Abstract
Capsule Diet composition differed significantly between winter cereals, winter oil-seed rape, stubble fields and permanent fallows.
Aims To determine the composition of the diet of Grey Partridges in autumn and winter in four agricultural land-cover types, characteristic of lowland areas of Central Europe.
Methods Faecal analysis was used to determine diet. Multivariate analysis of variance (manova), Simpson Index of Diversity (sid) and Detrended Correspondence Analysis (dca) were used to assess variation in the proportions of the six main dietary components (cereal and broad-leaved plant leaves, weed seeds cases, cereal grains, husks of grasses and other plant material).
Results Thirty-seven different kinds of plant food items were found, and the most numerous were cereal leaves (58.2% in total of all items), followed by leaves of broad-leaved plants (21.8%), weed seed cases (13.3%), cereals grains (3.5%), husks of grasses (1.2%) and other plant material (2.0%). Diet composition differed significantly between winter cereals, winter oil-seed rape, stubble fields and permanent fallows. The dca showed that the two first axes explain 38% of the total variance of the diet. The diet diversity was highest in stubble fields and permanent fallows, and the smallest in winter cereals. Dietary diversity was negatively correlated with the overall abundance of leaves, and positively with the abundance of weed seeds, cereal grains and husks of grasses.
Conclusion Cereal leaves might replace other food items, which suggests that food resources are not a critical factor limiting the population of Grey Partridges during winter. A high proportion of weed seeds and cereal grains in the diet of Grey Partridges in stubble fields confirms the importance of these fields as sources of food of high-calorific value. Maintenance of stubble fields and cover crops with natural regeneration of annual weeds should constitute an important element of a strategy for the recovery of Grey Partridge populations in arable landscapes in Europe.
Grey Partridges Perdix perdix are a species with decreasing populations in most European countries (BirdLife International Citation2004, Donald et al. Citation2006, review in Kuijper et al. Citation2009). The main reasons for their decline have been linked to the increasing intensification of agriculture which include the use of herbicides to eliminate weeds from cereals crops and thereby reducing the number of insects available as food for chicks, elimination of boundary nesting cover through field enlargement, and increased predation pressure on the remaining habitat (Potts Citation1980, Citation1986, Panek Citation1991, Citation1997, Citation2005, Baldi & Farago Citation2007, Aebischer & Ewald Citation2010).
Ongoing changes in the availability of different dietary components, weed seeds and cultivated plant products (e.g. spilled cereal grain), which are linked to the disappearance of stubble fields (Wilson et al. Citation1999, Holland et al. Citation2006), imply the need for further studies of the relationships between crop type and the diet of Grey Partridges. The high proportion of leaves (over 90%) in the autumn–winter and early-spring diet of Grey Partridges (Thaisz Citation1912, Middletton & Chitty 1937, reviews in Glutz von Blotzheim Citation1994, Cramp Citation1998), may suggest, however, that the decline of weed seed biomass in crop fields could have a smaller impact on this species (discussed in Potts Citation1970, Steenfeldt et al. Citation1991).
Our aim has been to determine the composition of the diet of Grey Partridges in autumn–winter in four land-cover types which are characteristic of lowland areas of Central Europe, and are the dominant land-use types within arable landscapes of Poland.
Our analysis determined the diet composition of Grey Partridges in two crop types with intensive tillage regimes (winter crops: cereals and oil-seed rape) versus extensive tillage, i.e. stubble fields with natural regeneration of weed flora, and fields excluded from agricultural activity (= abandoned; permanent fallows). We discuss the relationships between the main dietary components and diet diversity. To compare the diet of Grey Partridges in different land-cover types and habitat conditions we compiled available data from Europe and North America and drew up a table with references and detailed data of methods of analysis, land-cover type and food composition.
MATERIAL AND METHODS
Land-cover types and weed flora
Grey Partridge diet was determined using faecal analysis. Faecal samples were collected from agriculture areas of western and eastern parts of Poland in late autumn (10 dates in November) and winter (26 dates, December–February) in the seasons 2004/05–2009/10. Faeces from each site were treated as one sample (n = 36).
Faeces were collected from daily feeding sites of partridges. In some cases, coveys (up to 15 individuals) had been seen before, but in other cases only places with relatively fresh droppings were found. Faeces (up to about 90 pieces) were collected only if the birds were observed at the same place for some time, or if the droppings were distributed over a limited space (e.g. within a radius of several metres) and their number was at least a few dozen (sometimes up to a few hundred), which might indicate that partridges had stayed at the place for a long time during the day, for example for several hours. Crop edges adjacent to other crops (but not to ploughed land) were avoided during the collection of faeces.
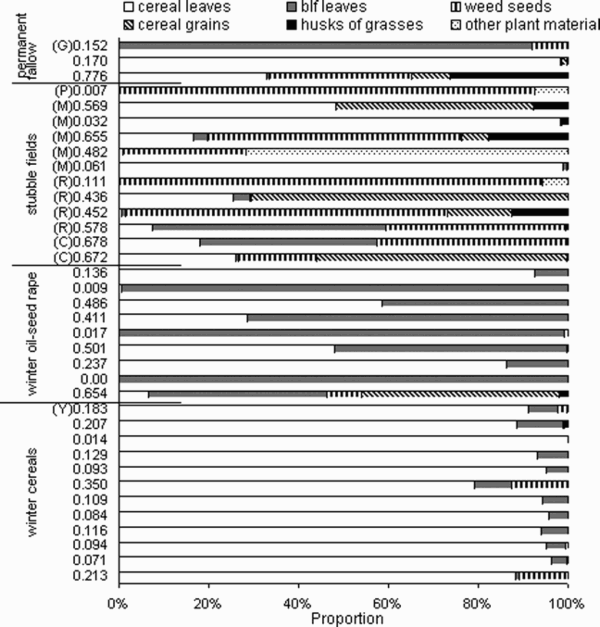
Land-cover types on which the droppings were collected could be divided into four main types (number of samples in brackets): (1) winter cereals, including one sample of droppings from a rye-grass Lolium perenne field (n = 12); (2) winter oil-seed rape (n = 9); (3) stubble fields (n = 12); (4) abandoned agricultural land called permanent fallows (n = 3). Both winter crops were seeded in autumn and were managed in accordance with routine conventional pesticide application and inorganic fertilizers. The category ‘stubble’ included fields temporarily left fallow (for the autumn–winter period) and prepared for spring sowing, i.e. fields without any autumn cultivation or fields shallowly ploughed after harvest. The previous crops on these fields were: maize (n = 5), oil-seed rape (n = 4), cereal (n = 2) and potato (n = 1). The ‘permanent fallows’ category included places of no agricultural activity conducted for at least several years. They were covered by permanent plants and located for instance on a railway bank, on the roadside, and the last one was an abandoned pasture (grassland).
The area of fields where the droppings were gathered ranged between 0.2 and 46 ha (average ± se, 9.0 ± 2.3 ha). The size of fields did not significantly differ (anova Kruskal–Wallis test, H 3,32 = 1.88, P = 0.60) between the four land-cover types; similarly, in a pair-wise post-hoc comparison we did not find any significant difference between the area of fields of the two land-cover types (P ≥ 0.36). We carried out only a basic description of the vegetation on all fields, without detailed quantification of food availability and diversity. Typical weed communities appeared in the particular land-cover type (crops). In winter crops (cereals and oil-seed rape) beyond the main cropped species, the density of weeds was very small, just a few plants per m2; several annual plants were found, mainly in oil-seed rape, with the dominance of Geranium spp., Lamium amplexicaule, Galinsoga spp., Veronica spp. and Stellaria media. The flora of stubble fields was considerably more differentiated; both types of stubbles (uncultivated and ploughed shallowly) often contained self-sown crop species from previous cultivation, i.e. cereals or oil-seed rape. Moreover, the species composition and the density of weeds on stubble fields were usually high. The post-emergence weed communities dominated in stubble fields, with a mosaic of compact vegetation cover formed by annual weed species, mainly Amaranthus retroflexus, Chenopodium album, Setaria viridis and Polygonum aviculare. Permanent fallows were dominated by wasteland communities consisting mainly of perennial species, including alien invasive species Solidago gigantea and S. canadensis accompanied by Tanacetum vulgare and Artemisia vulgaris.
In spite of the fact that our analysis is based on faeces collected during different months and years we believe that the whole period between late autumn and winter (i.e. from November to February) is quite similar with regard to the presence of cultivated plants, weed flora and agricultural practices. In general in Poland, starting with November (postharvest period), crop fields succumb to ‘winter conditions’, i.e. new winter crops already grow in cultivated fields and similar post-emergence weed communities appear in stubble fields. Furthermore, considering a potential daily movement of partridges between different crops and non-cropped habitats, e.g. hedges, wildflower strips and nocturnal sites located at large distances from field edges (Buner et al. Citation2005, Tillmann Citation2008), and on the other hand the broad accessibility to leaves of cultivated plants (being a staple diet), it seems that the assessment of potential food resources or their depletion over autumn–winter period (with the exception of weed seeds or spilled cereal grain whose density may fall throughout this period; Moorcroft et al. Citation2002, Butler et al. Citation2005), might be difficult to link with food composition of Grey Partridges. Hence, considering the above-mentioned similarity in habitat conditions and timing, we decided to conduct diet analysis for pooled faecal samples.
Diet analysis
Droppings were dried and stored at 20°C and 0.5 g of faeces was randomly selected from each sample from detailed analysis of diet composition. The samples were divided into five equal parts (0.1 g each), soaked in distilled water and spread in a thin layer in Petri dishes and then dried up. Samples were examined with a stereoscopic microscope at ×40 magnification and divided into 1 mm2 squares (in total 6358 squares per Petri dish) and the number of identifiable items counted. On this basis the approximate surface area covered by one particular type of food category was assessed, enabling us to calculate the percentage share of each type of food items in each sample. We expressed diet composition as the number and proportion of each of six different food categories. Categories were based on those identified in earlier studies of the diet of Grey Partridges (Green Citation1984; see data compiled in ): (1) cereal leaves; (2) broad-leaved plant leaves (Dicotyledones, mainly leaves of oil-seed rape); (3) fragments of weed seed cases left after the consumption of seeds (hereafter ‘weed seeds’); (4) fragments of grains (mainly cereal grains, hereafter ‘cereal grains’); (5) husks (glumes) of grasses; (6) other plant material (fragments of straws, stems, dry fruits and inflorescences). In the results section, the pooled data on all leaves are also given. The identification of fragments of cultivated plants and weed species was aided by the reference material obtained from the same fields where droppings were collected.
Table 2. Comparison (in chronological order of published study) of autumn–winter diet composition of Grey Partridges Perdix perdix (expressed as percentage area/volume of six main components) in Europe and North America. Abbreviations: cer leav, cereal leaves; blf, broad-leaves; o. plant mat., other plant material; n.d., not determined; Aut., Autumn; Win., Winter; the same letter means pooled diet components: A, cereal leaves and broad-leaved leaves; B, weed seeds and cereal grains.
Statistical analysis
We hypothesised that the diet composition of Grey Partridges would vary with land-cover type, as a result of the different availability of particular food items caused by different agricultural practices. These differences might concern mainly weed seeds (more abundant in stubble fields) and leaves (more abundant in both winter crops). The multivariate analysis of variance (manova) was applied to determine differences in diet among four types of land-cover (winter cereal, winter oil-seed rape, stubble fields and permanent fallows). Post-hoc comparisons (Sheffe test) were applied to assess the differences in abundance of six food categories between land-cover types. In order to allow for differences in the relative proportions of the six main dietary components, multivariate analysis of variance was calculated on the basis of the log-transformed number of items recorded in analysed faecal samples. The same procedure was applied earlier in the analysis of the diet of Reed Buntings Emberiza schoeniclus (Orłowski & Czarnecka Citation2007). In some analyses we combined stubble fields with permanent fallows.
We applied the Simpson Index of Diversity (sid) to assess the diversity of diet at particular sites of faeces gathering. The number of particular food items, presented in , was taken into consideration for the calculation of the sid which was calculated using the Mouillot & Lepretre Citation(1999) equation:
Table 1. The autumn–winter diet composition of Grey Partridges Perdix perdix in Poland based on the analysis of 36 faecal samples (0.5 g) from four land-cover types: winter cereals, winter oil-seed rape, stubble fields after cultivation of cereals (C), oil-seed rape (R), maize (M) and potato (P) and permanent fallow; in brackets the number of samples with a given item indicated; alife cycle of weeds: annual (A), biennial (B), perennial (P).
Detrended Correspondence Analysis (dca) was used to identify the scale of differences at two levels: among four land-cover types and among samples collected at one type of land-cover. The analysis was also used to help determine the main statistically important factors (proportion of different dietary components) responsible for the distribution of points (samples) along Axis 1 and Axis 2 of the diagram. It was used to determine: (1) the level of difference among samples coming from different land-cover types; (2) in which type of land-cover the diet was most diverse; (3) what factors (dietary components) were responsible for these differences. The dca analysis was performed with the mvsp software (Kovach Citation2005).
Finally, Spearman rank correlation coefficient was used to test the relationships between the six food categories and the diversity of the diet (sid), the dca scores and the proportion of particular food categories. The probability of P < 0.05 was assumed as statistically significant.
RESULTS
Thirty-seven different kinds of plant food items were found in the diet of Grey Partridges. The most numerous were the leaves (80.0% in total of all items), among which dominated the leaves of cereals (58.2%), followed by broad-leaved plants (21.8%). In decreasing order, the consecutive groups were weed seed cases, represented by 17 different weed taxa, all belonging to the Dicotyledones (13.3%); followed by cereals grains (3.5%) and husks of grasses (1.2%). Other plant material (fragments of straws, stems, dry fruits and inflorescences) constituted about 2.0%. Among weed seeds, 99.2% of all items were annual weed species (). However, the diet composition of Grey Partridges differed markedly between the analysed land-cover types ().
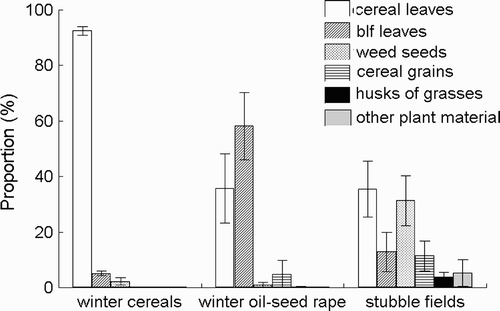
Figure 1. Comparison of the average (± 1 se) proportion of six main components in the diet of Grey Partridges Perdix perdix in winter cereals, winter oil-seed rape and stubble fields combined with permanent fallows. blf, broad-leaved.
Effect of land-cover type on diet composition
The manova for the six main dietary categories indicated significant differences in diet composition among the four main land-cover types (Wilks's Lambda, λ = 0.240, P < 0.001) and for three land-cover types with pooled stubble fields and permanent fallows (Wilks's Lambda, λ = 0.252, P < 0.001). Pair-wise comparison between land-cover types revealed significant differences in the diet composition between winter cereals and winter oil-seed rape (Wilks's Lambda, λ = 0.349, P = 0.011), and between stubble fields combined with permanent fallows and winter cereals (Wilks's Lambda, λ = 0.350, P < 0.001) and winter oil-seed rape fields (Wilks's Lambda, λ = 0.342, P = 0.003).
Post-hoc analyses (Sheffe test) revealed a significantly (P ≤ 0.028) higher abundance (average ± 1 se) of all types of leaves in the diet of birds from both types of winter crops (97.66% ± 1.29% and 93.89% ± 5.98%, for cereals and oil-seed rape, respectively) than in the case of birds from stubble fields (39.55% ± 10.61%); this difference was observed also when we pooled together the data from stubble fields and permanent fallows and compared them with winter cereals (P ≤ 0.016), but not versus winter oil-seed rape (P = 0.067). The abundance of cereal leaves was significantly higher in the diet of birds from winter cereals than in the case of winter oil-seed rape (P = 0.005) and stubble fields combined with permanent fallows (P ≤ 0.002; ). Significantly more broad-leaved plant leaves were present in the diet from winter oil-seed rape than from winter cereals and stubble fields and fallows put together (P ≤ 0.019). The abundance of weed seeds was significantly higher in the case of stubble fields (also combined with permanent fallows) than in both winter crops (P ≤ 0.012). Considering the abundance of weed seeds of the two most abundant weed species, Amaranthus retroflexus and Chenopodium album, the highest proportion of both species was found in stubble fields after rape cultivation ().
The rest of the food categories was not significantly different in any of the analysed land-cover types (), with the exception of cereal grains, whose abundance was close to being significantly higher in the case of combined stubble fields and permanent fallows compared with winter cereals (P = 0.053).
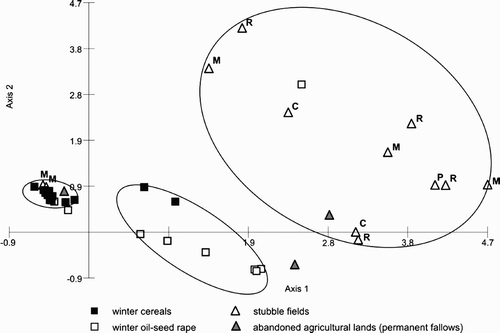
The dca showed that the two first axes explained 38% of the total variance (). A clearly isolated group consisted mainly of winter cereals, where the diet was uniform (see detailed data on the diet composition in particular sites in Appendix 1). The diet variation in the other land-cover types was more differentiated, although two other groups might be distinguished here: one with winter oil-seed rape and cereals, and the other with stubble fields and permanent fallows. The most important factors responsible for the distribution of the sites along axis 1 were the share of cereal leaves and weed seeds in the diet, and in the case of the second axis it was the abundance of broad-leaved plant leaves ().
Figure 2. Results of the dca analysis of Grey Partridges’ Perdix perdix diet composition indicating the breakdown in four land-cover type: winter cereals (▪), winter oil-seed rape (□), stubble fields (▵), permanent fallow (![]()
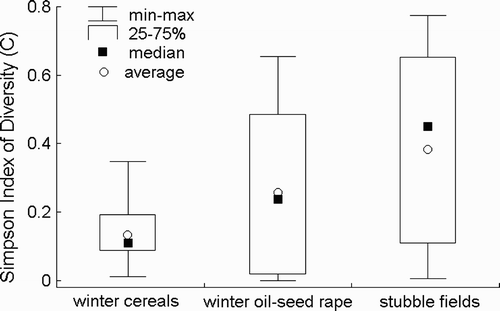
Among the 36 analysed faecal samples the sid was diversified and ranged between 0 and 0.776 (). In particular land-cover types the median and average values of the sid were similar, and they were highest in combined stubble fields and permanent fallows, and the lowest in winter cereals. We have found significant differences in the average value of the sid only between stubble fields combined with permanent fallows and winter cereals (Sheffe test, P = 0.022).
Figure 3. Diet diversity of Grey Partridges Perdix perdix expressed as the Simpson Index of Diversity (sid) in winter cereals, winter oil-seed rape and stubble fields combined with permanent fallows.
Among the six dietary components we found statistically significant positive correlation between weed seeds and other plant material (r s = 0.453, P = 0.005), between cereal grains and husks of grasses (r s = 0.693, P < 0.001), and negative correlation between cereal leaves and weed seeds (r s = –0.498, P = 0.002), between broad-leaved plant leaves and cereal grains (r s = –0.372, P = 0.025), husks of grasses (r s = –0.372, P = 0.025) and other plant material (r s = –0.361, P = 0.031).
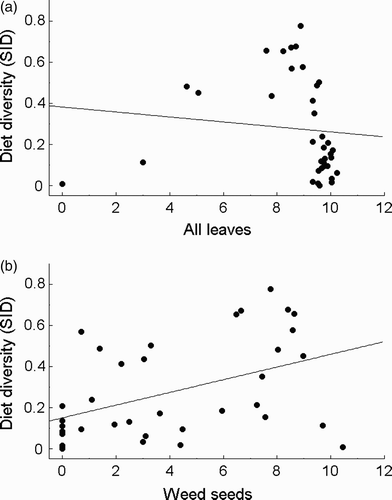
The diet diversity (sid) was negatively correlated with the overall abundance of leaves (; r s = –0.481, P = 0.003), and positively with the abundance of weed seeds (; r s = 0.451, P = 0.006), cereal grains (r s = 0.449, P = 0.006) and husks of grasses (r s = 0.335, P = 0.046).
Figure 4. Relationship between the diet diversity (expressed as the Simpson Index of Diversity, sid) and the log-transformed number of (A) all leaves and (B) weed seeds recorded in the diet of Grey Partridges Perdix perdix based on the analysis of 36 faecal samples from four land-cover types.
DISCUSSION
The results of our study have revealed that the diet composition of Grey Partridges significantly varied among areas of winter cereals, winter oil-seed rape, stubble fields and permanent fallows. The impact of the land-cover type explained 38% of the total variance of the diet, and this high value indicates that habitat features have crucial importance in explaining the diet composition of Grey Partridges in the autumn–winter period. The diet was especially unified in the case of winter cereals, where it was based on cereal leaves, i.e. the food item predominating in this land-cover type. In the other land-cover types (winter oil-seed rape, stubble fields and fallows), the diet was more differentiated and consisted of food items from the other crop types, e.g. we have found a high proportion of cereal leaves in the diet of birds feeding on oil-seed rape fields and a varied proportion of both kinds of leaves in the case of birds from stubble fields (Appendix 1). These discrepancies may be variously explained.
First, we differentiated between only two categories of leaves belonging to Mono- and Dicotyledones (cereals and broad-leaved plants), which in practice means that some samples may have contained various non-identified species from the two plant groups, including Monocotyledons other than cereals, e.g. wild grasses. Secondly, the reason for these discrepancies could also be connected with the movements of birds and the use by them of different kinds of agricultural land-cover types. In general, the winter oil-seed rape is a land-cover type preferred by Grey Partridges in winter (Buner et al. Citation2005), although they tend to choose only the sites with low vegetation, up to 12 cm in height; sites with taller plants are avoided (Tillman 2008). In our study, vegetation reached about 30–40 cm on some rape fields, and that might suggest a limited quality of these sites in the context of the anti-predator strategy of Grey Partridges (Watson et al. Citation2007, Tillman 2008); it might have prevented the birds from spending a long time in oil-seed rape and persuaded them to often move to neighbouring crops with lower vegetation, e.g. winter cereals. Thirdly, another important factor are the food resources in a given crop type (also in the context of snow cover and/or depletion of individual items), which is especially apparent in the case of stubble fields and cover crops with rich weed vegetation. The highest proportion of weed seeds was found in the diet of the birds from these land-cover types. Finally, general differences in diet composition apparently may reflect the availability of individual food items in particular agricultural land-cover types.
Food quality of Grey Partridges: seeds versus leaves. Potential implications for species management and conservation
An important question concerning strategy for the conservation of Grey Partridges in arable landscapes is the nutritive value of particular food items. In the context of our study this will require finding out whether cereal leaves alone provide sufficient nutrition. Most of the studies published to date have indicated the vital importance of winter weed-rich land-cover types, mainly stubble fields, for Grey Partridges’ abundance in the non-breeding period and their later occupancy of territories in spring (e.g. Henderson et al. Citation2009, Ewald et al. Citation2010). The previous experimental study of the preferences of Grey Partridges towards individual food items indicated a higher rate of acceptability of weed seeds, for example Galeopsis spp., than cereal leaves (Pulliainen Citation1965). However, Potts (Citation1970, Citation1986) did underline that green parts of cereals are a good source of high-quality protein and that a diet consisting entirely of green barley is probably not deficient; he also did not find a correlation between the type of food and the condition of adult partridges. Moreover, leaves of other plants, mainly clover and alfalfa, had constituted the staple diet of Grey Partridges (up to 98.7%) before the introduction of winter cereals (Middletton & Chitty 1937, ). As our study revealed, the high proportion of leaves resulted in a decrease in the diet diversity, and conversely, weed seeds increased it. In the light of this, it may be concluded that cereal leaves might replace other food items, especially weed seeds. This suggests that food resources are not a critical factor, in terms of absolute availability (quantity), in limiting the population of Grey Partridges during winter (see Stephens et al. Citation2003).
It should also be emphasized that the presence of uncultivated land and field edges, which result from set-aside or the management of tilled land as stubble fields, has a positive effect on Grey Partridges and other farmland wildlife associated with arable landscapes; similarly. However, these features also have positive effects on the presence of predators. In this context, both field edges, stubble fields or places of artificial feeding may act as ecological traps (Bro et al. Citation2004, Reid et al. Citation2007, Rantanen et al. Citation2010). Hence, winter crops could be considered as an important land-cover type for Grey Partridges in the autumn–winter period which assure both ‘safe’ roosting and suitable feeding sites. In the light of this, we conclude that the lower nutrient value of cereal leaves (discussed below) in comparison to seeds is compensated by their being a widespread and virtually undepletable resource.
However, according to Potts Citation(1986), food gathering would be far more efficient when partridges have access to grain. He observed that, ‘a partridge existing at a mean temperature of –15°C, has the alternative, so to speak, of 1000 pecks per day feeding on wheat grains or 24 500 pecks per day feeding on wheat leaves’. The calorific value of weed seeds and grain is considerably higher than cereal leaves, which in practice means that shorter time is needed to sustain energy demands feeding on seeds than on leaves (Potts Citation1986). Consequently, the major determinant of winter survival of Grey Partridges lies in the reduction in time allocation to predator vigilance because of the need to increase the amount of time available for foraging (Potts Citation1986, Putaala et al. Citation1995, Watson et al. Citation2007). Our results indicate that a high proportion of weed seeds and cereal grain in the diet of Grey Partridges in stubble fields confirms the importance of these land-cover types as sources of high-calorific value food.
Importance of stubble fields with post-emergence weed communities as feeding grounds for Grey Partridges
Cereal grains and weed seeds are indicated as important food items for Grey Partridges in autumn and winter (Potts Citation1986). Considerable amounts of grain were typically found in the winter diet of Grey Partridges in North America, for example 72–80% of food volume in Alberta (Westerskov Citation1966), 65% in Montana (Weigand Citation1980) and 72% in North Dakota (Kobriger Citation1980, see ). In our study, the general proportion of cereal grain in the diet of partridges was small (3.4%), although in some samples from stubble fields it sometimes reached about 70% (see Appendix 1). Species composition and proportion of weed seeds in Grey Partridges’ diet may considerably differ in space and time. For instance, Potts (Citation1970, Citation1980) indicates that Polygonum spp. seeds were most commonly eaten by these birds in England, but their amount in partridge crops in autumn decreased from 31% of total food in 1933–1937 to 4% in 1968–1977. Consequently, the proportion of Polygonum spp. seeds in the total dry weight of dicotyledonous seeds found in partridges’ food decreased from 88% in 1932–1936 to 44% in 1968 (Potts Citation1970). According to Pulliainen Citation(1984), Galeopsis spp. was the most important seed species for partridges in Finland, but its proportion in dry weight of autumn food declined between 1968–1970 and 1979–1981 from nearly 40% to 18%. The results of our study show that in some stubble fields the proportion of weed seeds in partridges’ diet reached up to 92% (Appendix 1). However, among the taxa mentioned above, only Polygonum spp. seeds occurred in low quantity (). On the other hand, it seems that nowadays the importance of two other dicotyledonous weed species has clearly increased, showing the apparent positive abundance trend on crop fields of Central Europe and providing alternative food resources for Grey Partridges, namely Amaranthus retroflexus and Chenopodium album (Baessler & Klotz Citation2006, Rola et al. Citation2002); they constitute 9.1% and 3.2% of all food items found in our study (). In western Europe only Chenopodium album has been mentioned previously as a major component of the diet of Grey Partridges during winter (Potts Citation1970, Wilson et al. Citation1999, Holland et al. Citation2006, ). Amaranthus retroflexus and Chenopodium album form aerial seed banks on crop fields of temperate regions of Northern Hemisphere (Costea et al. Citation2004). Most probably, the high importance of these species in the diet of Grey Partridges resulted from both the increasing density of seeds on crop fields (Rola et al. Citation2002) and the relatively high protein and nitrogen level in comparison to other weed seeds and plant tissues available on arable fields (Crawley Citation1997, Costea et al. Citation2004). Amaranthus retroflexus was an important element of the diet of Grey Partridges already in 1950–1960 in Hungary and Poland, the proportion of seeds of this weed in the diet reached 42.0% and 16.7% in December, respectively (Vertse et al. Citation1955, Oko Citation1963, compiled in ). Furthermore, it should be pointed out that 99.2% of all recorded items classified as weed seeds belonged to annual weeds associated with post-emergence plant communities growing on crop fields after harvesting time. These results have clear implications for the management and conservation of Grey Partridges on tilled land and indicate the need for the maintenance of ‘short-term’, autumn–winter stubbles with natural vegetation of annual weeds. In this context, the reduced tillage systems, that are used by an increasing number of farmers in western Europe and which rely on the use of herbicides to control weeds that emerge from the post-harvest stubble (Cunningham et al. Citation2004), are less beneficial to Grey Partridges and several other species of seed-eating bird, in comparison to the traditional maintance of stubble fields. Crop fields which are uncultivated for a longer period of time (2–3 years), overgrown by biennials and perennials due to vegetation succession, have a considerably smaller value for Grey Partridges (reviewed by Kuijper et al. Citation2009).
Methodological and practical conclusions
Eurasian Sky Larks Alauda arvensis are a species with a similar range of plant diets to that described for Grey Partridges (Green Citation1978, Citation1984, Donald et al. Citation2001), and individuals of both species seem to make use of a wide range of food resources from whatever is available in the landscape (Janda et al. Citation1952, Oko Citation1963, Potts Citation1970). This suggests that providing for the food requirements of these dietary generalists needs a habitat rather than an approach based upon the provision of certain food types (Steenfeldt et al. Citation1991). It might be concluded that general descriptions of the diet composition of Grey Partridges (prevailing in published works based on the crop content analysis of shot birds, ), most probably in a large extent overlap with a local land-use and composition of crop types (changing also in time), rather than show the changes in the diet composition affected by intensification of agriculture (discussed also in Wilson et al. Citation1999). Moreover, it seems that the generalisation of the diet composition, without considering an effect of land-cover type, may lead to false conclusions about the causes of dietary changes of Grey Partridges in relation to agricultural intensification. Hence, it should be clearly emphasized that diet composition should be determined on a large number of samples. It must be high enough to take into account high spatial and temporal variability, and it should be correlated with the local food availability and composition of different food taxa in the same area and time as diet composition studies are carried out (Wilson et al. Citation1999).
Summing up our results and reviewing the management measures for the overall conservation of this species (including food requirements of Grey Partridges chicks and adult birds during spring and summer; Panek Citation1997, Southwood & Cross Citation2002), we conclude that over winter and early spring cover (stubbles, cover crops) provide both weed seeds and anti-predator vegetation structure. Such land management could potentially increase the number of pairs in the breeding population. However, creating areas of both weed-rich land-cover types, i.e. stubble fields and cover crops with natural regeneration of annual weeds (as discussed above) alongside areas of winter cereal crops, will provide the best landscape for a recovery programme of Grey Partridges in arable regions of Europe (Kuijper et al. Citation2009, Ewald et al. Citation2010).
ACKNOWLEDGEMENTS
We are grateful to the anonymous reviewer for the very constructive remarks and comments which greatly improved the value of this article, and to Marek Martini and Artur Goławski for the collection of some droppings of Grey Partridges.
REFERENCES
- Aebischer , N. J. and Ewald , J. A. 2010 . Grey Partridge Perdix perdix in the UK: recovery status, set-aside and shooting . Ibis , 152 : 530 – 542 .
- Baessler , C. and Klotz , S. 2006 . Effects of changes in agricultural land-use on landscape structure and arable weed vegetation over the last 50 years . Agric. Ecos. Environ. , 115 : 43 – 50 .
- Baldi , A. and Farago , S. 2007 . Long-term changes of farmland game populations in a post-socialist country (Hungary) . Agric. Ecos. Environ. , 118 : 307 – 311 .
- BirdLife International . 2004 . Birds in Europe: Population Estimates, Trends and Conservation Status , Cambridge , , UK : BirdLife International .
- Bro , E. , Mayot , P. , Corda , E. and Reitz , F. 2004 . Impact of habitat management on grey partridge populations: assessing wildlife cover using a multisite BACI experiment . J. Appl. Ecol. , 41 : 846 – 857 .
- Brüll , H. 1960 . Der derzeitige Stand subtiler Äsungsstudies des Flugwildes . Inst. Biol. Field Res. Meded. , 50 : 18 – 28 . (in German)
- Buner , F. , Jenny , M. , Zbinden , N. and Naef-Daenzer , B. 2005 . Ecologically enhanced areas – a key habitat structure for re-introduced grey partridge Perdix perdix. Biol. Conserv . 124 : 373 – 381 .
- Butler , S. , Bradbury , R. and Whittingham , M. 2005 . Stubble height affects the use of stubble fields by farmland birds . J. Appl. Ecol. , 42 : 469 – 476 .
- Cambell , J. W. 1936–37 . On the food of some British birds . Br. Birds , 30 : 209 – 218 .
- Costea , M. , Weaver , S. and Tardif , F. J. 2004 . The biology of Canadian weeds. 130 . Amaranthus retroflexus L., A. powelli S. Watson and A. hybridus L. Can. J. Plant Sci. , 84 : 631 – 668 .
- Cramp , S. 1998 . The Complete Birds of the Western Palaearctic on CD-ROM , Edited by: Cramp , S. Oxford , , UK : Oxford University Press .
- Crawley , M. J. 1997 . “ Plant–herbivore dynamics ” . In Plant Ecology Edited by: Crawley , M. J. 401 – 474 . Blackwell Science, UK
- Cunningham , H. M. , Chaney , K. , Bradbury , R. B. and Wilcox , A. 2004 . Non-inversion tillage and farmland birds: a review with special reference to the UK and Europe . Ibis , 146 : 192 – 202 .
- Donald , P. F. , Buckinngham , D. L. , Moorcroft , D. , Muirhead , L. B. , Evans , A. D. and Kirby , W. B. 2001 . Habitat use and diet of skylarks Alauda arvensis wintering on lowland farmland in southern Britain . J. Appl. Ecol. , 38 : 536 – 547 .
- Donald , P. F. , Sanderson , F. J. , Burfield , I. J. and Van Bommel , F. P.J. 2006 . Further evidence of continent-wide impacts of agricultural intensification on European farmland birds, 1990–2000 . Agric. Ecos. Environ. , 116 : 189 – 196 .
- Ewald , J. A. , Aebischer , N. J. , Richardson , S. M. , Grice , P. V. and Cooke , A. I. 2010 . The effect of agri-environment schemes on grey partridge at the farm level in England . Agric. Ecos. Environ. , 138 : 55 – 63 .
- Glutz von Blotzheim, U. N. (ed.) 1994. Handbuch der Vögel Mitteleuropas. Band 5. Galliformes and Gruiformes, p. 272–281. Vogelzug-Verlag im Humanitas Buchversand. Aula-Verlag, Wiesbaden.
- Green , R. E. 1978 . Factors affecting the diet of farmland skylark Alauda arvensis . J. Anim. Ecol. , 47 : 913 – 928 .
- Green , R. E. 1984 . The feeding ecology and survival of partridge chicks (Alectoris rufa and Perdix perdix) on arable farmland in East Anglia . J. Appl. Ecol. , 21 : 817 – 830 .
- Hammer , M. , Køie , M. and Spärk , R. 1958 . Investigations on the food of Partridges, Pheasants and Black Grouse in Denmark . Dan. Rev. Game Biol. , 3 : 185 – 208 .
- Henderson , I. G. , Ravenscroft , N. , Smith , G. and Holloway , S. 2009 . Effects of crop diversification and low pesticide inputs on bird populations on arable land . Agric. Ecos. Environ. , 129 : 149 – 156 .
- Holland , J. M. , Hutchinson , M. A.S. , Smith , B. and Aebischer , N. J. 2006 . A review of invertebrates and seed-bearing plants as food for farmland birds in Europe . Ann. Appl. Biol. , 148 : 49 – 71 .
- Hupp , J. W. , Ratti , J. T. and Smith , L. M. 1988 . Gray partridge foraging ecology in eastern South Dakota . Great Basin Naturalist , 48 : 202 – 205 .
- Huss , D. A. 1983 . Zur Ernährung des Rebhuns (Perdix perdix) in einen Nordburgenländischen Ackerbaugebiet . Egretta , 26 : 1 – 14 . (in German)
- Janda , J. 1956 . Potrava dospelych koroptvi (Perdix perdix L.) v ČSR . Acta Soc. Zool. Bohemslov. , 20 : 147 – 161 . (in Czech)
- Janda , J. , Novakova , E. and Pujman , V. 1952 . Vysledky vyzkumne akce o koroptvi . Straz myslivosti , 30 : 111 – 124 . (in Czech)
- Kelso , L. 1932 . A note on the Food of the Hungarian Partridge . Auk , 49 : 204 – 207 .
- Kobriger , G. D. 1980 . “ Foods, food availability, nutrition, and body weights of Gray Partridge in North Dakota ” . In Proceedings of Perdix II Gray Partridge Workshop , Edited by: Peterson , S. R. and Nelson , L. 70 – 86 . Moscow , , Idaho : University of Idaho .
- Kovach, W.L. 2005. MVSP – A MultiVariate Statistical Package for Windows, ver. 3.1. Kovach Computing Services, Pentraeth, Wales, UK
- Kuijper , D. P.J. , Oosterveld , E. and Wymenga , E. 2009 . Decline and potential recovery of the European grey partridge (Perdix perdix) population – a review . Eur. J. Wild. Res. , 55 : 455 – 463 .
- Middleton , A. D. and Chitty , H. 1937 . The food of adult partridges, Perdix perdix and Alectoris rufa, in Great Britain . J. Anim. Ecol. , 6 : 322 – 336 .
- Moorcroft , D. , Whittingham , M. J. , Bradbury , R. B. and Wilson , J. D. 2002 . The selection of stubble fields by wintering granivorous birds reflects vegetation cover and food abundance . J. Appl. Ecol. , 39 : 535 – 547 .
- Mouillot , D. and Lepretre , A. 1999 . A comparison of species diversity estimators . Res. Popul. Ecol. , 41 : 203 – 215 .
- Novoa , C. , Gonzalez , R. G. and Aldezabal , A. 1999 . Autumn diet of the Grey Partridge (Perdix perdix hispaniensis) in eastern Pyrenees . Rev. Ecol. Ter. Vie , 54 : 149 – 166 . (in French)
- Oko, Z. 1963. Studies on the food of the adult partridges (Perdix perdix L.) in the Poznań Province in a year cycle 1960–1961. Prace Komisji Nauk Rolniczych i Komisji Nauk Leśnych 14(1): 39–96 (in Polish).
- Orłowski , G. and Czarnecka , J. 2007 . Winter diet of reed bunting Emberiza schoeniclus in fallow and stubble fields . Agric. Ecos. Environ. , 118 : 244 – 248 .
- Panek , M. 1991 . Veränderungen in der Populationsdynamic des Rebhuhns (Perdix perdix) in der Gegend von Czempiń, Westpolen, in den Jahren 1968 bis 1988 . Z. Jagdwiss. , 37 : 116 – 124 . (in German)
- Panek , M. 1997 . The effect of agricultural landscape structure on food resources and survival of grey partridge Perdix perdix chicks in Poland . J. Appl. Ecol. , 34 : 787 – 792 .
- Panek , M. 2005 . Demography of grey partridges Perdix perdix in Poland in the years 1991–2004: reasons of population decline . Eur. J. Wildl. Res. , 51 : 14 – 18 .
- Potts , G. R. 1970 . Studies on the changing role of weeds of the genus Polygonum in the diet of the partridge Perdix perdix L . J. Appl. Ecol. , 7 : 567 – 576 .
- Potts , G. R. 1980 . The effect of modern agriculture, nest predation and game management on the population ecology of partridges (Perdix perdix and Alectoris rufa) . Adv. Ecol. Res. , 11 : 2 – 79 .
- Potts, G.R. 1986. The Partridge: Pesticides, Predation and Conservation. Collins, London.
- Pulliainen, E. 1965. Studies on the weight, food and feeding behaviour of the partridge (Perdix perdix L.) in Finland. Ann. Acad. Sci. Fenn. A IV. 93: 1–76
- Pulliainen , E. 1984 . Changes in the composition of the autumn food of Perdix perdix in west Finland over 20 years . J. Appl. Ecol. , 21 : 133 – 139 .
- Putaala , A. , Hohtola , E. and Hissa , R. 1995 . The effect of group size on metabolism in huddling grey partridge . Comp. Biochem. Physiol. , 111B : 243 – 247 .
- Rantanen , E. M. , Buner , F. , Riordan , P. , Sotherton , N. and Macdonald , D. W. 2010 . Habitat preferences and survival in wildlife reintroductions: an ecological trap in reintroduced grey partridge . J. Appl. Ecol. , 47 : 1357 – 1364 .
- Reid , N. , Mcdonald , R. A. and Montgomery , W. I. 2007 . Mammals and agri-environmental schemes: hare haven or pest paradise? . J. Appl. Ecol. , 44 : 1200 – 1208 .
- Rola, H., Rola, J., Kucharski, M., Stupnicka-Rodzynkiewicz, E., Kapeluszny, J. & Skrzyczyńska, J. (eds.) 2002. Share of Amaranthus retroflexus, Chenopodium album and Echinochloa crus-galli in segetal communities under consideration of biotypes resistant to triazine herbicides. Institute of Soil Science and Plant Cultivation, Puławy, Poland. Pamiętnik Puławski 129. 273 pp (in Polish).
- Southwood , T. R.E. and Cross , D. J. 2002 . Food requirements of grey partridge Perdix perdix chicks . Wild. Biol. , 8 : 175 – 183 .
- Steenfeldt , S. , Rasmussen , P. N. and Jensen , T. S. 1991 . Food selection in a population of Partridge Perdix perdix in Danish arable farmland . Dansk. Orn. Tidsskr. , 85 : 67 – 76 .
- Stephens , P. A. , Freckleton , R. P. , Watkinson , A. R. and Sutherland , W. J. 2003 . Predicting the response of farmland bird populations to changing food supplies . J. Appl. Ecol. , 40 : 970 – 983 .
- Thaisz, L. 1912. Über den wirtschaftlichen Nutzen des Rebhuhns – Perdix perdix (L.). Aquila 19: 166–201
- Tillmann , J. E. 2008 . Fear of the dark: night-time roosting and anti-predation behaviour in the grey partridge (Perdix perdix L.) . Behaviour , 146 : 999 – 1023 .
- Vertse, A., Zsak, K. & Kaszab, Z. 1952–1955. Food and agricultural importance of the Partridge (Perdix perdix L.) in Hungary. Aquila 59-62: 13–68 (in Hungarian).
- Watson , M. , Aebischer , N. J. and Cresswell , W. 2007 . Vigilance and fitness in grey partridges Perdix perdix: the effects of group size and foraging-vigilance trade-offs on predation mortality . J. Anim. Ecol. , 76 : 211 – 221 .
- Weigand, J.P. 1980. Ecology of the Hungarian Partridge in North-central Montana. Wildlife Monographs 74.
- Westerskov , K. 1966 . Winter food and feeding habits of the partridge (Perdix perdix) in the Canadian prairie . Can. J. Zool. , 44 : 303 – 322 .
- Wilson , J. D. , Morris , A. J. , Arroyo , B. , Clark , S. and Bradbury , R. 1999 . A review of the abundance and diversity of invertebrate and plant foods of granivorous birds in northern Europe in relation to agricultural change . Agric. Ecos. Environ. , 75 : 13 – 30 .
APPENDIX 1.
Diet of Grey Partridges Perdix perdix in 36 sites including 4 land-cover types, expressed as the proportion of six main dietary components counted in 0.5 g sample of faeces. blf, broad-leaved; numbers near bars denote the Simpson Index of Diversity (sid). Crop type code: abandoned grassland (G), stubble fields after cultivation of rape (R), cereals (C), maize (M), potatoes (P), rye-grass Lolium perenne (Y).