Abstract
Capsule The majority of Robins in Iberia are sedentary and stay in their nesting areas despite the presumed increased competition that results from the seasonal flooding of the area by conspecific migrants.
Aims To evaluate if Robins in Iberia are displaced from their nesting areas by the presence of large numbers of competitors originating from higher latitudes.
Methods Data from constant effort ringing sites were used to estimate the proportion of Robins that are strictly sedentary and the seasonal variation in Robin numbers. Hydrogen stable isotope ratios (δ2H) in feathers, and sex ratios determined through molecular techniques, allowed an insight into the numerical importance of invading Robins from higher latitudes.
Results Robins were more numerous outside the breeding season, with clear influxes in autumn. Around half of the locally nesting Robins were captured during winter, indicating they are truly sedentary. δ2H in feathers suggests that a wide mixture of Robins from all European latitudes were present in winter, and these data, together with sex ratios, suggest that there may be similar numbers of local and foreign individuals in winter at the main study site.
Conclusion Despite the occupation of their range by migrants, local birds are largely sedentary.
For many widely distributed bird species in the northern hemisphere, migratory and resident populations share common wintering grounds in the southern part of their range (Newton & Dale Citation1996). This raises questions about the way individuals from different geographic origins co-exist in a scenario of presumed increased intraspecific competition, resulting from the co-occurrence of several populations in the same areas (Cox Citation1968, Greenberg Citation1986, Pérez-Tris & Tellería Citation2002a). Such questions have the potential to reveal insights into the evolution and maintenance of migratory systems and for the regulation of different populations (Bell Citation2005, Newton Citation2008).
Few studies have examined this issue at the local and individual level. For example, while it is often assumed that southern populations are mostly sedentary (with backing-up evidence from ringing recoveries), it remains unclear whether southern individuals are displaced by migratory conspecifics (for example, by moving to different habitats), or whether they manage to secure the winter sites of their choice despite increased levels of intraspecific competition.
Studies of Blackcaps Sylvia atricapilla and European Robins Erithacus rubecula (hereafter Robins) have documented that local populations largely hold onto their nesting habitats, while migrants predominate in habitats where no locals breed. While this pattern of (partial) population segregation has been clearly established (Tellería et al. Citation2001, Pérez-Tris & Tellería Citation2002a, Tellería & Pérez-Tris Citation2004), the studies inferred the breeding origins of previously unmarked individuals from morphological data and so responses at the individual level are uncertain (e.g. resident birds could still disperse within/between breeding habitats).
Robins are small (ca. 15–19 g) passerines with a range of movement strategies from year-round residents to long-distance migrants. They are common breeders in most of Iberia. In autumn, Robins from across much of Europe, particularly from the large Scandinavian and central European populations, move south to winter in Iberia and other Mediterranean areas (e.g. Cramp Citation1988, Bueno Citation1998, Remisiewicz Citation2002). Both migratory and resident birds often defend individual territories during the non-breeding season, while others behave as floaters. In mostly non-migratory populations, some adult Robins move from the nesting territories once the reproductive season is complete, either just before or just after the post-breeding moult (Cramp Citation1988). Juveniles can settle into winter territories either before or after the adults (Cramp Citation1988). During winter in Iberia, Robins feed on a mixture of invertebrates and fruits such as berries and acorns (Herrera Citation1977, Citation1998).
A recent study in mountain habitats of SE Spain showed that, in an area where Robins are present year-round, nesting individuals seasonally disappeared during the autumn–winter and were replaced by conspecifics from more migratory populations (Domínguez et al. Citation2007). These findings raise the question of whether southern resident populations generally are truly sedentary, or whether they get shifted around as a result of increased intraspecific competition caused by the arrival of migrants and by changes in available resources. Ringing data suggest that individual Iberian Robins can be strictly sedentary, while populations from more seasonal environments, such as mountains, show a greater tendency to move (Pérez-Tris & Tellería Citation2002b). However, there is little information on the relative numbers of truly sedentary birds in Iberian habitats. Could their spatial ecology be more similar to Robin populations in Ireland and Great Britain, where little seasonal flooding takes place and where many Robins stay on the same site all year round (Burkitt Citation1926, Lack Citation1943, Jackson Citation1958, Wernham et al. Citation2002), or is the pattern described by Dominguez et al. (2007) the one that more widely applies to Iberian individuals?
In the present study we determined capture rates, sex ratios and stable isotope ratios in the feathers of nesting and wintering populations of Robins in Iberia. We show that, despite being seasonally confronted by large numbers of incoming foreign individuals, a large proportion of Iberian Robins remain on their breeding territories over winter. Previous studies of partially migrant Robin populations also found that altitude, or type of habitat of the nesting areas, can have a strong influence on the proportion of individuals that display a truly sedentary behaviour (Adriaensen & Dhondt Citation1990, Pérez-Tris & Tellería Citation2002b). In light of this we also assessed whether Robins from different sites vary in their propensity to move away from breeding territories in winter.
MATERIALS AND METHODS
Constant effort sites
Data were obtained from Bird Ringers (Bird Banders) participating in the Portuguese Constant Effort Site bird ringing programme, in which ringers trap birds regularly (about every 2 weeks) using mist-nets at specific study sites. Net placement and capture effort are kept constant in trapping sessions that are regularly distributed within the annual cycle. Trapped birds are ringed and recaptures recorded. Where possible, the age (juvenile versus adult) of the trapped individuals is determined. Brood-patch development is also recorded, which, in the case of Robins, allows the sexing of some individuals during the breeding season. The occurrence of wing-moult is also noted. For the present study, we used data from five constant effort sites (CES) where Robins were known to breed. Sites covered a range of latitudes, altitudes, and habitat types, and had been in continuous operation over a period of at least 36 months ().
Table 1. Location, main habitat, and sampling period in each of the Constant Effort Sites (CES) included in this study.
Details on the location and sampling period of each CES are presented in . The main study site was at Mata da Margaraça, Serra do Açor, an area covered mostly by deciduous forest of oaks Quercus robur and chestnut trees Castanea sativa, with an undergrowth of several evergreen bushes, including Arbutus unedo and Virburnum tinus. Robins feed on the fruits of Virburnum in autumn and winter. Other study sites were: Parque Biológico de Gaia, a large suburban park with open lawns and woodland patches, mostly deciduous (Quercus, Castanea, Alnus), and diverse shrubs including Rubus spp., Sambucus nigra and Frangula alnus; Paul da Madriz, a wetland margin with willows Salix atrocinerea and brambles Rubus spp. fringing a large reedbed; Carvalhal da Azóia, an area of old abandoned orchards bordered with oaks Quercus faginea, pines Pinus pinaster and bramble edges; and Paul de Tornada, a wetland margin with willows and brambles fringing a reedbed area.
Complementary sampling
To compare the breeding and the wintering Robin communities, data from constant effort programs were supplemented at Margaraça by trapping birds with mist-nets and baited spring-traps in January and June 2008. This took place after the cessation of the constant effort trapping at this site, and it did not interfere with the CES programme. Note that this sampling was made for purposes other than recapturing previously marked individuals and birds captured on these occasions were not used to study individual behaviour, but rather to assess social composition at the population level. Feathers (innermost secondary) and blood samples were collected. They were used for stable isotope analysis and for molecular sexing respectively. Because Robins grow secondary feathers on the nesting grounds and very rarely lose them accidentally (which would imply replacement growth), the isotope composition of the metabolically inert feather keratin reflects that of the breeding area. The hydrogen isotope (deuterium) ratios (δ2H) in rainwater (and in bird feathers) show a strong gradient across Europe, with a depletion occurring in a northeast direction (Hobson et al. Citation2004). Hence hydrogen isotope ratios provide a useful tool for avian migration research (e.g. Bearhop et al. Citation2005), as they can differentiate the geographical origins of birds.
We also took feather samples for isotope analyses from Robins from northern Europe, to help put isotope values from Margaraça into context. Robins were sampled during migration through the southern tip of Sweden at Falsterbo (55° 23′ N, 12° 50′ E), where migrant individuals originating from all over Sweden and (less so) from Finland occur during autumn (Karlsson et al. Citation1988). There have been many recoveries in Portugal of Robins which orginate in Scandinavia, and these are likely to contribute to a large proportion of the wintering birds in Portugal (Bueno Citation1998, Catry et al. Citation2010). Trapping took place at Falsterbo on five different dates from mid-September to mid-October 2007, to ensure a broad coverage of the main migratory period. It is therefore likely that each of the 36 individuals sampled here came from a different breeding site.
Laboratory and data analysis
Feathers were washed, dried, finely cut, and then weighed (0.1 mg) into small silver capsules, and analysed for hydrogen isotope ratios (δ2H) using continuous flow isotope ratio mass spectrometry. Sexing from blood samples was done through the amplification by PCR (polymerase chain reaction) of a fragment of the CHD gene, using the primers P2 and P8 (Griffiths et al. Citation1998).
For analyses of site fidelity and residency we did not consider Robins that were captured only once at constant effort sites. Our aim was to exclude individuals that might be passing through on migration, engaged in post-breeding dispersal, or that were ‘floaters’ only incidentally visiting the CES. This procedure also helps to exclude (temporarily) resident individuals whose home range only marginally overlapped with the CES, hence making capture probability very low. Furthermore, for Robins captured more than once, we only considered birds that stayed at the study area for at least 21 days, to further exclude birds that might be just stopping over at the study site; studies of Robin migration suggest that most stopover durations are shorter than this (e.g. Polak & Szewczyk Citation2007). Finally, captures of young of the year during the period of juvenile dispersal (until the end of September) were not included in these analyses, as such individuals might have chosen to settle permanently in areas outside the CES.
Based on the migration and breeding phenology of Robins in Portugal (Bueno Citation1998, Catry et al. Citation2010, this study) we divided the annual cycle into four periods: Winter, 1 November until 28 February; Spring (breeding season), 1 April until 31 July; the period between 1 and 31 March, and the period between 1 August and 31 October. The latter were defined as transitional and cover the main migration periods as well as much of the post-juvenile dispersal and adult moulting season.
Robins selected according to the above criteria were then classified into six different categories. (1) Sedentary: individuals that were captured both in spring and in winter; (2) Spring–single: individuals captured during one single spring season (and possibly also during a transitional period); (3) Spring–return: individuals captured in more than one spring season; (4) Winter–single: individuals captured during one single winter season (and possibly also during a transitional period); (5) Winter–return: individuals captured in more than one winter season; (6) Uncertain: individuals only captured in transitional periods. We were especially interested in comparing the numbers of individuals classified as Sedentary (1) and as spring visitors (the sum of individuals classified as Spring–single (2) and Spring–return (3)).
RESULTS
Captures were high in October and November, when the main autumnal flux of migrants was felt, and were particularly low in March–May, when few movements occurred and only local nesting birds were present (). The very first juveniles did not appear before May or sometimes June, with large influxes in July (at Madriz, reproduction appeared to be earlier, with many juveniles in May in each year). Juveniles, apparently in post-fledging dispersal, continued to be captured in August and September. Adults were recorded moulting flight feathers from June until mid-September.
Figure 1. Monthly variation in the numbers (mean ± sd) of Robins mist-netted at the two Constant Effort Sites (CES) with larger numbers of captures. Totals include both new captures and recaptures of birds ringed on previous occasions. Data from four years pooled. (a) Margaraça, (b) Madriz.
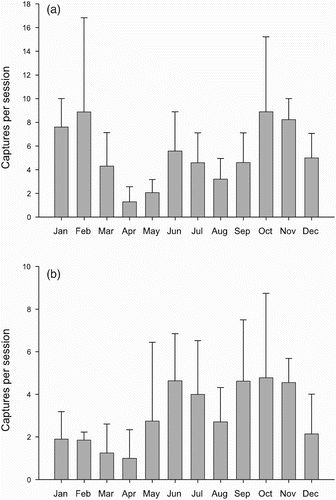
At each of the five study sites, at least some of the Robins were apparently sedentary (). Overall, of the Robins that settled in the study area and were present in the breeding season (adding sedentary birds and spring visitors, n = 66), 52 % were also captured in winter (34 individuals classified as sedentary). Birds staying for winter, at each of the five sites, included not only adults, but also juveniles of the year that were captured in their first summer (excluded from statistics presented in , see Methods).
Table 2. Numbers of individual Robins captured at five Constant Effort Sites (CES) and classified in each phenological category. Figures in brackets represent numbers of sexed individuals (male : female). Male and female numbers do not add to total in categories because many individuals were not sexed.
Comparing Margaraça (500–600 m a.s.l.), where 33 % of the Robins that settled in the breeding season (n = 27) were classified as Sedentary (), with the other four (low-altitude) sites (64 % Sedentary, n = 39), a significant difference in year-round site fidelity was found (Fisher's Exact Test, P = 0.024). Among the low-altitude sites, there was no difference in the proportion of Sedentary birds found at the suburban park in Gaia (78 %, n = 9) versus the other habitats (69 %, n = 26; Fisher's Exact Test, P = 1.0).
Among birds that settled in winter (Sedentary plus Winter visitors), the proportion of birds classified as Winter visitors in Margaraça (81 %, n = 48) differed from the proportion of Winter visitors at the four other sites (61 %, n = 64; Fisher's Exact Test, P = 0.024). Comparing Gaia (46 %, n = 13) with the other low-altitude sites (65 %, n = 51), no significant difference was found when considering the same statistic (Fisher's Exact Test, P = 0.34).
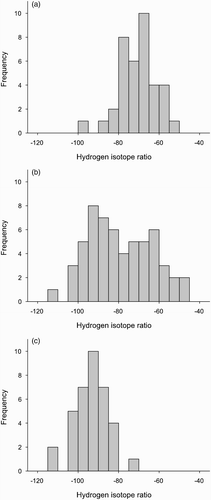
At Margaraça, the distribution of feather δ2H values had a much higher dispersion in winter (δ2H = –79.2 ± 15.5 ‰, n =57) than in spring (δ2H = –70.9 ± 9.1 ‰, n = 37; Homogeneity of Variance Test: Levene Statistic = 14.1, P < 0.001), with a clear increment of the proportion of Robins with more negative δ2H values (). The winter feather δ2H values from Margaraça also had a much higher variance than δ2H values of Robins from Scandinavia (δ2H = –93.5 ± 8.1 ‰, n = 36; Homogeneity of Variance Test: Levene Statistic = 21.5, P < 0.001).
Figure 2. Frequency of isotopic signatures of Robins (feathers) at Margaraça in (a) spring (June) and (b) winter (January) and (c) during autumn migration at Falsterbo, Southern Sweden.
Individuals sampled at Margaraça with more negative δ2H values had longer wing-lengths (Pearson correlation: r = –0.35, n = 57, P = 0.008).
The sex ratio of Robins at Margaraça during winter was fairly even, with 55 % male (n =57 sexed individuals). There were no significant differences in the variance or in the mean of isotopic composition of feathers of males (δ2H = –78.4 ± 13.2 ‰, n = 31) and females (δ2H = –81.1 ± 17.6 ‰, n =25; Levene Statistic = 2.75, P = 0.10; anova F1,55 = 0.41, P = 0.5). Among Robins that became sedentary during the wintering period, only 23 % were recorded in more than one autumn/winter ().
DISCUSSION
This study documents for the first time a geographically widespread sedentary behaviour by a passerine confronted with large numbers of conspecific visitors. Using an approach broadly similar to Domínguez et al. Citation(2007), we have shown that their result of a full population interchange of Robins is not generally applicable in Iberia. Our study sites encompass a range of altitudes, habitats, and local climates that is likely to be representative of much of the large nesting Robin population in Western Iberia (Equipa Atlas Citation2008). The existence of fully sedentary birds in Iberian populations had previously been demonstrated by Pérez-Tris & Tellería Citation(2002b), but from that study it was unclear whether such individuals were the rule or the exception in the sampled populations, or whether the study sites had indeed been flooded by foreign competitors. It had also previously been shown that local Robins partially segregated from migrant visitors in southern Spain during winter (Tellería et al. Citation2001, Tellería & Pérez-Tris Citation2004), with ‘local birds’ being more abundant in those habitats where they also nested. However, those studies did not assess the behaviour of individuals and took a broad population approach instead. The present study suggests that the pattern of general population segregation is caused to a large extent by a majority of local Robins (52%, in our overall sample) that are present in the immediate vicinity of their presumed nesting sites over winter. Nevertheless, there were differences between sites in the proportion of sedentary birds. Those differences may have been related to altitude (and local climate), as previously suggested by Pérez-Tris & Tellería Citation(2002b).
Substantial numbers of Robins that apparently nested near the CES were not captured in winter. This may be explained by three factors. Some birds may have died or permanently emigrated (this obviously does not apply to those captured in subsequent springs). Other birds may have been present, but were not captured because of their familiarity with the net sites and trap-shyness. Finally, some birds may have moved away for the winter. The fact that some birds are indeed likely to have moved away (they were captured multiple times in the breeding season but never in winter) does not necessarily mean that they were forced to do so by increased competition from migrants, as this type of behaviour is present even in populations subjected to no autumnal influx of long-distance migrants. For example, around 50% of the broadly resident Robins in Ireland and England move away from their breeding territories in autumn and early winter, and return in the following nesting season (Burkitt Citation1926, Lack Citation1943, Jackson Citation1958), and the British Isles is a region with very small numbers of migratory Robins (Remisiewicz Citation2002, Wernham et al. Citation2002). If migrants strongly influenced local birds, we should have expected that our study would have experienced very different results from those from the British Isles, which clearly was not the case.
Are Robins from our study sites truly confronted with significant numbers (relative to the local population) of incoming migrants during the non-breeding season? The higher capture rates in the non-breeding season do suggest this, as do the large numbers of recoveries of foreign Robins scattered through Iberia (Bueno 1999). Data from the intensively studied site of Margaraça provide new evidence on this subject. The negative correlation between wing-length and hydrogen isotope ratios in wintering Robins is in line with the assumption that Robins coming from higher latitudes (which have longer wings) also have lower δ2H (see Materials and Methods). This is also confirmed by δ2H values from our Scandinavian sample. The widely contrasting distribution of isotopic signatures of Robins sampled in spring and winter at Margaraça clearly indicates that there is an important influx of Robins from other breeding origins in the cold season. Furthermore, the large variance of winter δ2H values suggests that Robin populations from various sources are simultaneously present at this site. The range of values is consistent with measurements obtained from almost the full distributional range of Robins (), suggesting that the wintering population includes both local birds (also confirmed from CES data) and long-distance migrants from northern Europe.
To get an estimate of a plausible value for the proportion of long-distance migrants at Margaraça in winter, it is possible to use the isotope data presented in . If it is assumed that all of the birds with δ2H < 80 are migrants (a few locals have values < 0.80, but these will be compensated by migrants which have δ2H > 80), then the proportion of migrants would be 53 %. Of course, this is a crude estimate, because Robins from mid-latitudes in Europe will also be present, probably contributing birds to both sides of the cut-off line.
On the other hand, the wintering sex ratio of birds at Margaraça suggests that many local Robins are present. The sex ratio of Robin populations in autumn–winter, at Iberian sites where no Robins nest, is generally heavily female-biased, with 69% females reported in northern Spain (Arizaga et al. Citation2010) and 71–87% in southern Portugal (Catry et al. Citation2004). This results from Robins being differential migrants, with a tendency for females to move farther and males to winter at more northerly latitudes (Adriaensen & Dhondt Citation1990). The fact that, at Margaraça, females represented only 45% of the captures does not support the idea that foreign Robins largely dominate the wintering community, because in this case the sex ratio should be more female-biased. For example, if we assume that sedentary birds were 80% male (a figure obtained from local CES data and that is consistent with results obtained in the British Isles; Burkitt Citation1926, Lack Citation1943, Jackson Citation1958) and that 75% of the wintering migrants were female, an overall winter proportion of 45% females at Margaraça implies that only 46% of the wintering individuals were migrants. Overall, these two fully independent tentative estimates of 53% and 46% (of the winter population as long-distance migrants) give an order of magnitude and suggest that there is not a great imbalance between the numbers of residents and of migrants in the mid-winter Robin population at Margaraça.
The low frequency of individuals classified as Winter–return, as opposed to Winter–single, is well in line with multiple studies documenting that Robins wintering in Iberia have low between-year site fidelity (e.g. Cuadrado Citation1992, Herrera Citation1998, Catry Citation2005). To some extent, Robins spending the winter in Iberia track spatial variations in fruit abundance (Tellería et al. Citation2008) and this may explain the low interannual site fidelity of wintering individuals and possibly also the movements of some individuals of the local population.
In conclusion, this article suggests that incoming migratory populations do not force local Robins to vacate nesting territories and redistribute themselves in the shared wintering range. Whether this happens because local Robins are somewhat dominant over incoming conspecifics or for other reasons, such as different Robin populations having distinct ecological requirements or adaptations, will have to be clarified by further research.
ACKNOWLEDGEMENTS
Many people helped with fieldwork at the constant effort site ringing stations and Hany Alonso provided extra help with sampling at Margaraça. Instituto da Conservação da Natureza e da Biodiversidade gave permission for work to be carried out at several of its nature reserves. Joana Robalo helped with molecular sexing. We are indebted to Falsterbo Ringing Station and its staff for their support during the collection of samples. AC received a grant (SFRH/BD/30724/2006) from Fundação para a Ciência e a Tecnologia (FCT – Portugal); this grant was co-financed by Fundo Social Europeu (FSE) under the Programa Operacional Potencial Humano (POPH) from QREN. JMN was also supported by these institutions through the grant SFRH/BPD/40667/2007. We are grateful for the continued support of FCT through Programa Plurianual (UI&D 331/94). Hydrogen isotope ratio measurements were carried out at the NERC Life Sciences Mass Spectrometry Facility and funded via a NERC LSMSF grant (EK106-01/07). This study complies with the current laws concerning wildlife research in Portugal.
REFERENCES
- Adriaensen , F. and Dhondt , A. A. 1990 . Population dynamics and partial migration of the European Robin Erithacus rubecula in different habitats . J. Anim. Ecol. , 59 : 1077 – 1090 .
- Arizaga , J. , Alonso , D. and Barba , E. 2010 . Patterns of migration and wintering of Robins Erithacus rubecula in northern Iberia . Ring. Migr. , 25 : 7 – 14 .
- Bearhop , S. , Fiedler , W. , Furness , R. W. , Votier , S. C. , Waldron , S. , Newton , J. , Bowen , G. J. , Berthold , P. and Farnsworth , K. 2005 . Assortative mating as a mechanism for rapid evolution of a migratory divide . Science , 310 : 502 – 504 .
- Bell , C. P. 2005 . “ Inter- and intrapopulation migration patterns: ideas, evidence and research priorities ” . In Birds of Two Worlds: The Ecology and Evolution of Migration , Edited by: Greenberg , R. and Marra , P. P. Baltimore : John Hopkins University Press .
- Bueno , J. M. 1998 . Migracion e invernada de pequeños turdinos en la Peninsula Ibérica V. Petirrojo (Erithacus rubecula) . Ardeola , 45 : 193 – 200 .
- Burkitt , J. P. 1926 . A study of the Robin by means of marked birds . Br. Birds. , 20 : 91 – 101 .
- Catry , P. 2005 . Social composition, mass regulation and site fidelity of migrant Robins Erithacus rubecula wintering in a marginal habitat . Airo , 15 : 59 – 64 .
- Catry , P. , Campos , A. , Almada , V. and Cresswell , W. 2004 . Winter segregation of migrant European Robins Erithacus rubecula in relation to sex, age and size . J. Avian Biol. , 35 : 204 – 209 .
- Catry , P. , Costa , H. , Elias , G. and Matias , R. 2010 . Aves de Portugal. Ornitologia do Território Continental , Lisboa : Assírio & Alvim .
- Cox , G. W. 1968 . The role of competition in the evolution of migration . Evolution , 22 : 180 – 192 .
- Cramp , S. 1988 . The Birds of the Western Palearctic. Vol V , Edited by: Cramp , S. Oxford : Oxford University Press .
- Cuadrado , M. 1992 . Year to year recurrence and site-fidelity of Blackcaps Sylvia atricapilla and Robins Erithacus rubecula in a Mediterranean wintering area . Ring. Migr. , 13 : 36 – 42 .
- Domínguez , M. , Barba , E. , Canto , J. L. , López , G. M. and Monrós , J. S. 2007 . Seasonal interchange of the European Robin Erithacus rubecula populations in an evergreen holm oak forest . Acta Ornithol. , 42 : 15 – 21 .
- Equipa Atlas . 2008 . Atlas das Aves Nidificantes em Portugal (1999–2005) ICNB, SPEA, PNM e SRAM. Assírio & Alvim, Lisboa
- Greenberg , R. 1986 . “ Competition in migrant birds in the nonbreeding season ” . In Current Ornithology 3 , Edited by: Johnston , R. F. 281 – 307 . New York and London : Plenum Press .
- Griffiths , R. , Double , M. C. , Orr , K. and Dawson , R. J.G. 1998 . A DNA test to sex most birds . Mol. Ecol. , 7 : 1071 – 1075 .
- Herrera , C. M. 1977 . Ecología alimenticia del petirrojo (Erithacus rubecula) durante su invernada en encinares del sur de España . Doñana, Acta Vert. , 4 : 35 – 59 .
- Herrera , C. M. 1998 . Long-term dynamics of Mediterranean frugivorous birds and fleshy fruits: a 12-year study . Ecol. Monogr. , 68 : 511 – 538 .
- Hobson , K. A. , Bowen , G. , Wassenaar , L. I. , Ferrand , Y. and Lormee , H. 2004 . Using stable hydrogen isotope measurements of feathers to infer geographical origins of migrating European birds . Oecologia , 141 : 477 – 488 .
- Jackson , R. D. 1958 . A study of a population of Robins . Erithacus rubecula. Irish Nat. J. , 12 : 229 – 236 .
- Karlsson , L. , Persson , K. , Pettersson , J. and Walinder , G. 1988 . Fat–weight relationships and migratory strategies in the Robin Erithacus rubecula at two stop-over sites in South Sweden . Ring. Migr. , 9 : 160 – 168 .
- Lack , D. 1943 . The Life of the Robin , London : H.F. & G. Witherby .
- Newton , I. 2008 . The Migration Ecology of Birds , Amsterdam : Academic Press .
- Newton , I. and Dale , L. 1996 . Relationship between migration and latitude among west European birds . J. Anim. Ecol. , 65 : 137 – 146 .
- Pérez-Tris , J. and Tellería , J. L. 2002a . Migratory and sedentary blackcaps in sympatric non-breeding grounds: implications for the evolution of avian migration . J. Anim. Ecol. , 71 : 211 – 224 .
- Pérez-Tris , J. and Tellería , J. L. 2002b . Regional variation in seasonality affects migratory behaviour and life-history traits of two Mediterranean passerines . Acta Oecol. , 23 : 13 – 21 .
- Polak , M. and Szewczyk , P. 2007 . Relation between stopover length and time and body parameters of European Robin Erithacus rubecula (L., 1758) during autumn migration (Central Poland) . Polish J. Ecol. , 55 : 511 – 517 .
- Remisiewicz , M. 2002 . The temporal pattern to Robin Erithacus rubecula migration: evidence from ringing recoveries . Ardea , 90 : 489 – 502 .
- Tellería , J. L. and Pérez-Tris , J. 2004 . Consequences of the settlement of migrant European Robins Erithacus rubecula in wintering habitats occupied by conspecific residents . Ibis , 146 : 258 – 268 .
- Tellería , J. L. , Pérez-Tris , J. , Ramírez , A. , Fernández-Juridic , E. and Carbonell , R. 2001 . Distribution of Robins Erithacus rubecula in wintering grounds: effects of conspecific density, migratory status and age . Ardea , 89 : 363 – 373 .
- Tellería , J. L. , Ramirez , A. and Pérez-Tris , J. 2008 . Fruit tracking between sites and years by birds in Mediterranean wintering grounds . Ecography , 31 : 381 – 388 .
- Wernham , C. V. , Toms , M. P. , Marchant , J. H. , Clark , J. A. , Siriwardena , G. M. and Baillie , S. E. 2002 . The Migration Atlas: Movements of the Birds of Britain and Ireland , Edited by: Wernham , C. V. , Toms , M. P. , Marchant , J. H. , Clark , J. A. , Siriwardena , G. M. and Baillie , S. E. London : T. & A.D. Poyser .