
Abstract
Re-emerging human adenovirus types 14 (Ad14) and 55 (Ad55) have caused severe respiratory diseases and even deaths during recent outbreaks. However, the seroprevalence of neutralizing antibodies (nAbs) in healthy adults, which may reflect previous circulation and help to predict potential outbreaks, remains unclear. In this study, we established micro-neutralizing (MN) assays on the basis of recombinant Ad14 and Ad55 reporter viruses, and we investigated serum nAbs in healthy blood donors from Southern China. We found that the overall seropositive rates were 24.8% and 22.4% for Ad14 and Ad55 nAbs, respectively. The seropositive rates were low in individuals younger than 20, and they gradually increased with age. Ad55-seropositive individuals tended to have high nAb titers (>1000), while low (72–200) and moderate (201–1000) nAb levels were frequently observed in Ad14-seropositive ones. Surprisingly, the seropositive rates and nAb levels were associated with the blood type but not the gender of the blood donors, with type AB individuals displaying higher seropositive rates and nAb levels. Interestingly, a significant positive correlation was observed between Ad14 and Ad55 seroprevalence, and higher titers of nAbs were detected in double-positive individuals compared to single-positive ones. These results clarified the human humoral immune responses against Ad14 and Ad55 and revealed a low level of herd immunity in some subpopulations, which emphasized the importance of monitoring these two highly virulent adenoviruses and reinforced the development of prophylactic vaccines.
Introduction
Human adenoviruses (AdVs) are non-enveloped, double-stranded DNA viruses belonging to Adenoviridae.Citation1 To date, more than 69 AdV genotypes, which were classified into seven species (A to G), were identified according to their genetic, biophysical or biochemical characteristics.Citation2 AdV infection causes a broad spectrum of diseases in humans, including pharyngoconjunctivitis, keratoconjunctivitis, gastroenteritis, myocarditis, acute hemorrhagic cystitis, meningoencephalitis and pneumonia, depending on the infection genotype.Citation3 Although the symptoms are usually mild or self-limited in immuno-competent adults, outbreaks of acute respiratory diseases (ARDs), such as community-acquired pneumonia (CAP), have been documented in particular populations, including newborns, school students and military recruits.Citation2 In immuno-comprised individuals such as patients with AIDS or individuals receiving transplants, AdV infection can lead to fatal pneumonia.Citation4
Recently, two members of human AdV species B, type 14 (Ad14) and 55 (Ad55), were reported to cause severe CAP in military and civilian populations.Citation5, Citation6, Citation7, Citation8, Citation9 Ad14 was first identified in an outbreak of ARDs in 1955 and then vanished for a long period.Citation10 A newly emergent variant of Ad14, which was designated Ad14p1, has caused several outbreaks of acute febrile respiratory illness globally since 2006.Citation11, Citation12, Citation13, Citation14, Citation15 High rates of severe illness and even deaths were reported following these outbreaks.Citation13, Citation14, Citation15 Ad55, an inter-typic recombinant of Ad11 and Ad14, emerged since 2005 in Singapore and subsequently appeared in China, France and Israel.Citation16, Citation17 As a highly virulent pathogen, Ad55 was associated with life-threatening pneumonia during recent outbreaks.Citation18, Citation19 Their unpredictable high morbidity and mortality in otherwise healthy immuno-competent adults render Ad14 and Ad55 potential threats to public health.Citation20 Although the genomic and virological characteristics of Ad14 and Ad55 have been investigated,Citation21, Citation22 the prevalence of neutralizing antibodies (nAbs) in the general populations, which is helpful for understanding human anti-Ad14 and Ad55 immunity, remains unclear.
In this study, we established our micro-neutralizing (MN) assays on the basis of recombinant Ad14 and Ad55 reporter viruses, and we investigated the seroprevalence of nAbs against these two AdVs in healthy blood donors in Southern China. The distributions of nAbs in relation to different age, gender and blood type groups were also analyzed.
MATERIALS AND METHODS
Generation of recombinant Ad14 and Ad55 reporter viruses
Wild-type Ad14 (GenBank NO. JQ824845.1) and Ad55 (GenBank NO. KF911353.1) were kindly provided by Prof Qiwei Zhang (Southern Medical University, China) and Prof Chengfeng Qin (Beijing Institute of Microbiology and Epidemiology, China), respectively. In brief, wild-type Ad14 and Ad55 were propagated in A549 cells (ATCC, Manassas, VA, USA). The viral genomes were extracted and purified by sodium-dodecyl-sulfonate lysis (Sigma-Aldrich, St Louis, MO, USA) followed by phenol-chloroform extraction. The terminal regions of the Ad14 and Ad55 genomes were amplified by PCR and subcloned into pMD18T vectors (TaKaRa, Dalian, China) to obtain shuttle plasmids. The genomic plasmids pAd14 and pAd55 were then generated by homologous recombination between linearized shuttle plasmids and viral genomes in Escherichia coli BJ5183 competent cells (Agilent Technologies, Santa Clara, CA, USA). The fragments flanking the E3 region were subsequently amplified by PCR and subcloned into a pVAX1 plasmid (Thermo Fisher Scientific, Waltham, MA, USA) to obtain p14E3LR and p55E3LR. The E3 regions were then deleted by homologous recombination between E3LR fragments and linearized genomic plasmids to obtain pAd14ΔE3 and pAd55ΔE3. Simultaneously, the coding sequences for secreted alkaline phosphatase (SEAP) or enhanced green fluorescent protein (EGFP), which was flanked by a CMV promoter and a BGH poly(A) signal, were amplified by PCR and inserted into p14E3LR and p55E3LR to obtain shuttle reporter plasmids. Finally, pAd14-EGFP, pAd14-SEAP, pAd55-EGFP, and pAd55-SEAP were constructed by homologous recombination between linearized pAd14ΔE3 or pAd55ΔE3 and shuttle reporter plasmids. The genomes of the Ad14 and Ad55 reporter viruses were released by restricted enzyme digestion with AsiSI (New England Biolabs, Ipswich, MA, USA) and transfected into HEK293 cells (ATCC). The four reporter viruses Ad14-EGFP, Ad14-SEAP, Ad55-EGFP and Ad55-SEAP were then rescued and propagated. Purified viral stocks were obtained by cesium chloride gradient centrifugation, and the infectious virions were titrated as described.Citation23, Citation24
Human sample collection
Serum samples from 1009 healthy blood donors were randomly collected at the Guangzhou Blood Centre, Guangzhou, China, from August 2015 to December 2015. Any factors that could lead to a biased analysis, such as the educational background and the careers of the donors, were not considered as the criteria during the collection of blood samples. The ages of the volunteers ranged from 18 to 57 years old. The percentages of males to females were 31.7% and 68.3%. The percentages of blood types A, B, AB and O were 24.4%, 41.2%, 11.5% and 22.9%, respectively. The research using human samples was approved by the Ethics Committee of Guangzhou Institutes of Biomedicine and Health (GIBH). Informed consent was obtained from each volunteer.
Micro-neutralizing (MN) assays
The titers of nAbs that acted against Ad14 and Ad55 were detected using MN assays as previously described.Citation23, Citation24 In brief, the HEK293 cells were seeded into 96-well plates at 3 × 104 cells per well. One day later, serial dilutions of the human serums were heat-treated at 56 °C for 90 min and incubated with Ad14-SEAP or Ad55-SEAP at 4 × 106 viral particles per well for 1 h at 37 °C. The mixtures were subsequently added to the 96-well plates and incubated for 24 h at 37 °C. Finally, the supernatants were harvested, and the SEAP activity was detected using the Phospha-Light System according to the manufacturer’s instructions (Thermo Fisher Scientific). The relative light units (RLUs) were recorded and the titers were calculated as the dilutions that inhibited 50% RLU values. The values <72, 72–200, 201–1000 and >1000 were denoted as negative, low, moderate and high nAb titers, respectively. To validate the feasibility of SEAP activity-based MN assays, HEK293 cells were seeded into 96-well plates at 3 × 104 cells per well. Twenty-four hours later, serial dilutions of serum samples with low, moderate or high titers of Ad14 or Ad55 nAbs were incubated with Ad14-EGFP or Ad55-EGFP (4 × 106 viral particles per well), respectively. Subsequently, the mixtures were added to the plates. At 24 h after infection, EGFP-expressing cells were imaged using fluorescence microscopy.
Statistical analysis and figure development
Seroprevalence comparisons for different groups were conducted using a χ2-test. The trends in the seropositive rates throughout the groups were analyzed by using the Trend χ2-test. Comparisons of nAb titers among groups were performed by Mann–Whitney test. The trends in the nAb titers throughout the groups were analyzed by Kruskal–Wallis Test. All the statistical analyses were computed with SPSS version 13.0 (SPSS Inc., Chicago, IL, USA), and P values of less than 0.05 were considered statistically significant. The graphs were generated with GraphPad Prism version 5 (GraphPad Software, La Jolla, CA, USA) and Photoshop version 8 (Adobe Systems Incorporated, San Jose, CA, USA).
RESULTS
Establishment and validation of MN assays based on recombinant Ad14 and Ad55 reporter viruses
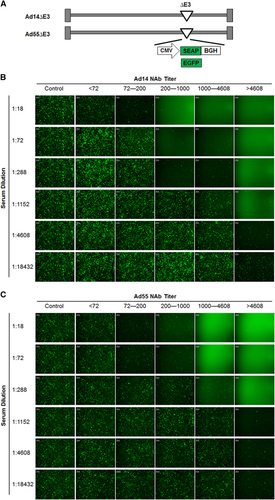
To generate replication-competent reporter viruses, the E3 regions of Ad14 and Ad55 were depleted, and the expression cassettes for EGFP- or SEAP were inserted (). The four reporter viruses Ad14-EGFP, Ad14-SEAP, Ad55-EGFP and Ad55-SEAP, were successfully obtained. The reporter genes were efficiently expressed in cells that were infected by these viruses (data not shown).
To validate the feasibility of the MN assays based on Ad14-SEAP and Ad55-SEAP, serums with different nAb titers (<72, 72–200, 201–1000, 1001–4608 and >4608) were serially diluted, incubated with Ad14-EGFP or Ad55-EGFP and infected into HEK293 cells. The results showed that serums with nAb titers <72 displayed no detectable inhibition in relation to Ad14-EGFP or Ad55-EGFP infection for all the dilutions, while those with nAb titers >72 exhibited various neutralizing activities, which was consistent with the titers that were determined by SEAP activity (). In addition, the MN assays based on SEAP or EGFP reporter genes were much more sensitive than the ones based on cytopathic effects (CPEs) (Supplementary Figure S1). These results suggested that the MN assays based on Ad14-SEAP and Ad55-SEAP were feasible.
The overall seroprevalence of Ad14 and Ad55 nAbs in healthy blood donors from Southern China
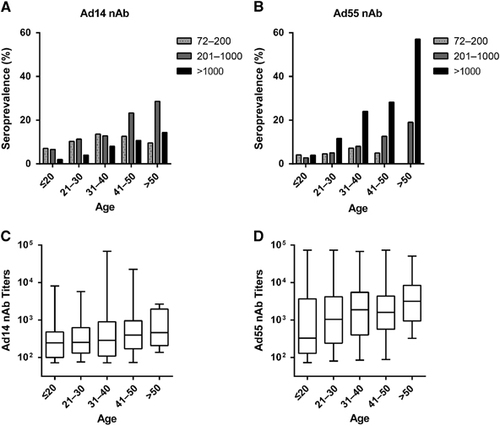
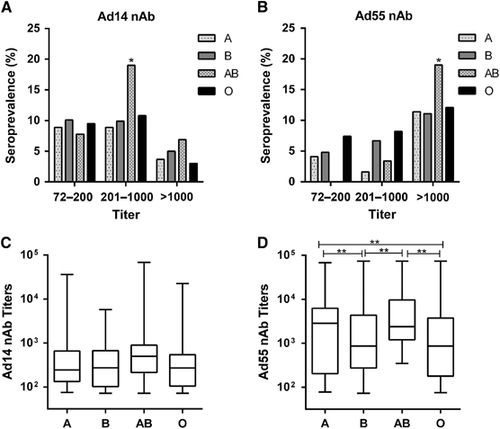
By using Ad14-SEAP and Ad55-SEAP, the nAbs in the serums from 1009 healthy blood donors were detected (Supplementary Tables S1 and S2). The overall seroprevalence levels of Ad14 and Ad55 nAbs were 24.8% (95% confidence interval, CI: 22.1%–27.5%) and 22.4% (95% CI: 19.8%–25.0%), respectively (Supplementary Tables S1 and S2). The trends in the Ad14 and Ad55-seropositive rates were similar in this cohort, and both increased with age (Trend Chi-square test, Ad14: P-value <0.001; Ad55: P-value <0.001; ). Notably, the seropositive rates in individuals below 20 years of age were extremely low for both Ad14 and Ad55 (15.5% and 10.9%, respectively; ), revealing that the infections by these two AdVs and the circulation of nAbs in young adults were relatively rare. The donors were categorized into four groups according to their nAb titers as follows: negative, <72; low, 72–200; moderate, 200–1000; and high, >1000. The distributions of the different nAb titers in the whole cohort were analyzed. Moderate and low nAb titers were frequently detected in Ad14-seropositive donors, while the majority of Ad55-seropositive ones had high nAb titers (), suggesting that Ad55 infection might elicit stronger nAb responses in comparison with Ad14 infection.
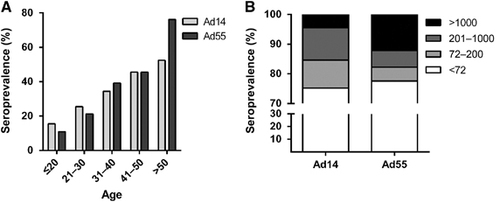
Titer distributions of Ad14 and Ad55 nAbs in different age groups
We then analyzed the distributions of nAb titers by age group. The frequency of serum samples with moderate or high nAb titers increased with age (Trend χ2-test, P-value <0.001 for moderate and high Ad14 and Ad55 nAbs, respectively), while the frequency for low nAb titers increased before 40 years old and decreased thereafter for both Ad14 and Ad55 (). Individuals with moderate nAb levels were dominant in Ad14-seropositive individuals older than 40 (), whereas the majority of Ad55-seropositive individuals older than 20 had high nAb titers (). Interestingly, almost 60% of volunteers older than 50 generated high levels of Ad55 nAb, and none of the individuals in this subgroup had a low titer of Ad55 nAb (). Thus, the overall seroprevalence levels for Ad14 and Ad55 nAbs were comparable, but their titer distributions within different age groups were dramatically different. Although no significant difference was observed for the Ad14 nAb levels in different age groups (Kruskal–Wallis test, P-value=0.07; ), the Ad55 nAb levels definitely increased with age (Kruskal–Wallis test, P-value <0.05; ), suggesting that nAb responses to Ad55 but not Ad14 infection tended to be enhanced when these individuals were re-infected by these strains.
Seropositive rate and titer distributions of Ad14 and Ad55 nAbs in different gender and blood type groups
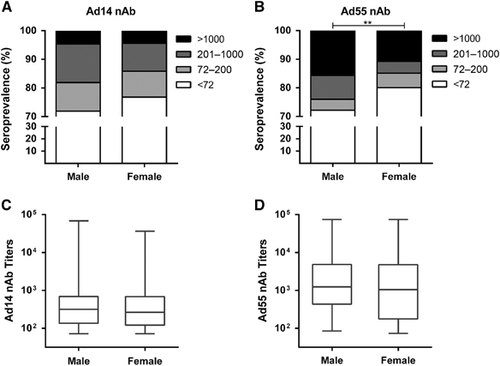
There was no significant difference for the Ad14-seropositive rates in the gender groups (χ2-test, P-value=0.093; ), whereas a higher Ad55-seropositive rate was detected in males compared to females (χ2-test, P-value <0.01; ). The titers of either Ad14 or Ad55 nAb were comparable in males and females (Mann–Whitney Test, P-value=0.552 and 0.600 for Ad14 and Ad55 nAb, respectively; ), suggesting that the gender was not a major impacting factor on either the Ad14 or Ad55 nAb levels. Notably, the frequency of Ad14-seropositive samples, especially those with moderate nAb titers, was significantly higher in the AB blood type group than in the other blood type groups (χ2-test, P-value <0.05; ). In addition, the percentage of Ad55-seropositive samples with high nAb titers was also higher in this group (χ2-test, P-value <0.05; ). In addition, none of the volunteers with blood type AB had low Ad55 nAbs, and significantly fewer blood type A or AB donors generated moderate Ad55 nAbs (), which might imply that once the individuals with blood type AB were infected by Ad55, high titers of nAbs would be elicited. Consistent with this point, although the AB blood type individuals only had slightly higher levels of Ad14 nAb compared to other blood type groups (Kruskal–Wallis test, P-value=0.086; ), the titer of Ad55 nAb in this group was significantly higher than that of the blood type B or O groups (Mann–Whitney test, P-value <0.05; ). These results suggested that the blood type may influence Ad14 and Ad55 infection and the generation of nAb responses to these two AdVs.
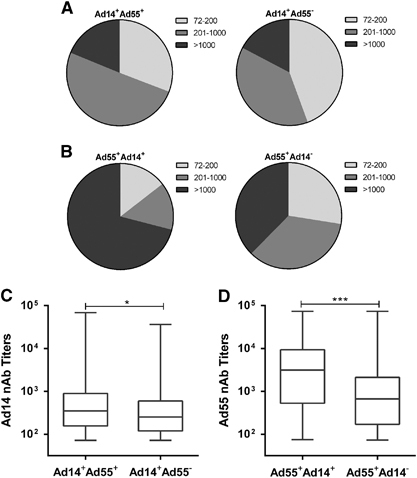
Seroprevalence of Ad14 nAb in Ad55-seropositive donors was higher than that in Ad55-seronegative ones, and vice versa
Next, we analyzed the frequency of Ad14-seropositive donors in Ad55-seropositive or seronegative individuals. The Ad14-seropositive rate was significantly higher in Ad55-seropositive donors than in Ad55-seronegative ones (Mann–Whitney test, P-value <0.01; Table ). Similar trends were detected for the Ad55-seropositive rate in Ad14-seropositive or seronegative donors (P-value <0.01; Table ). The frequency of blood donors with moderate Ad14 nAb levels was slightly higher in Ad55-seropositive compared to Ad55-seronegative donors (). However, in Ad14-seropositive donors, many more individuals had high titers of Ad55 nAb in comparison with Ad14-seronegative ones (χ2-test, P-value <0.001; ). Interestingly, the titers of Ad14 nAb in Ad55-seropositive donors were significantly higher than they were in Ad55-seronegative ones (Mann–Whitney test, P-value <0.05; ). In addition, the Ad55 nAb titers in Ad14-seropositive donors were much higher than they were in Ad14-seronegative donors (P-value <0.001; ). All these results suggested that Ad55-seropositive donors tended to be Ad14-seropositive and vice versa, and double-positive serums had higher nAb titers than single-positive ones.
Table 1 Correlation of Ad14 and Ad55 nAb seropositive rates
DISCUSSION
Recently, two highly virulent AdVs called Ad14 and Ad55 have attracted increasing attention; these viruses caused severe clinical consequences in immuno-competent adults over a series of outbreaks.Citation2 Clarifying the circulation of Ad14 and Ad55 nAbs in general populations is essential for understanding the herd immunity. Here, we investigated the seroprevalence of Ad14 and Ad55 nAbs in a large cohort from Southern China, and we found that the seropositive rates of Ad14 and Ad55 were much lower than that of Ad5 according to our previous results.Citation24 This finding may imply that the general populations, especially those younger than 40, were threatened by these two AdVs. Therefore, Ad14 and Ad55 should be taken into account when patients with severe ARDs are hospitalized.Citation2
In this study, we adopted EGFP- or SEAP-expressing reporter viruses based on clinical isolates of Ad14 and Ad55, and we established sensitive and high-throughput MN assays for assessing the nAbs against these two AdVs (). These assays possessed at least two advantages over the methods that were described previously.Citation25 Because the nAbs against AdV could be raised against hexon as well as fiber proteins, the chimeric Ad3 that carried the Ad14 hexon as well as Ad3 fiber proteins could only detect Ad14 nAbs that were specific for hexon.Citation26 In addition, MN assays based on SEAP-expressing AdVs were reported to be more sensitive and objective than previously used CPE-based methods.Citation27 Consistent results were obtained when SEAP and EGFP reporter viruses were applied (), reflecting the feasibility of these MN assays. Given that nAbs have shown beneficial effects for treating patients infected with H5N1 influenza virus or Ebola virus,Citation28, Citation29 these MN assays could be utilized to screen serums containing high levels of nAbs or monoclonal nAbs against Ad14 and Ad55, to treat patients with severe illnesses caused by these two AdVs (unpublished data).
For the purpose of genetic vector development, numerous studies have investigated the pre-existing nAbs against human and chimpanzee AdVs.Citation24, Citation26, Citation27, Citation30, Citation31, Citation32, Citation33, Citation34 Regarding Ad14 and Ad55, although several case reports or epidemiological studies have been described,Citation5, Citation6, Citation7, Citation8, Citation9, Citation11, Citation12, Citation13, Citation14, Citation35 the surveillance of their pre-existing nAbs was seldom performed. One recent study described seropositive rates of Ad55 nAbs in pediatric patients, and another study analyzed the Ad14 nAbs in a small population.Citation25, Citation36 However, the seroprevalence of Ad14 and Ad55 nAbs in healthy adults remained to be explored, and it relied on randomly collected large-scale cohorts. Moreover, detailed analyses within age, gender or blood type groups could not be performed due to the limited samples in these two studies. Here, we investigated the seroprevalance of Ad14 and Ad55 nAbs based on a randomly collected cohort including up to 1009 healthy blood donors, and we found that the seropositive rates in young adults were low and increased with age (), which was in accordance with the findings observed in other AdVs.Citation32 The seropositive rates and nAb titers in different subgroups were successfully analyzed. We also found that Ad14-seropositive donors tended to generate moderate nAb titers while Ad55-seropositive ones were prone to having high nAb titers, especially in individuals older than 20 (). These findings added to our knowledge of human immunity against Ad14 and Ad55 infections.
Similar to the results of other studies,Citation33, Citation37 the seroprevalence of Ad14 and Ad55 nAbs was independent of the donor’s gender (). However, our results showed that the blood type significantly impacted the susceptibility and nAb responses of individuals who were exposed to Ad14 or Ad55 infection (). Although a relationship between the blood type and AdV infection was not described previously, the histo-blood group antigens were involved in the infection of hepatitis B virus and norovirus.Citation38, Citation39, Citation40 We showed that individuals with blood type AB displayed the highest seropositive rates for Ad14 and Ad55 nAbs (), indicating that this group of people might be more susceptible to infection by Ad14 or Ad55. In addition, the nAb titers in AB blood type donors were higher than that of other groups, revealing that stronger immune responses were induced by Ad14 or Ad55 infection in these individuals.
We also showed that Ad14-seropositive donors tended to have higher Ad55-seropositive rates and nAb titers than Ad14-seronegative ones, and vice versa (), indicating that there was a positive correlation between Ad14 and Ad55 seroprevalence. The nAb titers in double-positive donors were significantly higher than they were in Ad14 or Ad55 single-positive ones (). These results may be attributed to fiber protein, which is one of the major capsid antigens of AdVs. As a recombinant of Ad11 and Ad14, Ad55 shares a similar hexon protein with Ad11 and a fiber protein with Ad14.Citation21 Although many studies reported that hexon-specific antibodies were predominantly elicited in individuals who were infected by other AdVs,Citation41, Citation42 fiber-specific nAbs were also frequently detected.Citation26, Citation43, Citation44 If fiber-specific nAbs were generated in individuals who were infected by either Ad14 or Ad55, it may neutralize both variants and thus contribute to the cross-neutralizing activity of double-positive serums. However, these speculations should be confirmed in future studies.
In summary, we established convenient MN assays to survey the effects of nAbs against Ad14 and Ad55, and we investigated the seroprevalence of these two AdVs in a general population from Southern China. We also provided the distributions of nAbs by age, gender and blood type groups. These results emphasized the importance of monitoring these two AdVs in general populations and developing vaccines for disease prevention.
Supplementary Figure S1
Download PDF (49 KB)Supplementary Table S1
Download MS Word (20.1 KB)Supplementary Table S2
Download MS Word (20.1 KB)Acknowledgments
This study was supported by the National Natural Science Foundation of China (NO 31200693 and 31470892), the Guangzhou Health Care and Cooperative Innovation Major Project (NO 201508020252), the Pearl River S&T Nova Program of Guangzhou (NO 201506010076), and a grant from the CAS Youth Innovation Promotion Association to Liqiang Feng (NO 2014328). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. We also thanked Prof Qiwei Zhang (Southern Medical University, China) and Prof Chengfeng Qin (Beijing Institute of Microbiology and Epidemiology, China) for their kind gift of the Ad14 and Ad55 viruses.
Supplementary Information for this article can be found on the Emerging Microbes & Infections website (http://www.nature.com/emi)
Related Research Data
References
- Davison AJ, Benko M, Harrach B.Genetic content and evolution of adenoviruses. J Gen Virol 2003;84: 2895–2908.
- Tan D, Zhu H, Fu Yet al.Severe community-acquired pneumonia caused by human adenovirus in immunocompetent adults: a multicenter case series. PLoS One 2016;11: e0151199.
- Kunz AN, Ottolini M.The role of adenovirus in respiratory tract infections. Curr Infect Dis Rep 2010;12: 81–87.
- Tebruegge M, Curtis N.Adenovirus infection in the immunocompromised host. Adv Exp Med Biol 2010;659: 153–174.
- Chmielewicz B, Benzler J, Pauli Get al.Respiratory disease caused by a species B2 adenovirus in a military camp in Turkey. J Med Virol 2005;77: 232–237.
- Lewis PF, Schmidt MA, Lu Xet al.A community-based outbreak of severe respiratory illness caused by human adenovirus serotype 14. J Infect Dis 2009;199: 1427–1434.
- Jin Y, Zhang RF, Xie ZPet al.Prevalence of adenovirus in children with acute respiratory tract infection in Lanzhou, China. Virol J 2013;10: 271.
- Lu QB, Tong YG, Wo Yet al.Epidemiology of human adenovirus and molecular characterization of human adenovirus 55 in China, 2009-2012. Influenza Other Respir Viruses 2014;8: 302–308.
- Chen M, Zhu Z, Huang Fet al.Adenoviruses associated with acute respiratory diseases reported in Beijing from 2011 to 2013. PLoS One 2015;10: e0121375.
- Vanderveen J, Kok G.Isolation and typing of adenoviruses recovered from military recruits with acute respiratory disease in the Netherlands. Am J Hyg 1957;65: 119–129.
- Metzgar D, Osuna M, Kajon AEet al.Abrupt emergence of diverse species B adenoviruses at US military recruit training centers. J Infect Dis 2007;196: 1465–1473.
- Huang G, Yu D, Zhu Zet al.Outbreak of febrile respiratory illness associated with human adenovirus type 14p1 in Gansu Province, China. Influenza Other Respir Viruses 2013;7: 1048–1054.
- Kajon AE, Lu X, Erdman DDet al.Molecular epidemiology and brief history of emerging adenovirus 14-associated respiratory disease in the United States. J Infect Dis 2010;202: 93–103.
- Carr MJ, Kajon AE, Lu Xet al.Deaths associated with human adenovirus-14p1 infections, Europe, 2009-2010. Emerg Infect Dis 2011;17: 1402–1408.
- Tang L, An J, Xie Zet al.Genome and bioinformatic analysis of a HAdV-B14p1 virus isolated from a baby with pneumonia in Beijing, China. PLoS One 2013;8: e60345.
- Salama M, Amitai Z, Nutman Aet al.Outbreak of adenovirus type 55 infection in Israel. J Clin Virol 2016;78: 31–35.
- Lafolie J, Mirand A, Salmona Met al.Severe pneumonia associated with adenovirus type 55 infection, France, 2014. Emerg Infect Dis 2016;22: 2012–2014.
- Walsh MP, Seto J, Jones MSet al.Computational analysis identifies human adenovirus type 55 as a re-emergent acute respiratory disease pathogen. J Clin Microbiol 2010;48: 991–993.
- Cao B, Huang G-H, Pu Z-Het al.Emergence of community-acquired adenovirus type 55 as a cause of community-onset pneumonia. Chest 2014;145: 79–86.
- Schildgen O, Gu L, Qu Jet al.Sustained viremia and high viral load in respiratory tract secretions are predictors for death in immunocompetent adults with adenovirus pneumonia. Plos One 2016;11: e0160777.
- Liu J, Nian QG, Zhang Yet al.In vitro characterization of human adenovirus type 55 in comparison with its parental adenoviruses, types 11 and 14. PLoS One 2014;9: e100665.
- Radke JR, Yong SL, Cook JL.Low-level expression of the E1B 20-kilodalton protein by adenovirus 14p1 enhances viral immunopathogenesis. J Virol 2016;90: 497–505.
- Aste-Amezaga M, Bett AJ, Wang Fet al.Quantitative adenovirus neutralization assays based on the secreted alkaline phosphatase reporter gene: application in epidemiologic studies and in the design of adenovector vaccines. Hum Gene Ther 2004;15: 293–304.
- Sun C, Zhang Y, Feng Let al.Epidemiology of adenovirus type 5 neutralizing antibodies in healthy people and AIDS patients in Guangzhou, southern China. Vaccine 2011;29: 3837–3841.
- Su XB, Tian XG, Jiang ZXet al.Human adenovirus serotype 3 vector packaged by a rare serotype 14 hexon. Plos One 2016;11: e0156984.
- Yu B, Dong J, Wang Cet al.Characteristics of neutralizing antibodies to adenovirus capsid proteins in human and animal sera. Virology 2013;437: 118–123.
- Zhang S, Huang W, Zhou Xet al.Seroprevalence of neutralizing antibodies to human adenoviruses type-5 and type-26 and chimpanzee adenovirus type-68 in healthy Chinese adults. J Med Virol 2013;85: 1077–1084.
- Mupapa K, Massamba M, Kibadi Ket al.Treatment of Ebola hemorrhagic fever with blood transfusions from convalescent patients. J Infect Dis 1999;179: S18–S23.
- Zhou BP, Zhong NS, Guan Y.Treatment with convalescent plasma for influenza A (H5N1) infection. N Engl J Med 2007;357: 1450–1451.
- Mast TC, Kierstead L, Gupta SBet al.International epidemiology of human pre-existing adenovirus (Ad) type-5, type-6, type-26 and type-36 neutralizing antibodies: correlates of high Ad5 titers and implications for potential HIV vaccine trials. Vaccine 2010;28: 950–957.
- Abbink P, Lemckert AA, Ewald BAet al.Comparative seroprevalence and immunogenicity of six rare serotype recombinant adenovirus vaccine vectors from subgroups B and D. J Virol 2007;81: 4654–4663.
- Thorner AR, Vogels R, Kaspers Jet al.Age dependence of adenovirus-specific neutralizing antibody titers in individuals from sub-Saharan Africa. J Clin Microbiol 2006;44: 3781–3783.
- Wang X, Xing M, Zhang Cet al.Neutralizing antibody responses to enterovirus and adenovirus in healthy adults in China. Emerg Microbes Infect 2014;3: e30.
- Huang D, Hennequi M, Elvachev Aet al.The sero-prevalence of anti-adenovirus 5 neutralizing antibodies is independent of a chronic hepatitis B carrier state in China. Arch Virol 2015;160: 1125–1130.
- Kajon AE, Dickson LM, Metzgar Det al.Outbreak of febrile respiratory illness associated with adenovirus 11a infection in a Singapore military training cAMP. J Clin Microbiol 2010;48: 1438–1441.
- Tian X, Jiang Z, Ma Qet al.Prevalence of neutralizing antibodies to common respiratory viruses in intravenous immunoglobulin and in healthy donors in southern China. J Thorac Dis 2016;8: 803–812.
- Yang W-X, Zou X-H, Jiang S-Yet al.Prevalence of serum neutralizing antibodies to adenovirus type 5 (Ad5) and 41 (Ad41) in children is associated with age and sanitary conditions. Vaccine 2016;34: 5579–5586.
- Shirato H.Norovirus and histo-blood group antigens. Jpn J Infect Dis 2011;64: 95–103.
- Branch DR.Blood groups and susceptibility to virus infection: new developments. Curr Opin Hematol 2010;17: 558–564.
- Wang D-S, Chen D-L, Ren Cet al.ABO blood group, hepatitis B viral infection and risk of pancreatic cancer. Int J Cancer 2012;131: 461–468.
- Roberts DM, Nanda A, Havenga MJet al.Hexon-chimaeric adenovirus serotype 5 vectors circumvent pre-existing anti-vector immunity. Nature 2006;441: 239–243.
- Qiu H, Li X, Tian Xet al.Serotype-specific neutralizing antibody epitopes of human adenovirus type 3 (HAdV-3) and HAdV-7 reside in multiple hexon hypervariable regions. J Virol 2012;86: 7964–7975.
- Bradley RR, Lynch DM, Iampietro MJet al.Adenovirus serotype 5 neutralizing antibodies target both hexon and fiber following vaccination and natural infection. J Virol 2012;86: 625–629.
- Cheng C, Gall JG, Nason Met al.Differential specificity and immunogenicity of adenovirus type 5 neutralizing antibodies elicited by natural infection or immunization. J Virol 2010;84: 630–638.