 challenge. (F) Survival rate of mice after 5 LD50 H3N2 challenge. For panels E and F, *a significant difference (*P<0.05) and **very significant difference (**P<0.01) were observed between the PEI- and PBS-pretreated groups. Data are expressed as means±SD. All results were repeated and verified at least twice.
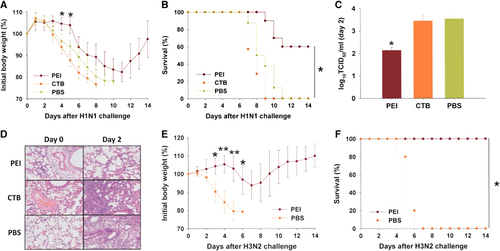
To determine the efficacy of PEI against another subtype of influenza virus, we pretreated mice with PBS containing PEI or PBS alone and challenged them with 5 LD50 influenza virus H3N2 (A/Guizhou/54/89). Mice in the PEI-pretreated group were fully protected against H3N2 challenge, showing no significant body weight loss () and a 100% survival rate, whereas those in the PBS group lost more than 25% of their body weight and showed a 0% survival rate on day 6 post-challenge (). Therefore, PEI-mediated protection against influenza virus infection is not subtype-specific.
To elucidate the mechanism of action of PEI, we examined the RNA levels of IFN-α4, IFN-β, IFN-γ, GM-CSF, IFITM3 and IL-17 in the lungs of mice pretreated with PEI, CTB, and PBS, respectively, before viral challenge using quantitative reverse transcription-PCR (qRT-PCR). As previously reported, some of these cytokines, such as interferon, GM-CSF and IFITM3, were effective in protecting against influenza infection.Citation6, Citation7, Citation8 As shown in Supplementary Figure S1, PEI induced a significantly higher RNA level of IFN-α4 than CTB or PBS. Furthermore, the RNA levels of GM-CSF and IFITM3 elicited by PEI were similar to those induced by CTB but much higher than those induced by PBS. The RNA level of IL-17 in mice pretreated with CTB, an IL-17-inducing adjuvant, was approximately 7- and 42-fold higher than that in mice pretreated with PEI and PBS, respectively. These results suggest that IFN-α4, GM-CSF, and IFITM3 are ‘good cytokines’ because they act as protective mediators against influenza virus infection, whereas IL-17 is a ‘bad cytokine’ that exacerbates pathology, primarily in the lung, after influenza infection.
In summary, we demonstrated that PEI, a mucosal stimulant for topical intranasal administration, is highly effective in preventing influenza virus infection. Compared to the bacteria-produced toxin CTB, the chemically synthesized polymer PEI is safer for mucosal application in humans. PEI has been tested in several clinical trials for gene delivery in vivo, demonstrating a good safety profile.Citation9 Moreover, its low cost of production and abundance makes PEI more suitable for urgent and widespread use during a time of influenza epidemic or pandemic.
Supplementary Figure S1
Download PDF (306.8 KB)Acknowledgments
We thank Dr Ze Chen at the Shanghai institute of biological products in China for providing influenza virus H3N2 (A/Guizhou/54/89). This work was supported by grants from the national nature science foundation of China (81590762 to Shibo Jiang, 81373456 to Lu Lu) and Shanghai Kai Star Project (16QA1400300) to Lu Lu.
Supplementary Information for this article can be found on the Emerging Microbes & Infections website (http://www.nature.com/emi)
- AdkinsI,HolubovaJ,KosovaMet al.Bacteria and their toxins tamed for immunotherapy.Curr Pharm Biotechnol2012; 13:1446–1473.
- GopalR,Rangel-MorenoJ,Fallert JuneckoBAet al.Mucosal pre-exposure to Th17-inducing adjuvants exacerbates pathology after influenza infection.Am J Pathol2014; 184:55–63.
- WegmannF,GartlanKH,HarandiAMet al.Polyethyleneimine is a potent mucosal adjuvant for viral glycoprotein antigens.Nat Biotechnol2012; 30:883–888.
- HeB,XiaS,YuFet al.Putative suppressing effect of IgG Fc-conjugated haemagglutinin (HA) stalk of influenza virus H7N9 on the neutralizing immunogenicity of Fc-conjugated HA head: implication for rational design of HA-based influenza vaccines.J Gen Virol2016; 97:327–333.
- HeB,ChangH,LiuZet al.Infection of influenza virus neuraminidase-vaccinated mice with homologous influenza virus leads to strong protection against heterologous influenza viruses.J Gen Virol2014; 95:2627–2637.
- StrayerDR,CarterWA,StouchBCet al.Protection from pulmonary tissue damage associated with infection of cynomolgus macaques by highly pathogenic avian influenza virus (H5N1) by low dose natural human IFN-alpha administered to the buccal mucosa.Antiviral Res2014; 110:175–180.
- HuangFF,BarnesPF,FengYet al.GM-CSF in the lung protects against lethal influenza infection.Am J Respir Crit Care Med2011; 184:259–268.
- EverittAR,ClareS,PertelTet al.IFITM3 restricts the morbidity and mortality associated with influenza.Nature2012; 484:519–523.
- NeubergP,KichlerA.Recent developments in nucleic acid delivery with polyethylenimines.Adv Genet2014; 88:263–288.