Abstract
CapsuleBeneficial effects of cutting grass are relatively short-lived for a range of bird species.
AimsTo investigate how cutting affects a range of birds occurring on farmland, how long these effects last, and whether there is any effect of the timing of cutting operations.
MethodsWe surveyed birds on 33 grass fields on three farms in northern England, both before and after agricultural cutting operations. The data were then modelled using generalized linear mixed models (GLMMs).
Results Species relying on below-ground invertebrates (e.g. corvid species) prefer cut swards, while some species make greater use of longer grass swards (e.g. pheasant). Use of fields after cutting declined by 50 % by day 11 for corvids, by day 8 for aerial feeders (e.g. hirundines), by day 6 for gulls, and by day 4 for kestrels.
ConclusionsWe recorded mainly common, generalist species that make use of agricultural grassland. The foraging behaviour of these species is similar to other, rarer or declining species, and so our findings can be extrapolated to a range of farmland birds. However, it is important to note that some studies indicate direct negative effects of cutting on some species not found in our surveys.
Farmland biodiversity has been in decline since the mid 20th century, and this has been linked to the increasing intensity of agriculture during that period (Benton et al. Citation2003). Farmland birds have been the subject of much agro-ecological research, and there is clear evidence linking changes in agricultural practice to declines in farmland bird populations across Europe (e.g. Donald et al. Citation2001, Citation2006, Gregory et al. Citation2005). Agri-environment Schemes (AESs) have been introduced in many countries throughout Europe, in part, at least, to lessen these negative effects on biodiversity (Whittingham Citation2007). Within the category ‘farmland birds’ are many species which rely mainly on grassland and the management options for grassland birds within AESs have been informed by several recent studies (e.g. Perkins et al. Citation2000, Vickery et al. Citation2001, Barnett et al. Citation2004, Atkinson et al. Citation2004, Citation2005, Whittingham & Devereux Citation2008, Buckingham et al. Citation2004, Citation2006). Grassland sward management has consistently come out as a key management tool to manipulate predation risk, and also invertebrate food abundance and accessibility, by enabling birds more readily to access below-ground grassland invertebrates (Atkinson et al. Citation2004, Buckingham et al. Citation2006). Changes to predation risk and food abundance/accessibility all combine to alter intake rates, which ultimately alter the quality for patch choice (e.g. Stephens & Krebs Citation1986, Lima & Dill Citation1990, Whittingham et al. Citation2004).
Some popular AES options focus on the creation and maintenance of new grassland habitats, such as grass margins, buffer strips and low input grassland (Natural England Citation2010a,Citationb), but once in place, these newly created grassland areas need careful management in order to maximize their benefits for wildlife. We know that changing sward height alters its attractiveness to a range of bird species (e.g. Perkins et al. Citation2000, Atkinson et al. Citation2004, Citation2005, Whittingham & Devereux Citation2008, Douglas et al. Citation2009), but it is not clear over what time period short grass acts as high-quality habitat for the birds which use it. Such information is useful if we are to adapt current AES sward management prescriptions to maximize the usefulness of those swards for as wide a number of species as possible.
Conversely, we sometimes need to understand bird habitat usage in order to discourage their use of an area, such as the large patches of grasslands at airports (e.g. Brough & Bridgman Citation1980, Blackwell et al. Citation2009). Birds are a primary risk group for causing aircraft crashes (e.g. Blackwell et al. Citationin review), and so discouraging the use of grasslands by some larger species, such as corvids, gulls and wildfowl, and even smaller species, such as Common Starlings Sturnus vulgaris, is important to minimize strike risk.
In this study we show the relationship between temporal aspects of cutting and use of grassland by a range of birds (see Methods and Materials for list of study groups), and demonstrate that the drop-off in use of cut fields is rapid. As a secondary aim we also repeat earlier work investigating the effects of cutting per se on grassland use by farmland birds in the summer and our results concur with earlier work. It is important to note that past studies have shown that some species (not found in our study) are negatively affected by grass cutting and so our study needs to be interpreted in light of these studies (see conclusions).
METHODS AND MATERIALS
CTP surveyed 33 grass fields at three farms (4, 20 and 9 fields at each farm, respectively) in County Durham, England, over 70 days from mid June to August, 2009. The mean field size was 8.47 ± 1.5 (1 se) ha (range: minimum 1 ha, maximum 26 ha). Twenty-nine of the fields were permanent pasture (they had been grass fields for more than 5 years), and four fields were grass leys (they had been ploughed less than 5 years ago).
Surveys were not conducted during wet and/or windy weather, due to the effects of those conditions on visibility, hindering bird identification and observed activity (Bibby et al. Citation1992). All surveys were carried out between 10.00 and 18.30 (BST) to avoid periods when birds were either leaving or arriving at roost sites. All fields were cut for either silage or hay during the observation period (at farm 1: three fields cut between 25 July and 1 August with one field cut on 24 June; at farm 2: all fields cut between 27 July and 7 August; at farm 3: all fields cut between 28 July and 17 August), so the use of fields by foraging birds was recorded on each field both before and after grass cutting.
Vegetation height measurements
Pre-cut sward height was measured during the first survey of each field and post-cut sward height was measured during the first survey following cutting. Ten random sites were chosen on each field and height was obtained using a tape measure, and then the mean for each field was used to calculate the data presented in the results.
Pre-cut bird surveys
An average of 2.2 visits (maximum visits = 4, minimum = 2) were made to each of the 33 fields before they were cut. All visits were at least one day apart except for two fields which were visited twice on the same day (early and late in the day).
Field surveys consisted of walking transects from one side of the field to the opposite side (the starting point for the first transect in each field was chosen randomly). Transects were used to avoid bias in the observability of birds between fields with differing grass length, as birds hiding in the long grass were flushed by the observer. The transects were approximately 50 m apart so that the observer visited every part of the field to within 25 m. Birds that were either flushed from the grass by the observer, foraging in the grass, or foraging over the grass (e.g. Common Kestrels Falco tinnunculus hovering and aerial feeders flying low (<5 m) over the field to forage) were identified to species level and their numbers recorded (the size of large flocks was estimated as accurately as possible). Birds flying over fields but not landing on them (other than in the circumstances outlined above), or those at the edge of the field (e.g. in hedges) were not recorded. Although the three study farms were separated geographically, within each farm some of the surveyed fields were adjacent to one another. In order to minimize double-counting of birds, the observer noted the direction taken by birds which were flushed during the course of the surveying of each field, and thus avoided (as far as possible) re-counting the same individual in adjacent fields surveyed on the same visit.
Post-cut bird surveys
An average of 4.8 visits (maximum visits = 9, minimum = 3) were made to each of the 33 grass fields after they were cut for harvest. Those surveys carried out while grass was being cut (0 days post-cut), turned, or baled did not involve transects, but instead vantage point observations were made from a point nearby. Otherwise, transects every 50 m were carried out and birds recorded as detailed above. If livestock were introduced to a field in the intervening period between observation visits, then no more surveys of that field were carried out to avoid conflicting effects due to grazing.
Statistical analysis
We grouped species into functional units for analysis. We did this for two reasons. First, numbers of some species were too few to analyse independently. Secondly, and more important, some bird species feed in similar ways; for example, hirundines and Common Swifts Apus apus all feed on aerial insects and so it was sensible to group them together. Likewise, corvids all feed mainly on ground-dwelling and below-ground invertebrates. Thirdly, because some species often occur in mixed species flocks, each species is not likely to be statistically independent of other similar species (e.g. the presence of foraging Eurasian Jackdaws Corvus monedula L. is likely to attract Rooks Corvus frugilegus L. and Carrion Crows Corvus corone L.). Thus it makes ecological sense to group the species recorded into six functional units for analysis: (1) aerial feeders (Barn Swallows Hirundo rustica, House Martins Delichon urbicum, Common Swifts); (2) corvids (Carrion Crows, Eurasian Jackdaws, Rooks and Black-billed Magpies Pica pica); (3) gulls (Black-headed Gulls Larus ridibundus, Common Gulls Larus canus); (4) pigeons (Common Wood Pigeons Columba palumbus, Eurasian Collared Doves Streptopelia decaocto; (5) Common Kestrels; (6) Common Pheasants Phasianus colchicus.
There is a potential lack of independence because some of the fields at each farm were located in blocks bordering one another, and so the numbers of birds on any given field could affect numbers on nearby fields. However, this issue was minimized by making repeat visits to the same fields (see above), such that use of a given field was more likely to be picked up over multiple visits. In addition, this source of lack of independence is likely to cause Type 2 error (absence of a significant effect of birds selecting shorter grass fields when there is actually one present) and so the results we report are likely to be robust.
Generalized Linear Mixed Models (GLMMs) were used to analyse the data in R (version 2.9.1) (R Development Core Team Citation2009) using the package lme4 v.0.99875.9 (Bates et al. Citation2008). The response variable was the number of occasions that each functional group was recorded in a field, and the number of surveys was specified as the binomial denominator [with a binomial error structure with logit link function (Crawley Citation2007), e.g. if corvids were present on two visits out of six then two was specified as the response variable and six as the denominator]. The assumption within our analysis is that birds can make an independent choice of fields on each visit. The method we use has been previously shown to correlate number of occurrences (e.g. species ‘a’ was recorded on field ‘b’ on two out of six visits) with total count of birds on a field (e.g. 14 individuals of species ‘a’ recorded on field ‘b’ across all six visits; see Perkins et al. Citation2000, Moorcroft et al. Citation2002). This method gives a good indication of ‘use’ of a particular field. While we acknowledge that this is not perfect, it was not possible to analyse total species counts because of highly skewed data. Many birds are social foragers and the decision of where to feed is influenced by where their flock-mates are. This renders count data non-independent and its analysis would violate statistical test assumptions.
The term ‘field’ was included in all models as a random effect to allow within-field comparisons, while controlling for variation resulting from site-specific parameters. Two models were fitted to each of the six functional groups. Model A included ‘cutting’, fitted as a two-way factor (before cutting was coded as ‘1’, and after cutting was coded as ‘2’) to account for the effects of manipulation of grass height. Model B was fitted in each instance with just data on post-cutting. Thus the factor ‘cutting’ was not included in Model B, but an additional two variables were incorporated into Model B: ‘timing of visit’ was incorporated into the model to investigate whether the probability of occurrence of birds on plots changed with time since mowing; ‘date of cutting’ was included, with the earliest cut field given a value of ‘1’ and the number of days between that and the cutting of each subsequent field then added (thus a field cut 10 days after the earliest field was assigned ‘11’). This was to investigate whether there was any difference in use of fields depending on whether they were cut earlier or later in the season. In addition a two-way factor for the presence or absence of ‘bales’ in the field and a three-level factor for ‘farm’ was also included in Model B. The former was fitted as it was noted that birds often made use of bales as perches.
The significance of each predictor in the analyses of both model A and B was assessed using the change in deviance (ΔD), which is distributed asymptotically as χ2, on removal of each term from a model including all predictors (i.e. a full model, Whittingham et al. Citation2006). The fit of the model to the assumptions of a binomial distribution can be approximated by comparing the ratio of residual deviance/residual degrees of freedom; ratios close to one indicate a reasonable fit to the data, whereas ratios greater than 2.5 indicate a poor, overdispersed fit (Crawley Citation1993). The data did not need to be corrected for overdispersion in any of the models due to low residual deviance to residual degrees of freedom ratios (highest ratio was 1.23). All probabilities quoted are two-tailed. Means and standard errors are presented in the form mean ± 1 se.
RESULTS
The mean pre-cut sward height over all fields was 89.2 cm (maximum mean height = 129.5 cm, minimum mean height = 69.0 cm), and the mean post-cut sward height over all fields was 4.6 cm (maximum mean height = 5.8 cm, minimum mean height = 3.6 cm).
During the surveys, 21 bird species (3958 individuals) were recorded (). Some species did not have sufficient data to be analysed (e.g. Sky Lark Alauda arvensis) (). Species with similar foraging behaviour and body size were grouped into functional groups for analysis (see , and Methods and Materials for a description of the groups and which bird species they contain).
Table 1. The total number of individuals of each species observed pre-cut and post-cut, with mean number per survey in brackets. For example, 207 swallows in total were counted on the pre-cut surveys, with an average of 2.80 swallows recorded per survey visit per field. A total of 74 pre-cut surveys were carried out in contrast to 158 post-cut surveys; therefore, the mean number per visit per field is a better indicator of differences between treatments than total numbers.
Table 2. The number of times each species was present out of 74 pre-cut surveys and 158 post-cut surveys and the same number expressed as a proportion in brackets. For example, one or more aerial feeders were recorded on 49 occasions pre-cut out of 74 visits, giving a proportion of 49/74 (0.66 or 66 %).
Effect on birds of cutting grass
Aerial feeders were more frequently recorded on fields before cutting occurred than after (χ2 = 53.01, 1 df, P < 0.001), as were Pheasants (χ2 = 34.40, 1 df, P < 0.001). Gulls, corvids and pigeons showed the reverse pattern and were recorded significantly more frequently after cutting (gulls: χ2 = 47.80, 1 df, P < 0.001, corvids: χ2 = 72.60, 1 df, P < 0.001, pigeons: χ2 = 11.10, 1 df, P < 0.001). There were more Kestrels present during post-cut observations, but this was not significant (χ2 = 2.01, 1 df, P = 0.160).
How did the date on which the field was cut and time since cutting affect birds?
Corvids were recorded more frequently on fields that were cut earlier in the year (date of cutting) (χ2 = 4.72, 1 df, P = 0.03) and there was a non-significant trend in the same direction for gulls (χ2 = 3.26, 1 df, P = 0.07). However, the date on which the field was cut did not affect the probability of field use by any other group: aerial feeders (χ2 = 0.57, 1 df, P = 0.45), Kestrels (χ2 = 0.33, 1 df, P = 0.56) and pigeons (χ2 = 0.63, 1 df, P = 0.42) (there were insufficient data to model the effect on pheasants).
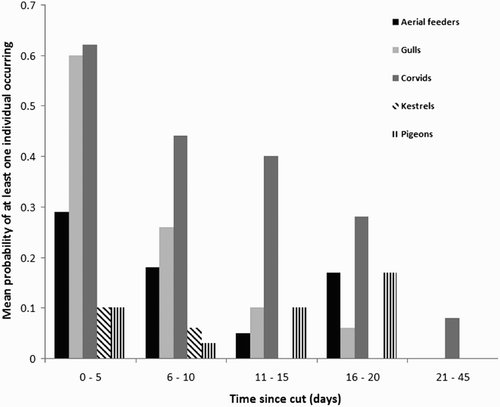
Time since cut had a significant negative effect on aerial feeders (χ2 = 11.21, 1 df, P < 0.001), gulls (χ2 = 51.81, 1 df, P < 0.001), corvids (χ2 = 24.5, 1 df, P < 0.001), and Kestrels (χ2 = 7.12, 1 df, P = 0.008) (). Time since cut did not have a significant effect on the presence of pigeons (χ2 = 0.04, 1 df, p = 0.85) and again there were insufficient data to analyse the data on Pheasants.
Figure 1. The effect of time since mowing on the presence of different birds during post-cut surveys. Mean values are the mean number of occurrences per survey over each time period. For example, aerial feeders occurred in 21 of the 73 post-cut surveys carried out between days 0 to 5, which gives a mean value of 0.29 aerial feeders per survey for that time period. Note that although statistical analysis was performed using the actual number of days since the field was mown, for ease of presentation we have shown the data here based on grouping into 5-day periods (except the last category which was summed into a 25-day period due to the scarcity of data for that period).
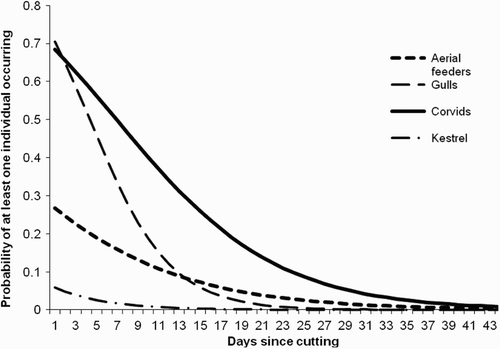
The best-fit relationship was then plotted for those relationships which were significant, to examine how quickly this drop off in use occurred (). Using this best-fit line, we calculated on which day since cutting began use dropped by more than 50 % from the starting level, and found this occurred for corvids by day 11, aerial feeders by day 8, gulls by day 6, and Kestrels by day 4.
Figure 2. Effect of time since cutting on the likelihood of recording four functional groups of birds. All relationships shown were significant (see Results) and the back-transformed parameter estimates of those relationships are plotted here over the range of time since cutting from which data were recorded.
Effect of presence of bales
There was no significant effect of the presence of bales on aerial feeders (χ2 = 0.03, 1 df, P = 0.87), gulls (χ2 = 0.09, 1 df, P = 0.76), corvids (χ2 = 3.20, 1 df, P = 0.07), Kestrels (χ2 = 1.50, 1 df, P = 0.22) or pigeons (χ2 = 1.38, 1 df, P = 0.24). There were insufficient data on Pheasants for analysis.
DISCUSSION
This study concurs with earlier studies reporting preferences for short grass by a range of species. These earlier studies showed that corvids, pigeons and Kestrels preferred short grass swards (Perkins et al. Citation2000, Atkinson et al. Citation2005, Whittingham & Devereux Citation2008, Garratt et al. Citation2011), but did not investigate the time period over which the effects of grass cutting attract foraging birds. The key result reported here is the rapid drop off in field use by many bird species after cutting ().
Why do some species prefer shorter grass while others show the reverse pattern?
There are three key factors which affect choice of foraging location: predation risk, food accessibility and food abundance. The ways in which each of these factors is likely to influence the results from our study are considered and discussed.
For many small birds, perceived predation risk is a principle factor affecting which habitats are selected for foraging activities (Whittingham & Evans Citation2004, Whittingham et al. Citation2004, Whittingham & Devereux Citation2008). Predation risk differs depending on escape strategy (fleeing or crypsis). Shorter grass provides easier detection of predators and is preferred by species that flee upon detection of a predator. Long grass provides somewhere to hide and is generally preferred over short grass by those species which use crypsis as an anti-predator strategy, for example Pheasants (Dowell Citation1990).
Prey abundance is an important determinant of patch choice, but is often mediated by accessibility to influence actual food availability and intake rates. For example, a patch containing some prey items which are relatively exposed is likely to be chosen over a patch containing a greater number of prey items which are difficult to access (e.g. due to dense vegetation: Whittingham & Markland Citation2002). However, for one group in our study, prey abundance is likely to be the key factor determining intake rate. Aerial feeders (e.g. swifts and hirundines such as Barn Swallows and House Martins) forage on flying (mainly foliar) invertebrates, which are likely to be more abundant in long, established grassland (Ambrosini et al. Citation2002, Atkinson et al. Citation2004). Cutting can result in a temporary flush of invertebrate availability, and hirundines will often congregate on freshly cut hay or silage (Vickery et al. Citation2001), but our results show this effect to be reasonably short-lived, and there is a significantly lower occurrence of aerial feeders with time since cutting ().
In contrast to aerial feeders, many species of farmland bird feed on surface invertebrates such as beetles, below-ground soil invertebrates such as earthworms and Tipulid spp. (e.g. corvids, gulls), or small mammals (e.g. Kestrels). The intake rate of birds which feed on these prey is determined not just by their abundance but by their accessibility. Experiments with Common Starlings foraging on below-ground invertebrates on grassland swards are useful to demonstrate the principle. Devereux and colleagues Citation(2004) showed that prey intake rates by Starlings are higher on short swards (3 cm) than on longer swards (13 cm), and that this is likely to be due to a combination of greater food accessibility and lower predation risk due to improved detection of predators. In our study we found that both gull and corvid species occurred more often on cut grass. Because of their size, neither of these groups is likely to be concerned with predation risk. Instead they are more likely to be responding to changes in accessibility of their prey, e.g. earthworms Lumbricus spp. Similarly, Kestrels also showed a trend for greater occurrence on shorter swards in line with previous studies (Sheffield et al. Citation2001, Aschwanden et al. Citation2005, Whittingham & Devereux Citation2008, Garratt et al. Citation2011). Kestrels are diurnal hunters (Village Citation1990) which either hover over or perch overlooking grass in order to visually detect their prey, most commonly small mammals such as voles Microtus spp., and small birds (the latter particularly in June/July). However, they will also take large surface and foliar invertebrates, and, more rarely, amphibians and reptiles (Village Citation1990). Reduction of cover is likely to increase accessibility to many of these prey items for Kestrels (Preston Citation1990, Viitala et al. Citation1995, Jacob & Brown Citation2000, Jacob & Hempel Citation2003, Garratt et al. Citation2011).
The preference for shorter swards by Common Wood Pigeons is likely to be due to their preference for grazing on low-growing, small-leaved weeds such as clover Trifolium spp., and they have been shown in several studies to prefer short swards maintained by either mowing, grazing, or trampling (Kenward & Sibly Citation1977, Brough & Bridgman Citation1980, Buckingham & Peach Citation2005).
Why does use of cut grass decline rapidly after cutting?
The occurrence of all species/groups with sufficient post-cut data for analysis decreased as the time since cut increased (). The temporal effects of grass cutting on the foraging behaviour of the birds in our study are likely to be primarily caused by the effects that cutting grass has on prey abundance and distribution. Tall grass supports greater arthropod diversity and abundance than short grass (Morris Citation2000, Vickery et al. Citation2001), and foliar invertebrates are likely to disperse quickly after cutting or are killed by the cutting operation (e.g. Humbert et al. Citation2010), which probably explains the trend shown by the aerial feeders (see above). However, the immediate effects of cutting on soil and surface invertebrates are likely to be less marked, with the most noticeable initial decline in abundance of the newly exposed prey probably due to depletion by foraging birds and other predators. Mowing of the grass will also alter the microclimate of the soil and sward, which is likely to cause invertebrates to disperse and their availability to decline naturally over time (Devereux et al. Citation2006). This decline in prey abundance will result in a decline in foraging activity of those birds that feed on them. Kestrel prey items, such as voles, will also have a relatively high availability in cut grass initially (whether dead or alive) and then decline (Edge et al. Citation1995, C. M. Garratt, unpubl data) as they either move to find cover or are eaten. Many studies have found clear reactions to sudden changes in vegetation height in voles Microtus spp., such as dispersal and decreases in home range size (Hansson Citation1977, Edge et al. Citation1995, Sheffield et al. Citation2001, Jacob & Hempel Citation2003).
Cutting of grass fields on other farms surrounding our study fields could potentially affect our results, but we were unable to control for this as we did not collect information on the timing of cutting in the surrounding landscape. However, we could partially address this issue by examining whether there was a preference for fields that were cut earlier (which are likely to have represented a scarcer resource than later cut fields). Most of the fields on each of our three study farms were cut within one or two weeks of each other (with the exception of one field cut a month early – see Methods and Materials). The date that each field was cut was not significantly related to occurrence of any group except corvids (and a close to significant result for gulls), which tended to make more use of fields that were cut earlier. This suggests that the numbers of cut fields present in the surrounding area affects use of a particular field by corvids and gulls (i.e. the later a field is cut, the higher the likelihood of the availability of other cut fields on and around each farm). However, it is important to note that, although we could not control for this issue entirely, the date of cutting was included in all of the models in the results section. Coordination of the timing of cutting by an individual farmer (or neighbouring farmers) could be used as a management tool to manipulate habitat quality for farmland birds.
Management implications
Conserving wild bird populations is a key conservation objective at a Europe-wide level (Gibbons Citation2000, Gregory et al. Citation2005, European Commission Citation2010) and in response to this, up until June 2010, a UK government target was in place to reverse the declines of the 19 farmland bird species on the ‘Farmland Bird Index’ (FaBI) by 2020. This target was selected by the UK government as a measurable surrogate for assessing the success of its policies, which seek to conserve wildlife, in general, in the English countryside (Gregory et al. Citation2005). Four of the species included in our study are on the FaBI list (namely Rooks, Jackdaws, Wood Pigeons, and Kestrels). Thus our findings suggest management that is directly pertinent to this index, although of the four species Kestrel are the only one whose numbers are still declining.
It is important to note that our study sites did not support many species of conservation concern (with the notable exception of Grey Partridges Perdix perdix, Swifts, Sky Larks, Tree Sparrows Passer montanus, Northern Lapwings Vanellus vanellus L. and Kestrels). However, some of the species which did occur in great enough numbers for analysis are functionally similar to other key species. For example, the corvids and gulls we recorded feed mainly on below-ground invertebrates, and so the patterns we report are likely to apply to other species that feed in a similar way and that are on the UK list of Birds of Conservation Concern (for example, the red-listed Song Thrushes Turdus philomelos and Common Starlings, and the amber-listed Red-billed Choughs Pyrrhocorax pyrrhocorax).
We acknowledge the important point that past studies have shown that some species (not found in our study) are negatively affected by grass cutting due to nest destruction and decreased chick mortality (e.g. Corncrakes Crex crex: Tyler et al. Citation1998; Black-tailed Godwits Limosa limosa: Schekkerman & Bientema Citation2007, Kleijn et al. Citation2010). These species are unlikely to nest in small patches of grassland such as would be found in grass margins (a key Entry Level Scheme management option, Natural England Citation2010a), but our results need to be interpreted in the light of these studies as well as those showing the benefits of grass cutting for some species, and the potential for negative effects needs to be borne in mind when implementing our findings.
The temporal effects shown in this study suggest the need to provide cut grass for foraging at different times throughout the summer months. Our results suggest that providing freshly cut grass patches every 2–3 weeks would be of benefit to a range of different farmland birds, and farmers or landowners could be recommended to carry out multiple cuts on grassland, or stagger the cuts for each field (logistics allowing) to provide a more constant food supply. We suggest our results should be applied in a bespoke manner by land mangers wishing to either deter the use of grasslands (such as on airports) who may wish to retain longer swards (for a fuller exploration of this issue see Brough & Bridgman Citation1980 and Blackwell et al. in review) or those wishing to provide beneficial habitats for foraging birds on farmland by creating a mix of short and long grass in close juxtaposition (Aschwanden et al. Citation2005, Douglas et al. Citation2009, Garratt et al. Citation2011).
ACKNOWLEDGEMENTS
We thank the three farmers for permission to access their grass fields and information regarding field use and management: David Cowton (Carr's Farm), David Gibson (Hall Hill Farm), and Bill Hall (North Farm). Claudia Garratt was funded by a grant from Natural England.
REFERENCES
- Ambrosini , R. , Bolzern , A. M. , Canova , L. , Arieni , S. , Møller , A. P. and Saino , N. 2002 . The distribution and colony size of barn swallows in relation to agricultural land use . J. Appl. Ecol. , 39 : 524 – 534 .
- Aschwanden , J. , Birrer , S. and Jenni , L. 2005 . Are ecological compensation areas attractive hunting sites for common kestrels (Falco tinnunculus) and long-eared owls (Asio otus) . J. Ornithol. , 146 : 279 – 286 .
- Atkinson , P. W. , Buckingham , D. and Morris , A. J. 2004 . What factors determine where invertebrate-feeding birds forage in dry agricultural grasslands? Ibis . 146 ( Suppl. 2 ) : 99 – 107 .
- Atkinson , P. W. , Fuller , R. J. , Vickery , J. A. , Conway , G. J. , Tallowin , J. R.B. , Smith , R. E.N. , Haysom , K. A. , Ings , T. C. , Asteraki , E. J. and Brown , V. K. 2005 . Influence of agricultural management, sward structure and food resources on grassland field use by birds in lowland England . J. Appl. Ecol , 42 : 932 – 942 .
- Barnett , P. R. , Whittingham , M. J. , Bradbury , R. B. and Wilson , J. D. 2004 . Use of unimproved and improved lowland grassland by wintering birds in the UK . Agr. Ecosyst. Environ. , 102 : 49 – 60 .
- Bates , D. , Maechler , M. and Dai , B. 2008 . Lme4: Linear mixed-effects models using S4 classes. R package version 0.999375-28 . Available online at: http://lme4.r-forge.r-project.org/ (accessed 10 October 2010)
- Benton , T. G. , Vickery , J. A. and Wilson , J. D. 2003 . Farmland biodiversity: is habitat heterogeneity the key? . Trends Ecol. Evol. , 18 : 182 – 188 .
- Bibby , C. , Burgess , N. D. and Hill , D. A. 1992 . Bird Census Techniques , London : Academic Press .
- Blackwell , B. F. , DeVault , T. L. , Fernández-Juricic , E. and Dolbeer , R. A. 2009 . Wildlife collisions with aircraft: a missing component of land-use planning on and near airports? . Landscape Urban Plan. , 93 : 1 – 9 .
- Blackwell, B.F., Seamans, T.W., Schmidt, P.M., DeVault, T.L., Belant, J.L., Whittingham, M.J., Martin, J.A. & Fernandez-Juricic, E. (in review). Airports, grasslands, and birds: a framework for management amidst conflicting priorities
- Brough , T. and Bridgman , C. J. 1980 . An evaluation of long grass as a bird deterrent on British airfields . J. Appl. Ecol. , 17 : 243 – 253 .
- Buckingham , D. L. and Peach , W. J. 2005 . The influence of livestock management on habitat quality for farmland birds . Anim. Sci , 81 : 199 – 203 .
- Buckingham , D. L. , Atkinson , P. W. and Rook , A. J. 2004 . Testing solutions in grass-dominated landscapes: a review of the current research . Ibis , 146 ( Suppl. 2 ) : 163 – 170 .
- Buckingham , D. L. , Peach , W. J. and Fox , D. S. 2006 . Effects of agricultural management on the use of lowland grassland by foraging birds . Agr. Ecosyst. Environ. , 112 : 21 – 40 .
- Crawley , M. J. 1993 . GLIM for Ecologists Blackwell Scientific Publications, Oxford
- Crawley , M. J. 2007 . The R Book , 1 , Chichester : Wiley & Sons Ltd .
- Devereux , C. L. , McKeever , C. U. , Benton , T. G. and Whittingham , M. J. 2004 . The effect of sward height and drainage on Common Starlings Sturnus vulgaris and Northern Lapwings Vanellus vanellus foraging in grassland habitats . Ibis , 146 ( Suppl. 2 ) : 115 – 122 .
- Devereux , C. L. , Whittingham , M. J. , Krebs , J. R. , Fernández-Juricic , E. and Vickery , J. A. 2006 . What attracts birds to newly mown pasture? Decoupling the action of mowing from the provision of short swards . Ibis , 148 : 302 – 306 .
- Donald , P. F. , Green , R. E. and Heath , M. F. 2001 . Agricultural intensification and the collapse of Europe's farmland bird populations . Proc. R. Soc. Lond. B. , 268 : 25 – 29 .
- Donald , P. F. , Sanderson , F. J. , Burfield , I. J. and van Bommel , F. P.J. 2006 . Further evidence of continent-wide impacts of agricultural intensification on European farmland birds, 1990–2000 . Agr. Ecosyst. Environ. , 116 : 189 – 196 .
- Douglas , D. J.T. , Vickery , J. A. and Benton , T. G. 2009 . Improving the value of field margins as foraging habitat for farmland birds . J. Appl. Ecol , 46 : 353 – 362 .
- Dowell , S. D. 1990 . “ The development of anti-predator responses in grey partridges and common pheasant ” . In Pheasants of Asia 1989 , Edited by: Hill , D. A. , Garson , P. J. and Jenkins , D. 193 – 199 . Reading , , UK : World Pheasant Association .
- Edge , W. D. , Wolff , J. O. and Carey , R. L. 1995 . Density-dependent responses of gray-tailed voles to mowing . J. Wildlife Manage. , 59 : 245 – 251 .
- European Commission . 2010 . “ Directive 2009/147/EC (codified version of Directive 79/409/EEC as amended) ” . In The Birds Directive Available online at: http://ec.europa.eu/environment/nature/legislation/birdsdirective/index_en.htm (accessed 15 November 2010)
- Garratt , C. G. , Hughes , M. , Eagle , G. , Fowler , T. , Grice , P. V. and Whittingham , M. J. 2011 . Foraging habitat selection by breeding kestrels Falco tinnunculus on lowland farmland in England . Bird Study , 58 : 90 – 98 .
- Gibbons , D. W. 2000 . Development of pan-European breeding bird monitoring . Ring , 22 : 25 – 33 .
- Gregory , R. D. , Van Strien , A. , Vorisek , P. , Gmelig Meyling , A. W. , Noble , D. G. , Foppen , R. P.B. and Gibbons , D. W. 2005 . Developing indicators for European birds . Phil. Trans. R. Soc. B. , 360 : 269 – 288 .
- Hansson , L. 1977 . Spatial dynamics of Field Voles Microtus agrestis in heterogeneous landscapes . Oikos , 29 : 539 – 544 .
- Humbert , J. Y. , Ghazoul , J. , Sauter , G. J. and Walter , T. 2010 . Impact of different meadow mowing techniques on field invertebrates . J. Appl. Entomol. , 134 : 592 – 599 .
- Jacob , J. and Brown , J. S. 2000 . Microhabitat use, giving-up densities and temporal activity as short- and long-term anti-predator behaviours in common voles . Oikos , 91 : 131 – 138 .
- Jacob , J. and Hempel , N. 2003 . Effects of farming practices on spatial behaviour of common voles . J. Ethol. , 21 : 45 – 50 .
- Kenward , R. E. and Sibly , R. M. 1977 . A woodpigeon Columba palumbus feeding preference explained by a digestive bottle-neck . J. Appl. Ecol. , 14 : 815 – 826 .
- Kleijn , D. , Schekkerman , H. , Dimmers , W. J. , Van Kats , R. J.M. , Melman , D. and Teunissen , W. A. 2010 . Adverse effects of agricultural intensification and climate change on breeding habitat quality of Black-tailed Godwits Limosa l. limosa in the Netherlands . Ibis , 152 : 475 – 486 .
- Lima , S. L. and Dill , L. M. 1990 . Behavioral decisions made under the risk of predation: a review and prospectus . Can. J. Zool. , 68 : 619 – 640 .
- Moorcroft , D. , Whittingham , M. J. , Bradbury , R. B. and Wilson , J. D. 2002 . The selection of stubble fields by wintering granivorous birds reflects vegetation cover and food abundance . J. Appl. Ecol. , 39 : 535 – 547 .
- Morris , M. G. 2000 . The effects of structure and its dynamics on the ecology and conservation of arthropods in British grasslands . Biol. Conserv. , 95 : 129 – 142 .
- Natural England . 2010a . Entry Level Stewardship Environmental Stewardship Handbook , 3 February 2010. Available online at: www.naturalengland.org.uk (accessed 12 October 2010)
- Natural England . 2010b . Higher Level Stewardship Environmental Stewardship Handbook , 3 February 2010. Available online at: www.naturalengland.org.uk (accessed 12 October 2010)
- Perkins , A. J. , Whittingham , M. J. , Bradbury , R. B. , Wilson , J. D. , Morris , A. J. and Barnett , P. R. 2000 . Habitat characteristics affecting use of lowland agricultural grassland by birds in winter . Biol. Conserv. , 95 : 279 – 294 .
- Preston , C. R. 1990 . Distribution of raptor foraging in relation to prey biomass and habitat structure . Condor , 92 : 107 – 112 .
- R Development Core Team . 2009 . R: A Language and Environment for Statistical Computing R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0. http://www.R-project.org
- Schekkerman , H. and Beintema , A. J. 2007 . Abundance of invertebrates and foraging success of black-tailed godwit Limosa limosa chicks in relation to agricultural grassland management . Ardea , 95 : 39 – 54 .
- Sheffield , L. M. , Crait , J. R. , Edge , D. W. and Wang , G. 2001 . Response of American kestrels and gray-tailed voles to vegetation height and supplemental perches . Can. J. Zool. , 79 : 380 – 385 .
- Stephens , D. W. and Krebs , J. R. 1986 . Foraging Theory , Princeton , NJ : Princeton University Press .
- Tyler , G. A. , Green , R. E. and Casey , C. 1998 . Survival and behaviour of Corncrake Crex crex chicks during the mowing of agricultural grassland . Bird Study , 45 : 35 – 50 .
- Vickery , J. A. , Tallowin , J. R. , Feber , R. E. , Asteraki , E. J. , Atkinson , P. W. , Fuller , R. J. and Brown , V. K. 2001 . The management of lowland neutral grasslands in Britain: effects of agricultural practices on birds and their food resources . J. Appl. Ecol. , 38 : 647 – 664 .
- Viitala , J. , Korpimäki , E. , Palokangas , P. and Kolvula , M. 1995 . Attraction of kestrels to vole scent marks visible in ultraviolet light . Nature , 373 : 425 – 427 .
- Village , A. 1990 . The Kestrel , London : Poyser .
- Whittingham , M. J. 2007 . Will agri-environment schemes deliver substantial biodiversity gain, and if not why not? . J. Appl. Ecol. , 44 : 1 – 5 .
- Whittingham , M. J. and Devereux , C. L. 2008 . Changing grass height alters foraging site selection by wintering farmland birds . Basic Appl. Ecol. , 9 : 779 – 788 .
- Whittingham , M. J. and Evans , K. L. 2004 . The effects of habitat structure on predation risk of birds in agricultural landscapes . Ibis , 146 : 210 – 220 .
- Whittingham , M. J. and Markland , H. M. 2002 . The influence of substrate on the functional response of an avian granivore and its implications for farmland bird conservation . Oecologia , 130 : 637 – 644 .
- Whittingham , M. J. , Butler , S. J. , Quinn , J. L. and Cresswell , W. 2004 . The effect of limited visibility on vigilance behaviour and speed of predator detection: implications for the conservation of granivorous passerines . Oikos , 106 : 377 – 385 .
- Whittingham , M. J. , Stephens , P. A. , Bradbury , R. B. and Freckleton , R. P. 2006 . Why do we still use stepwise modelling in ecology and behaviour? . J. Anim. Ecol. , 75 : 1182 – 1189 .