Abstract
Capsule Immature Short-toed Snake Eagles hatched in Spain leave their Sahelian wintering grounds to spend the summer in northern Africa.
The importance of data on the non-breeding fraction of a bird population for understanding demography and for determining conservation strategies is well recognized, especially in long-lived species such as raptors (Ferrer & Harte Citation1997, Kenward et al. Citation2000, Penteriani et al. Citation2005, Citation2011, Soutullo et al. Citation2008a). Nevertheless, little is known of the behaviour of immature individuals of migratory raptor species during the breeding season (but see Sergio et al. Citation2009). Knowledge about this phase of a raptor's life cycle could be very important for ensuring a high population level or for ensuring the conservation of high-quality breeding areas that may be occupied at a future time (Newton Citation1979).
GPS-satellite telemetry is a useful tool for indicating the movement patterns of large birds. To date, most of the studies have focused on analysing the routes of migratory species (Klaassen et al. Citation2008, Garcia-Ripollés et al. Citation2010) or on juvenile dispersal of resident species (Soutullo et al. Citation2008b, Cadahia et al. Citation2010). In contrast, the ranging behaviour of non-breeding individuals of migratory species has been largely neglected.
Short-toed Snake Eagles Circaetus gallicus are migratory raptors that breed in Europe and northern Africa, spending the winter in sub-Saharan Africa (Cramp & Simmons Citation1980). Many aspects of their biology have already been studied, including reproduction (Bakaloudis et al. Citation2005), habitat selection (López-Iborra et al. Citation2011) and migration (Agostini et al. Citation2002, Muñoz et al. Citation2010, Pavón et al. Citation2010). Nonetheless, knowledge on summer ranging behaviour of non-breeding individuals of this species is lacking, although traditionally it has been supposed that non-breeding birds spend the summer in the same areas where they winter, i.e. south of the Sahara desert (Campora & Cattaneo Citation2005, Newton Citation2008). Like other long-lived raptor species, Short-toed Snake Eagles are not able to breed at least until the fourth calendar year (Campora & Cattaneo Citation2005). In this study, we use satellite telemetry to describe movements and ranging behaviour of immature (second and third calendar-year) Short-toed Snake Eagles.
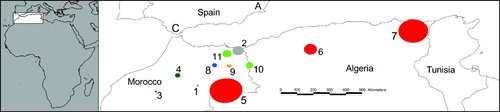
Nine Short-toed Snake Eagle nestlings were tagged with satellite transmitters a few days before fledging. Six individuals were tagged with Microwave Telemetry 45-g solar/GPS transmitters (see Pavón et al. Citation2010 for more details) in the province of Alicante (southeastern Spain; ‘A’ in ) in 2008 and 2009. The other three birds were tagged with North Star 30-g solar/GPS transmitters in the province of Cadiz (southern Spain; ‘C’ in ) in 2009. Overall, we recorded data from seven summering events involving six individuals; all birds were tracked during their second calendar-year, with one of them being tracked during two consecutive years (thus, also in its third calendar-year; #80423, see ). For the remaining tagged birds, no data were obtained due to failure of equipment (one bird) and because two birds died during their stay in the wintering grounds in the Sahel (see also Pavón et al. Citation2010).
Table 1. Histories of the seven summering events of six Short-toed Snake Eagles. ‘Spring’ and ‘Autumn’ are the dates of migration onsets. ‘Period’, ‘Size’ and ‘Distance’ (ranging distance in km) are given for each one of the 11 areas identified with the kernel analyses.
Figure 1. Staging areas of immature Short-toed Snake Eagles during summer. Each number corresponds to a different staging area (cross-referenced in ). ‘C’ (Cadiz) and ‘A’ (Alicante) are the nesting areas where the birds were tagged.
Our aim was to analyse these eagles' movements occurring between the end of the spring migration and the beginning of the autumn migration, hereafter called ‘summering’. We only considered locations recorded above 31°N, because, above this latitude, birds started to perform non-directional movements, which were interpreted as the end of the northwards spring migration. We checked for the existence of individual staging areas during this period; these staging areas can be defined as individual areas where eagles' movements tend to concentrate and are short and non-directional. To identify the areas, we calculated the 50% fixed kernel density contours (Worton 1989) of all locations obtained for every summering event, using the Animal Movement v2.04 Extension for Arcview 3.2 (Hooge & Eichenlaub Citation1997). To calculate the actual size of these areas (in km2), we projected the generated kernel contours using the ‘Projector!’ extension, and an Equal-Area Cylindrical transformation.
Due to differences in the duty cycle and technical performance of the transmitters, there were differences in the frequency of locations recorded (range: 2–7 locations per day, ). Hence, we also performed the same kernel analyses with a reduced data set in which the number of locations per day was similar across the different summering events. We found that the sizes of the areas identified with the reduced data set were 23.7% larger (all areas pooled), but there were no differences in the number of identified staging areas for every summering event and, thus, we here present the results based on the complete data set.
Besides the size of every staging area, we also calculated a ‘ranging distance’, in order to give a linear measurement to the ranging behaviour for each area previously identified with the kernel analyses (see Haworth et al. Citation2006). To this end, we calculated the centre of every staging area as the harmonic mean of all the locations received during the whole period between the first and the last locations encompassed within every individual staging area (Dixon & Chapman Citation1980, Kenward et al. Citation2001). Then, we measured the distances between the centre of a given area and the same above-mentioned locations to calculate an average ranging distance for every staging area (). Additionally, these centres were also used to calculate the distances between the different areas used by the same individual.
Immature Short-toed Snake Eagles started the northwards spring migration mainly in April (mean onset date: 20 April; ). None of the birds returned to European breeding areas of the species during their second calendar year (this also applied to the individual tracked in two consecutive years), but spent the summer in northern Africa. The eagles were mostly located in northeastern Morocco, but also in northern Algeria (). According to the 50% fixed kernel contours, birds had between one and three individual staging areas for every summering event (). shows the number of individual areas identified by kernel analyses for every summering event as well as size and ranging distances. Individual summering areas had a mean size of 5153 km2 (n = 11; sd = 8652). The birds' overall mean ranging distance within each individual staging area was 13.55 km (n = 11; sd = 23.42). Birds left their staging areas in northern Africa heading back to the Sahel mainly in early September (mean date of autumn migration onset: 8 September). Therefore, immature eagles spent between four and six months at their summering areas ().
In four summering events Short-toed Snake Eagles stayed in a single staging area (individuals; #80423 for two years, #85674, #96231); one bird (#80421) used two areas; and in two cases eagles used three different areas (individuals; #80424, #96233). The transmitter of bird #96233 worked intermittently during July 2010. This bird left the second area and stayed for about five weeks in a third restricted area (data not depicted in ) but, due to irregularity in data reception, the kernel analysis did not identify this area. Hence, we assume that this individual used three areas (although we do not have accurate data to estimate the ranging behaviour in the third area).
Regarding the distances between the different staging areas from the same individual during the same summering event, the two areas used by #80421 were 337 km apart, whereas for #80424 there were 498 km between the first and the second staging areas and 538 km between the second and the third. The first two areas used by #96233 were 142 km apart, with July's locations concentrated in a third area located close to the Mediterranean coast in northern Morocco, at about 80 km from the second area. The only bird tracked for two consecutive years (#80423) did not show site fidelity to summering areas, having a single but different staging area in each year and these were 165 km apart.
Immature Short-toed Snake Eagles left their wintering Sahelian grounds by mid-April, which is rather late in the season compared to adults (which arrive in their European breeding grounds in early March; Cramp & Simmons Citation1980). After crossing the Sahara Desert, birds settled in Morocco and Algeria, thus not returning to Europe either in their second or third summer. This delayed migration and the sojourn in these areas may be due to the fact that immature eagles are not motivated to return to European breeding areas because they are not able to breed during the second or the third calendar-year. For this reason, and because of their behaviour during this period (using different roosting sites and sometimes also several staging areas), we discount the possibility that these birds bred in northern Africa during the period considered here. Immature Short-toed Snake Eagles may have found suitable foraging areas along the migration route and in these regions intraspecific competition was probably lower than at European breeding grounds. Furthermore, these eagles avoided crossing the Strait of Gibraltar, which is likely to represent a highly energy-demanding challenge for a soaring raptor (Bildstein Citation2006). These summering areas are within the breeding range of the species (Thevenot et al. Citation2003) and future settling of these individuals as breeders, when they reach sexual maturity, cannot be excluded. Nevertheless breeding density is probably lower than in Europe because of the scarcity of woodlands that the species needs for nesting (López-Iborra et al. Citation2011).
There was individual variation regarding the use of summer staging areas, with some individuals settling in a single small area and others using multiple areas, including one bird (#80424) which travelled more than 1000 km across three different areas (). This could be related to spatial and temporal variation in food abundance and local conditions, but specific studies are needed to determine this.
Data concerning the return rate of immature Short-toed Snake Eagles to breeding areas in Europe are scarce. In Spain and Italy, a few immature Short-toed Snake Eagles (second and third calendar-year) are observed migrating every spring, mainly late in the season, between April and July (Fundación Migres, unpub. data, Finlayson Citation1992, Garcia & Bensusan Citation2006, Premuda et al. 2010). In Italy, a small but regular summer communal roost, containing mainly non-breeding immature birds, has been reported (Premuda 2010). On the other hand, a juvenile tracked from France spent the summer in Mali, south of the Sahara (Cavallin et al. Citation2007). Given that all individuals tracked in this study showed the same strategy (i.e. leaving their wintering grounds but not reaching European breeding areas), we may argue that this is a conservative behavioural strategy that may act to maintain a high survival rate, probably avoiding intraspecific competition with breeding adults. In this case, a higher return rate would be expected in Short-toed Snake Eagles hatched in lower-density populations in comparison with the Spanish one, such as the Italian population (Agostini & Mellone Citation2008).
In other migratory long-lived raptor species, such as Ospreys Pandion haliaetus, Egyptian Vultures Neophron percnopterus and Honey Buzzards Pernis apivorus, immature birds stay in their wintering grounds after their first winter, returning to the natal areas only once they are sexually mature (Hake et al. Citation2001, Meyburg et al. Citation2004, Panuccio & Agostini Citation2006). Short-toed Snake Eagles tagged in southern Spain chose not visit their natal area and moved to regions that, despite being within the breeding range of the species, probably hold a low population density. Their behaviour is probably more dictated by survival in the short term, rather than by the need to explore potential breeding sites. According to Newton Citation(2008), some individuals of long-distance migratory bird species only cover a part of their complete journey between their wintering and breeding areas, spending the summer at en-route sites. Probably Short-toed Snake Eagles moved northwards, like the majority of Palaearctic migrants and since they found en-route sites with a high density of food they stopped there. Short-toed Snake Eagles are specialist predators of reptiles (Gil-Sánchez & Pleguezuelos Citation2001) and northern Africa is an area with one of the highest reptile abundances within the Mediterranean region (Cox et al. Citation2006).
Our study indicates that conservation strategies for migratory long-lived species should not be solely focused on breeding and wintering grounds (Martin et al. Citation2007), but should consider also those events occurring in non-breeding summering areas, which could have important consequences at the population level (Penteriani et al. Citation2005, Soutullo et al. Citation2008a). Further research should investigate the importance of this summering behaviour in immature Short-toed Snake Eagles, especially its influence on lifetime reproductive success.
ACKNOWLEDGEMENTS
Funding was provided by Diputación Provincial de Alicante and by Fundación Migres through a collaboration project with the Asociación Eólica de Tarifa. The Servicio de Biodiversidad from the Conselleria de Medio Ambiente, Agua, Urbanismo y Vivienda (Generalitat Valenciana), the Consejeria de Medio Ambiente–Junta de Andalucia and the CAS Retin – Ministerio de Defensa – provided research permissions. We are grateful to Alejandro Izquierdo, Alejandro de la Vega, Sergio Morán, Pepe García, José Aragoneses, Miguel Gallardo, Juan Manuel Pérez-García, Pascual López, Carlos Torralvo, Javier Elorriaga, Rafael Sánchez-Carrión and José Maria García who helped during the fieldwork, and to Edward Kluen for checking the English of the manuscript. We also thank two anonymous referees for their useful comments on an earlier draft. RL has a postdoctoral grant (reference 10/12-C) co-funded by Consejería de Educación y Ciencia (Junta de Comunidades de Castilla–La Mancha) and the European Social Fund. UM is supported by a FPU grant of the Spanish Ministry of Education (AP2008-0947). BY's research is financed by a grant from Fundacion Migres. DP was awarded a grant by the Research Foundation of the University of Helsinki (Helsingin Yliopiston tiedesäätiö).
REFERENCES
- Agostini , N. and Mellone , U. 2008 . Does migration flyway of Short-toed Snake-Eagles breeding in central Italy reflect the colonization history? . J. Raptor Res. , 42 : 158 – 159 .
- Agostini , N. , Baghino , L. , Coleiro , C. , Corbi , F. and Premuda , G. 2002 . Circuitous autumn migration in the Short-toed Eagle (Circaetus gallicus) . J. Raptor Res. , 36 : 111 – 114 .
- Bakaloudis , D. E. , Vlachos , C. G. and Holloway , G. J. 2005 . Nest spacing and breeding performance in Short-toed Eagle Circaetus gallicus in northeast Greece . Bird Study , 52 : 330 – 338 .
- Bildstein , K. L. 2006 . Migrating Raptors of the World , Ithaca, NY : Cornell University Press .
- Cadahìa , L. , López-López , P. , Urios , V. and Negro , J. J. 2010 . Satellite telemetry reveals individual variation in juvenile Bonelli's eagle dispersal areas . Eur. J. Wildl. Res. , 56 : 923 – 930 .
- Campora , M. and Cattaneo , G. 2005 . Ageing and sexing Short-toed Eagles . Br. Birds , 98 : 370 – 376 .
- Cavallin , P. , Ventroux , J. , Chevallier , D. and Baillon , F. 2007 . Suivi par balise Argos: de la Vienne au Mali . La Plume du Circaète , 6 : 4 – 5 .
- Cox , N. , Chanson , J. and Stuart , S. 2006 . The Status and Distribution of Reptiles and Amphibians of the Mediterranean Basin , Edited by: Cox , N. , Chanson , J. and Stuart , S. Gland, Switzerland and Cambridge, UK : IUCN .
- Cramp , S. and Simmons , K. E.L. 1980 . The Birds of the Western Palaearctic , Oxford : Oxford University Press .
- Dixon , K. R. and Chapman , J. A. 1980 . Harmonic mean measure of animal activity areas . Ecology , 61 : 1040 – 1044 .
- Ferrer , M. and Harte , M. 1997 . Habitat selection by immature Spanish imperial eagles during the dispersal period . J. Appl. Ecol. , 34 : 1359 – 1364 .
- Finlayson , C. 1992 . Birds of the Strait of Gibraltar , London : T. & A.D. Poyser .
- Garcia , E. F.J. and Bensusan , K. J. 2006 . Northbound migrant raptors in June and July at the Strait of Gibraltar . Br. Birds , 99 : 569 – 575 .
- García-Ripollés , C. , López-López , P. and Urios , V. 2010 . First description of migration and wintering of adult Egyptian Vultures Neophron percnopterus tracked by GPS satellite telemetry . Bird Study , 57 : 261 – 265 .
- Gil-Sánchez , J. M. and Pleguezuelos , J. M. 2001 . Prey and prey-size selection by the short-toed eagle (Circaetus gallicus) during the breeding season in Granada (south-eastern Spain) . J. Zool. Lond. , 255 : 131 – 137 .
- Hake , M. , Kjellén , N. and Alerstam , T. 2001 . Satellite tracking of Swedish Ospreys Pandion Haliaetus: autumn migration routes and orientation . J. Avian Biol. , 32 : 47 – 56 .
- Haworth , P. F. , McGrady , M. J. , Whitfield , D. P. , Fielding , A. H. and McLeod , D. R.A. 2006 . Ranging distance of resident Golden Eagles Aquila chrysaetos in western Scotland according to season and breeding status . Bird Study , 53 : 265 – 273 .
- Hooge , P. N. and Eichenlaub , B. 1997 . Animal Movement Extension for ArcView , Anchorage, USA : Alaska Science Centre, Biological Science Office, US Geological Survey .
- Kenward , R. E. , Walls , S. S. , Hodder , K. H. , Pahkala , M. , Freeman , S. N. and Simpson , V. R. 2000 . The prevalence of non-breeders in raptor populations: evidence from rings, radio-tags and transect surveys . Oikos , 91 : 271 – 279 .
- Kenward , R. E. , Clarke , R. T. , Hodder , K. H. and Walls , S. S. 2001 . Density and linkage estimators of home range: nearest-neighbor clustering defines multinuclear cores . Ecology , 82 : 1905 – 1920 .
- Klaassen , R. H.G. , Strandberg , R. , Hake , M. and Alerstam , T. 2008 . Flexibility in daily travel routines causes regional variation in bird migration speed . Behav. Ecol. Sociobiol. , 62 : 1427 – 1432 .
- López-Iborra , G.M. , Limiñana , R. , Pavón , D. and Martínez-Pérez , J.E. 2011 . Modelling the distribution of short-toed eagle (Circaetus gallicus) in semi-arid Mediterranean landscapes: identifying important explanatory variables and their implications for its conservation . Eur. J. Wildl. Res , 57 : 83 – 93 .
- Martin , T. G. , Chadès , I. , Arcese , P. , Marra , P. P , Possingham , H. P. and Norris , D. R. 2007 . Optimal Conservation of Migratory Species . PLoS ONE , 2 ( 8 ) : e751
- Meyburg , B. U. , Gallardo , M. , Meyburg , C. and Dimitrova , E. 2004 . Migrations and sojourn in Africa of Egyptian Vultures (Neophron percnopterus) tracked by satellite . J. Ornithol. , 145 : 273 – 280 .
- Muñoz , A. R. , Toxopeus , B. , Elorriaga , J. , Gonzalez , J. M. and Yáñez , B. 2010 . First record of a communal roost of Short-toed Eagles Circaetus gallicus . Ibis , 152 : 173 – 175 .
- Newton , I. 1979 . Population Ecology of Raptors , London : T. & A.D. Poyser .
- Newton , I. 2008 . The Migration Ecology of Birds , London : Academic Press .
- Panuccio , M. and Agostini , N. 2006 . Spring passage of second-calendar-year Honey-buzzards at the Strait of Messina . Br. Birds , 99 : 95 – 97 .
- Pavón , D. , Limiñana , R. , Urios , V. , Izquierdo , A. , Yáñez , B. , Ferrer , M. and de la Vega , A. 2010 . Autumn migration of juvenile Short-toed Eagles Circaetus gallicus from southeastern Spain . Ardea , 98 : 113 – 117 .
- Penteriani , V. , Otalora , F. , Sergio , F. and Ferrer , M. 2005 . Environmental stochasticity in dispersal areas can explain the ‘mysterious’ disappearance of breeding populations . Proc. R. Soc. B , 272 : 1265 – 1269 .
- Penteriani , V. , Ferrer , M. and Delgado , M. M. 2011 . Floater strategies and dynamics in birds, and their importance in conservation biology: towards an understanding of nonbreeders in avian populations . Anim. Cons. , 14 : 233 – 241 .
- Premuda , G. 2010 . Trends at a roost of Short-toed Eagles Circaetus gallicus over ten years . Avocetta , 34 : 63 – 64 .
- Premuda , G. , Baghino , L. , Gustin , M. and Borioni , M. 2010 . Migration of immature Short-toed Eagles Circaetus gallicus through the Central Mediterranean: a survey in spring 2007 . Avocetta , 34 : 65 – 68 .
- Sergio , F. , Blas , J. and Hiraldo , F. 2009 . Predictors of floater status in a long-lived bird: a cross-sectional and longitudinal test of hypotheses . J. Anim. Ecol. , 78 : 109 – 118 .
- Soutullo , A. , López-López , P. and Urios , V. 2008a . Incorporating spatial structure and stochasticity in endangered Bonelli's eagle's population models: implications for conservation and management . Biol. Cons. , 141 : 1013 – 1020 .
- Soutullo , A. , Urios , V. , Ferrer , M. and López-López , P. 2008b . Habitat use by juvenile Golden Eagles Aquila chrysaetos in Spain . Bird Study , 55 : 236 – 240 .
- Thevenot , M. , Vernon , J. D.R. and Bergier , P. 2003 . The Birds of Morocco , British Ornithologist Union Checklist Series 20
- Worton, B.J. 1989. Kernel methods for estimating the utilization distribution in home-range studies. Ecology 70: 164–168.