Abstract
Capsule The information on the spatial distribution of cattle farming stored in public bovine computerized databases can predict the distribution and abundance of breeding Barn Swallow Hirundo rustica L. across Europe.
Aims To develop and validate models of habitat factors which account for the distribution of breeding Barn Swallow colonies.
Results The models were based on data on the distribution of cattle farming provided by the public Bovine Computerized Database of the Regione Lombardia (northern Italy). Cattle distribution was a strong predictor of presence and size of Barn Swallow colonies as well as of the number of swallow colonies in a municipality. The models were robust and passed a cross-validation procedure and were used to estimate the spatial distribution of about 116,000 breeding pairs in a wide area (8695 km2) of the low Po plain of northern Italy in 2001.
Conclusions Bovine computerized databases are mandatory in all European Union (EU) countries according to the EU Regulation (CE) 1760/2000. They may serve as a basis for wide scale modelling of the distribution and abundance of breeding Barn Swallow in Europe.
Farmland birds are suffering marked declines in their populations, partly owing to changes in agricultural practices throughout Europe (Tucker & Heath Citation1994, Donald et al. Citation2001, Butler et al. Citation2007). Biological conservation plans aiming at counteracting such negative trends need an appropriate knowledge of the spatial distribution of species at a large geographical scale. Ideally, this requires both intensive and large-scale field studies, which are impractical in most cases. Animal–habitat models, i.e. models relating organisms to their environments (Fielding & Haworth Citation1995, Vaugan & Ormerod Citation2003), provide tools for partly overcoming this constraint as they allow predicting species distribution in unstudied areas, provided that they are properly validated and robust, i.e. applicable also to areas with partly different ecological conditions (Fielding & Haworth Citation1995). Even when these conditions are met, the ability of animal–habitat models to predict the distribution of organisms depends on the spatial scale at which they were built. Indeed, it has been shown that it is incorrect to assume a priori that an ecological process observed at a given spatial scale holds at larger spatial scales, as the link between ecological conditions and species response can markedly change and even be reversed according to the scale at which the process is being observed (Wiens et al. Citation1986, Denny et al. Citation2004, Cadotte & Fukami Citation2005). Models can, therefore, be properly applied only on data collected at the same spatial scale of those used to build them. On the other hand, a detailed knowledge of the spatial scales at which ecological relations hold would be of prime importance for planning cost-effective conservation strategies for declining species, as the conservation value of geographical areas can be ranked according to the expected number of individuals that they can host (Vaugan & Ormerod Citation2003).
Barn Swallows Hirundo rustica are a species of conservation interest in European farmland ecosystems. They have suffered a sharp decline in most parts of their breeding range (Tucker & Heath Citation1994). In the area where the present study was carried out (the Po plain of northern Italy) population decline has been –4.3% breeding pairs per year between 1992 and 2007 (Bani et al. Citation2009). Barn Swallows breed semi-colonially in rural buildings and their distribution is strongly linked to the presence of livestock, particularly cattle farming (Møller Citation2001, Ambrosini, Bolzern, Canova, Arieni et al. Citation2002, Turner Citation2006). Several studies have indeed demonstrated that swallow colonies are larger in farms where cattle are reared and have declined markedly after cessation of cattle farming (Møller Citation2001, Ambrosini, Bolzern, Canova & Saino Citation2002). This evidence obviously suggests that the spatial distribution of cattle farming largely influences that of Barn Swallows by affecting both colony distribution and number of breeding pairs at each colony. However, it is still unknown whether this association holds at spatial scales larger than individual farms.
In the European Union (EU), information on the spatial distribution of cattle farming is stored in public computerized databases held by national or regional administrations. According to the EU Regulation (CE) 1760/2000, which establishes a system for the identification and registration of bovine animals and the labelling of beef and beef products, computerized databases are mandatory in all member states of the EU. Specifically, the computerized databases must hold information on the identification number of all animals of the bovine species present on a holding (that corresponds to any place where bovines are held, i.e. ‘farms’ for the purposes of the present research) (Directive 64/432/EEC, amended by Council Directive 97/12/EC of 17 March 1997).
In this study we used data on cattle farming distribution provided by the Anagrafe Bovina della Regione Lombardia (Bovine Computerized Database of Lombardy; hereafter BCDL) to develop and validate animal–habitat models of the distribution and abundance of breeding Barn Swallows at the spatial scale of a farm and at that of a municipality. The model at the municipality spatial scale was then used to produce maps of the predicted distribution of Barn Swallow colonies in a wide (8695 km2) agricultural area in Lombardy (northern Italy). Hence, this paper refers to a geographical scale one order of magnitude wider than any previous studies of the Barn Swallow distribution (Ambrosini, Bolzern, Canova, Arieni et al. Citation2002, Ambrosini & Saino Citation2010).
METHODS
Study area and data
The Parco Regionale Adda Sud (hereafter referred to as the Park) is an intensively cultivated area (about 240 km2) approximately located in the middle of the low Po plain of Lombardy (,b; see also later). For the purposes of the present research we defined a ‘farm’ as a group of rural buildings separated by at least 100 m from other groups of buildings (Ambrosini, Bolzern, Canova, Arieni et al. Citation2002). In 160 randomly chosen farms in the Park (corresponding to 23.5% of those present in the Park) (, all nests were inspected at least once every 14 days between April and July 2001, and the number of breeding pairs at each farm was estimated as the maximum number of nests simultaneously active (i.e. with eggs or nestlings); see Ambrosini, Bolzern, Canova, Arieni et al. (Citation2002) for a description of the study area and census methods. Data from the census were used to build models of the distribution of breeding Barn Swallows both at farm and municipality spatial scales.
Figure 1. The study area: (a) Lombardy (black) in Italy (grey) and Europe; (b) Lombardy with the 489 municipalities in the low Po plain (grey), the Parco Regionale Adda Sud (white border) and the 168 point counts (dots) used for map validation; (c) the Park with the 160 censused farms (open circles, farms with no breeding Barn Swallows; grey circles, 1–9 breeding pairs; full circles, 10–76 breeding pairs); (d) predicted colony presence and size at all the 682 farms in the municipalities of the Park based on both logistic and linear models (open circles, farms where the logistic model predicted absence of swallows and the quasi-Poisson model predicted presence of 2 breeding pairs[se =± 0.66, 95% CI = 1–4]; grey circles, farms with 7 predicted breeding pairs [se =± 3.16, 95% CI = 3–17]; full circles, farms with 14 predicted breeding pairs [se =± 1.41, 95% CI = 12–17]).
![Figure 1. The study area: (a) Lombardy (black) in Italy (grey) and Europe; (b) Lombardy with the 489 municipalities in the low Po plain (grey), the Parco Regionale Adda Sud (white border) and the 168 point counts (dots) used for map validation; (c) the Park with the 160 censused farms (open circles, farms with no breeding Barn Swallows; grey circles, 1–9 breeding pairs; full circles, 10–76 breeding pairs); (d) predicted colony presence and size at all the 682 farms in the municipalities of the Park based on both logistic and linear models (open circles, farms where the logistic model predicted absence of swallows and the quasi-Poisson model predicted presence of 2 breeding pairs[se =± 0.66, 95% CI = 1–4]; grey circles, farms with 7 predicted breeding pairs [se =± 3.16, 95% CI = 3–17]; full circles, farms with 14 predicted breeding pairs [se =± 1.41, 95% CI = 12–17]).](/cms/asset/d5b35fd5-e18e-4773-be89-14e7ecef974a/tbis_a_609883_o_f0001g.gif)
The Park intersects the territory of 35 municipalities, in 33 of which 1–17 farms were censused. Models at municipality spatial scale were applied to all the municipalities in the low Po plain of Lombardy (i.e. the part of the Po plain south of a line of fault springs also called ‘fontanili’; Rossetti et al. Citation2005), an intensively cultivated area in northern Italy. Municipalities south of Stradella and Voghera (Pavia province) were excluded as they are in the Apennines mountain area. This selection resulted in the inclusion of 489 municipalities covering 8695 km2.
Point count data were collected during the MITO2000 monitoring programme of birds breeding in Italy (Fornasari et al. Citation2002, www.mito2000.it) with the unlimited distance 10-minute point count technique (Fornasari et al. Citation1998). Between 10 May and 20 June 2001 168 point counts from 70 municipalities in the low Po plain in Lombardy ( were performed from sunrise to 11:00 hours in good weather (sunny to cloudy, with no rain or strong wind). Data from point counts were used to validate the prediction from the model at municipality spatial scale.
Data concerning cattle farming were obtained from the BCDL database that holds the address of each farm with cattle farming in Lombardy, and that of farms where cattle farming ceased in the last four years. We preliminarily geo-referenced from detailed maps (scale 1:10,000) all 682 farms in all the municipalities of the Park. This allowed us to identify the farms of the Park which have been without cattle farming for more than four years, as they corresponded to the geo-referenced farms not included in the BCDL. At the farm spatial scale, BCDL data were summarized as a three level factor: ‘cattle farming’ indicating whether cattle was reared in a farm in 2001, if cattle farming had ceased in the previous four years, or if it had not been practiced for more than four years (or it had never been practiced). At the municipality spatial scale these data were summarized as continuous variables accounting for the log-transformed number of farms at each municipality with cattle farming and the log-transformed number of farms where cattle farming ceased in the last four years. The number of farms where cattle farming was not practiced for more than four years was not available for the whole low Po plain in Lombardy, as this information is not stored in the BCDL, and obtaining it would have implied geo-referencing all the farms in this wide area, which was unpractical.
Statistical methods
At the farm spatial scale, three different models were produced: (1) presence or absence of Barn Swallows was analysed in a binomial generalized linear model (glm) where the factor ‘cattle farming’ was entered as predictor; (2) colony size (i.e. the number of breeding pairs at each farm that hosted a Barn Swallow colony) was modelled according to the same predictor in a zero-truncated Poisson glm (Zuur et al. Citation2009) to assess whether cattle farming influences swallow abundance at those farms which host a colony; and (3) the number of breeding pairs per farm (this also including farms where no swallows were found) was modelled in a Poisson glm with the same independent variable. Models were corrected for data overdispersion using a quasi-likelihood estimation procedure whenever appropriate (Zuur et al. Citation2009). Post-hoc tests (Tukey method) were also performed.
Binomial model performances were assessed using both the proportion of observations correctly classified by the model (overall predictive success, hereafter OPS) and Cohen's kappa (0.41 ≤ Cohen's kappa ≤ 0.60, ‘moderate’ model predictive power; 0.61 ≤ Cohen's kappa ≤ 0.80, ‘substantial’ predictive power; Landis & Koch Citation1993). Overall significance of the zero-truncated Poisson model was assessed by a likelihood ratio test comparison to a null model, i.e. a model including only the intercept (Zuur et al. Citation2009).
To estimate the number of colonies at each municipality in the low Po plain, we first multiplied the proportion of censused farms with swallows in each municipality of the Park by the total number of geo-referenced farms in that municipality. This number, rounded to the closest integer, was then modelled in a Poisson glm where the log-number of farms with cattle farming and the log-number of farms where cattle farming ceased in the last four years were entered as predictors, while the proportion of geo-referenced farms that were surveyed at each municipality was used as a weight to account for different sampling effort. We did not use the number of farms with no cattle farming for more than four years because this information was not available for the whole low Po plain.
The mean number of breeding pairs per colony (mean colony size) at each municipality of the Park was calculated and related to the same predictors as previously in linear regression models weighted for the proportion of farms with colonies among the censused farms. Modelling the mean number of breeding pairs per colony rather than per farm (i.e. excluding censused farms with no swallows) is justified by the aim of predicting the number of breeding pairs in the whole low Po plain in Lombardy, an area for which the number of Barn Swallow colonies could be estimated based on the previous model, while the total number of farms was unknown (see earlier).
We also notice that the total number of colonies and the mean colony size per municipality estimated from the models at the farm spatial scale could not be entered as a dependent variable in the models at the municipality spatial scale; this was because these estimates are a linear function of the number of farms with cattle farming and of the number of farms where cattle farming ceased in the last four years at each municipality that we aimed at entering as predictors in these models. Moreover, estimates from models at the farm spatial scale would not account for the among-municipalities variability in the association between number and size of colonies and number of farms in different livestock categories, which is the focus of the analyses at the municipality spatial scale.
The models of presence and abundance of breeding pairs at each farm and at each colony were cross-validated by randomly dividing the whole dataset into two subsets of equal size. Models with the same predictors as those applied to all farms were then fitted to each data subset and the coefficients thus obtained were used to predict presence and abundance of swallows in the other subset. Performances of the cross-validated binomial glm were assessed by calculating OPS and Cohen's kappa on predicted presence in a subset of farms based on the coefficients of the model from the other subset. Performances of the cross-validated Poisson models were assessed by calculating a cross-validation R 2 (R 2 cv) equal to the proportion of variance in the number of breeding pairs accounted for by a model (i.e. 1 minus the ratio between the sum of the squared differences between the actual number of pairs and that predicted from a model, and the deviance in the actual number of pairs). Models at municipality spatial scale were cross-validated with the same procedure.
The models at the farm spatial scale that passed the cross-validation procedure were applied to all the 682 geo-referenced farms and served to produce a map of the predicted presence and size of breeding colonies in the farms in the municipalities of the Park (. The model at municipality spatial scale was applied to all the 489 municipalities of the low Po plain in Lombardy and the predicted number of colonies at each municipality was also converted into density (i.e. number of colonies per squared kilometre).
Censused municipalities are not a random sample of all the municipalities in the low Po plain in Lombardy, so that models may give biased estimates when extended to other municipalities. For this reason, the map of the predicted density of colonies was validated by correlating estimated densities with the mean number of swallows seen at each point count within a municipality. This procedure is justified by the observation that breeding Barn Swallows forage within 400–500 m from the colony (Møller Citation1994, Ambrosini, Bolzern, Canova, Arieni et al. Citation2002, Turner Citation2006) so that the probability of observing foraging swallows in a randomly located point count increases with the density of colonies in the area. This validation procedure was also applied to subsets of municipalities located at different distances from the Park boundaries, namely <20 km, 20–40 km, 40–60 km, 60–80 km and >80 km. Significance of the correlations was assessed using one-tailed tests because positive correlations between estimated densities and mean number of swallows observed at each point count were expected.
All statistical analyses were performed using r 2.8.1 (R Development Core Team Citation2008); zero-truncated Poisson glms were fitted by the vgam package (Yee Citation2009); post-hoc tests were performed with the multcomp package (Bretz et al. 2001); maps were produced using the sp-package (Pebesma & Bivand Citation2005) and arcview gis 3.2 (ESRI, Redlands, California).
RESULTS
Breeding colonies occurred in 109 out of the 160 (68%) censused farms. The mean number of breeding pairs per farm was 9.29 ± 1.00 se (n = 160), while the mean number of breeding pairs per colony was 13.63 ± 1.26 se (n = 109); 0–81 colonies (mean = 13.61 ± 2.88 se, n = 33) were estimated in the municipalities of the Park, with a within-municipality average colony size of 12.73 ± 1.59 se (n = 27) breeding pairs.
The factor ‘cattle farming’ significantly predicted presence of breeding Barn Swallows in the whole dataset () as well as in both subset A (χ2 2 = 35.00, P < 0.001, Cohen's kappa = 0.608, OPS = 81.25%) and subset B (χ2 2 = 25.07, P < 0.001, Cohen's kappa = 0.544, OPS = 80.00%). The coefficients in show that the probability of the presence of swallows was significantly larger than 0.5 in farms with cattle farming (probability = 0.910, 95% CI = 0.830–0.954, P < 0.001), significantly lower than 0.5 in farms without cattle farming for more than four years (probability = 0.339, 95% CI = 0.232–0.464, P = 0.013), but not significantly different from 0.5 in farms where cattle farming ceased in the last four years (probability = 0.778, 95% CI = 0.421–0.944, P = 0.118). Post-hoc tests revealed a significant difference in the probability of presence between farms with cattle farming and farms without cattle farming for more than four years (z = 6.522, P < 0.001), and a marginal non-significant difference between these latter farms and those where cattle farming ceased in the last four years (z = 2.273, P = 0.056). No significant difference was observed between farms with cattle farming and those where cattle farming ceased in the last four years (z = 1.203, P = 0.439).
Table 1. Binomial, zero-truncated Poisson and Poisson glm (the latter fitted by quasi-likelihood estimation) of the presence, colony size and number of breeding pairs per farm of Barn Swallows.
The model built on subset A correctly predicted presence of breeding Barn Swallows in farms of subset B (Cohen's kappa = 0.544, OPS = 80.00%), and the reciprocal held true as well (Cohen's kappa = 0.608, OPS = 81.25%). Both models predicted presence of Barn Swallows in a farm exactly in the same way, as indicated by the fact that identical statistics were obtained in the cross-validation procedure.
Among the farms with breeding Barn Swallows, those with cattle farming hosted relatively large colonies (15.83 ± 1.58 se pairs, n = 81); colonies of intermediate size were found in farms where cattle farming ceased in the last four years (9.43 ± 2.94 se breeding pairs, n = 7); whereas farms without cattle farming for more than four years hosted the smallest colonies (6.57 ± 1.35 se, n = 21). The factor ‘cattle farming’ significantly predicted mean colony size in the whole set of 109 colonies (), and in both subsets of farms (subset A: n = 54 colonies, χ2 2 = 94.89, P < 0.001, R 2 cv = 0.111; subset B: n = 55 colonies, χ2 2 = 46.33, P < 0.001, R 2 cv = 0.066). In addition, post-hoc tests revealed significant differences between all categories of farms (|z| ≥ 2.412, P ≤ 0.039). However, the cross-validation procedure indicated that the model built on subset B poorly predicted size of colonies in subset A (R 2 cv = 0.053) and the model built on subset A was unable to predict observed colony size in subset B (R 2 cv = –0.030). We note that a negative R 2 cv value can be obtained in this case as predicted values were obtained from a model fitted to a different dataset. Hence, models of colony size could not be used to predict colony size in a larger set of farms.
The mean number of breeding pairs per farm was found to be large in farms with cattle farming (14.40 ± 1.51 se, n = 89), small where cattle farming ceased in the last four years (7.33 ± 2.64 se, n = 9), and minimum in farms without cattle farming for more than four years (2.22 ± 0.60 se, n = 62). The factor ‘cattle farming’ significantly predicted the number of breeding pairs per farm both in the whole dataset () and in the two subsets (subset A: F 2,77 = 19.038, P < 0.001, R 2 cv = 0.271; subset B: F 2,77 = 10.734, P < 0.001, R 2 cv = 0.179). Post-hoc tests revealed a significant difference between farms without cattle for more than four years and farms with cattle (z = 5.952, P < 0.001) and a marginally non-significant difference between farms without cattle for more than four years and those where cattle farming ceased in the last four years (z = 2.275, P = 0.056), while no significant difference existed between farms with cattle and farms where cattle farming ceased in the last four years (z = 1.527, P = 0.267).
The cross-validation procedure indicated that the model built on subset B was able to predict colony size at farms of subset A (R 2 cv = 0.207), while the model built on subset A poorly predicted the number of breeding pairs at colonies of subset B (R 2 cv = 0.063).
Models of presence and number of breeding pairs built on the whole set of farms were finally applied to all the 682 geo-referenced farms (.
The final model of the estimated number of colonies at each municipality in the Park only included the log-transformed number of farms with cattle farming as a significant predictor, with a positive effect (, ). This single variable explained 68% of variation in number of colonies (). The model showed a slight underdispersion (dispersion parameter = 0.746) but, conservatively, we preferred not to correct standard errors estimated for individual parameters.
Table 2. Poisson glm of the estimated number of Barn Swallow colonies per municipality in the Park.
Figure 2. Estimated number of Barn Swallow colonies in relation to the number of farms with cattle farming at each municipality of the Park. The fitted Poisson glm curve from (solid line) and its 95% CLs (dashed lines) are shown.
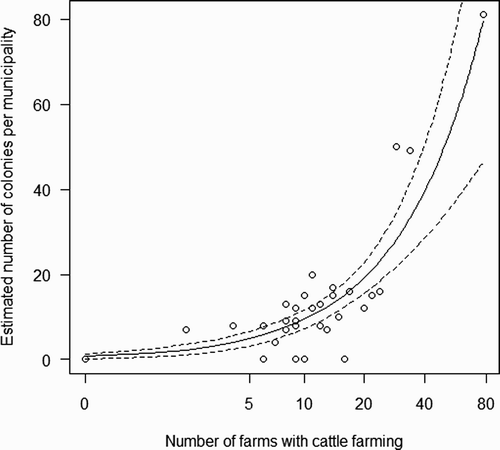
The same final model was obtained in the two subsets of data (subset A: χ2 1 = 47.67, P < 0.001, R 2 cv = 0.789; subset B: χ2 1 = 12.55, P < 0.001, R 2 cv = 0.302). Model cross-validation indicated that the model built on subset A predicted the estimated number of colonies in the municipalities of subset B (R 2 cv = 0.473) and the model built on subset B accurately predicted those in subset A (R 2 cv = 0.879).
Mean colony size was not significantly related to any predictor (P ≥ 0.533; other details not shown) and, therefore, no cross-validation procedure was applied to this model.
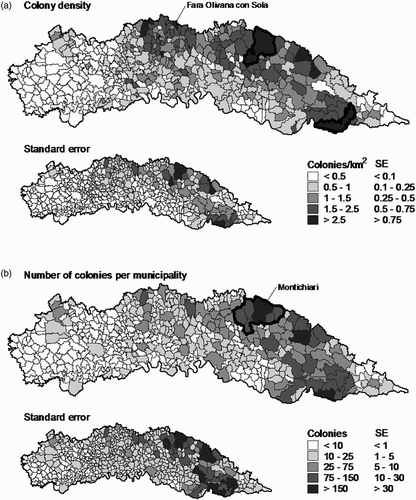
The final model of the estimated number of colonies was then applied to all the municipalities of the low Po plain of Lombardy to obtain the estimated number and density of colonies per municipality represented respectively in , b.
Figure 3. Predicted Barn Swallow distribution at regional scale: (a) predicted density of Barn Swallow colonies; (b) predicted number of Barn Swallow colonies in all the 489 municipalities of the low Po plain. The municipalities with the predicted maximum density (4.16 ± 0.42 se colonies km−2) and the predicted maximum number of colonies (208 ± 67 se) are indicated. Highlighted areas in (a) include 11.7% of colonies in 3.7% of the study area; highlighted area in (b) includes 10.2% of colonies in seven municipalities; small maps represent standard errors of estimates at each municipality.
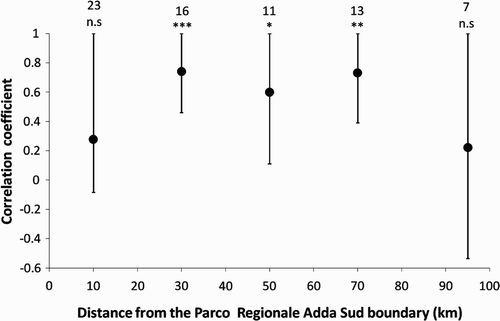
The mean number of swallows recorded at the point counts at each municipality was significantly correlated with the estimated density of colonies (r = 0.527, P < 0.001, one-tailed test, n = 70). In addition, large correlation coefficients were obtained also when selecting only municipalities at different distance belts from the study area, although such correlations were not always significant ().
Figure 4. Correlation coefficients between estimated density of colonies and mean number of Barn Swallow pairs per point count in municipalities at different distance belts (<20 km; 20–40 km; 40–60 km; 60–80 km; and >80 km) from the Park boundary. Bars represent one-sided 95% CLs; numbers represent sample sizes; n.s., P > 0.05; *P ≤ 0.05; **P < 0.01; ***P < 0.001 (one-tailed tests).
DISCUSSION
Models based on the data provided by the BCDL significantly predicted the presence, colony size (i.e. number of breeding pairs in the farms where Barn Swallows were present) and number of breeding Barn Swallows at a farm, and the number of breeding colonies within a municipality of the Park. At farm scale, the models indicated that the probability of presence, the number of breeding pairs and the size of Barn Swallow colonies were largest in farms with cattle farming and smallest in farms with no cattle farming during the four years preceding the census, being intermediate for farms where animal farming ceased during the last four years.
All models had large predictive capability and only the model of colony size failed the cross-validation procedure. The cause of the poor performance of this latter model is probably related to stochastic differences among the two subsets in the mean colony size of farms in different cattle categories, which reduces the ability of the model to predict colony size in the farms of the other subset. The same problem did not occur in the model of the number of breeding pairs per farm, probably because of the larger sample size. The partitioning of the farms among the two subsets may influence the results of the cross-validation procedure. However, explorative analyses in which we re-ran the cross-validation procedure on different subsets always confirmed the poor performance of the model of colony size and the good performance of the other models (details not shown for brevity). We are, therefore, confident that our conclusions on model performance are robust, and would be substantially unchanged if a different partition of the censused farms among the two subsets were used.
The results of the models at farm spatial scale are consistent with those of previous studies, both in the same area (Ambrosini, Bolzern, Canova, Arieni et al. Citation2002) and in northern Europe (Møller Citation1994, Møller Citation2001, Turner Citation2006), that related colony presence and size with presence of livestock farming, particularly cattle. Presence of cattle can determine a large production of flying insects (Møller Citation2001) or be associated with a large extent of meadows and pastures near the colony, so determining abundant food supplies for the insectivorous Barn Swallows (Ambrosini, Bolzern, Canova, Arieni et al. Citation2002, Evans et al. Citation2007). In addition, cowsheds are the favoured nesting sites for the Barn Swallow, and presence of cattle in nesting rooms may mitigate abrupt temperature changes during incubation and nestling rearing (Ambrosini et al. Citation2006, Ambrosini & Saino Citation2010). Hence, presence of cattle can determine favourable conditions for Barn Swallow reproduction (Grüebler et al. Citation2010).
The models at the municipality spatial scale confirmed that the spatial distribution of swallow colonies is tightly related to that of cattle farming also at a larger scale than individual farms. However, no significant relationship could be found between the number of farms with cattle farming and the mean colony size at each municipality.
The models of the presence and the mean number of breeding pairs per farm, and of the number of colonies at each municipality, have large predictive capability. In addition, the fact that data from the same database can be used to predict the distribution of breeding swallows at different spatial scales suggests that the spatial distribution of this passerine bird is determined by the same ecological factors acting at different spatial scales. Indeed, the present results, combined with those from previous studies conducted in the same study area, demonstrate that the presence of livestock farming, and cattle in particular, is the major determinant of swallow distribution at all spatial scales, from nesting micro-environment within a farm (Ambrosini & Saino Citation2010), to colony size (Ambrosini, Bolzern, Canova, Arieni et al. Citation2002, the present study), to large-scale spatial distribution of colonies (the present study).
The distribution of breeding Barn Swallows seems to be related to presence of cattle at a farm, but not to the number of heads reared (Møller Citation2001, Ambrosini, Bolzern, Canova, Arieni et al. Citation2002, Grüebler et al. Citation2010). A reduction in the number of farms with cattle should, therefore, determine a reduction both in the number of colonies and in the mean colony size, even if the number of heads reared at farms that maintain farming remains constant or increases. Hence, the predictive capability of our models should not be affected by the concentration of cattle in rather few larger farms. Conversely, the model's performance may be negatively affected by the general negative trend of Barn Swallow populations in the whole of Lombardy (–4.3% per year; Bani et al. Citation2009), which may lower the number of breeding pairs at farms in all cattle categories, and determine the rapid extinction of small colonies.
The good model performances and the strong consistency in the ecological determinants of Barn Swallow distributions across spatial scales justify the application of our cross-validated models to all the farms in the municipalities of the Park and to all the municipalities in the low Po plain in Lombardy, which represents one of the most important breeding areas for the Barn Swallow in Italy. However, before further discussing the results from the application of our models to large geographical areas, a major caveat deserves attention. The models at municipality spatial scale were based on a rather small number of municipalities that were not randomly chosen, as only the municipalities within the Park were sampled. In addition, the model of the number of colonies per municipality was used to predict colony distribution in a very wide area, so that results may be affected by large uncertainty. Unfortunately, a proper census of breeding colonies in a number of municipalities that could constitute a proper sample of data for model validation would have required an enormous amount of fieldwork that was not practicable. Therefore, we cautiously suggest considering the maps in as tentative, although the correlation between the predicted density of colonies and the number of swallows observed in the point counts gave support to the correctness of the results from the model. In addition, the strong consistency between the ecological determinants of breeding Barn Swallow distribution outlined previously assured the biological realism of our results. However, further extension of this model toward areas with markedly different ecological conditions should be cautiously done, as other ecological determinants such as altitude or climate, may profoundly affect the spatial distribution of breeding Barn Swallow colonies.
The maps at farm spatial scale showed a rather uniform distribution of colonies in the Park with the only exception of the north-eastern zone, where a larger number of colonies reflected the larger number of cattle farms in the area (. The map of the distribution of colonies in the low Po plain showed a west–east increasing trend in colony density, which reflected variation in cattle farming across Lombardy (,b). The model allowed us to estimate 8548 (±172 se) colonies in the low Po plain in Lombardy in 2001. Considering an average colony size of 13.63 (±1.26 se) pairs per colony (see Results), we estimated 116,535(±11,053 se) breeding pairs in the low Po plain of Lombardy in 2001, in good accordance with results of a previous study based on land use that estimated about 140,000 breeding pairs in the whole of Lombardy in the same year (Bani Citation2008). According to the negative demographic trend that this species is suffering in the whole of Lombardy (–4.3% per year, Bani et al. Citation2009), the number of breeding pairs should have declined to about 89,000 (–23.5%) in 2009.
Owing to the sharp decline suffered by their populations in the last years, Barn Swallows may soon become a species of urgent conservation interest. The models we developed may help planning conservation actions for this declining species in the low Po plain. As an exercise, and with the aim of illustrating the potentialities of the present results in the field of conservation biology, we used our models for identifying priority areas for Barn Swallow conservation according to two different criteria. First, under the assumption that resources for biological conservation are limited, we hypothesized that it would be efficient to protect the largest possible number of colonies in the smallest possible area. According to this criterion we identified two different areas, overall covering as few as 328 km2 (3.7% of the study area; , that included as many as 1004 (11.7%; 95% CI: 591–1717) colonies. An alternative criterion would be to select the smallest number of contiguous municipalities that include at least 10% of colonies. This might be efficient if, for example, municipality regulations were needed to implement conservation strategies. The area so identified according to our models hosts 878 colonies (10.2%; 95% CI: 518–1496) in as little as 346 km2 (corresponding to 3.9% of the whole study area; . These results should be further validated with intense surveys of breeding colonies in focal areas. Nevertheless, they allow a first assessment of potentially suitable areas for Barn Swallow reproduction and are, therefore, of primary interest in actions aimed at efficient conservation of breeding colonies.
Finally, we stress that the independent variables entered in the models at both spatial scales at which the analyses were run were obtained from a bovine computerized database, which is mandatory in the EU according to the Regulation (CE) 1760/2000. Similar information should, therefore, be available in all the countries of the EU, so that our models may serve as a basis for wide-scale modelling of breeding Barn Swallow distribution and, therefore, contribute to properly designing conservation plans for this species at continental level. Hence, this paper adds a totally novel contribution to the current knowledge of the ecology and distribution of Barn Swallows, as it shows that the same ecological factors act at different scales in determining the distribution of this species. In addition, biologists studying the conservation of this declining species can profit from existing publicly available information to focus better their population monitoring and conservation actions.
ACKNOWLEDGEMENTS
We are grateful to the Unità Organizzativa Veterinaria, Direzione Generale Sanità, of the Regione Lombardia for providing the bovine computerized database data, to the FaunaViva association for providing the MITO2000 data and to all farm owners that allowed us to census swallows in their barns and houses. Michele Paba, Francesca Addario, Silvia Arieni, Barbara Fabbo, Alessandra Galleni and Daria Tosi gave an invaluable contribution to fieldwork. The Parco Regionale Adda Sud financially supported the Barn Swallow census. We also thank the Unità Organizzativa Multifunzionalità e Sostenibilità del Territorio, Direzione Generale Agricoltura, of the Regione Lombardia for financial support.
REFERENCES
- Ambrosini , R. and Saino , N. 2010 . Environmental effects at two nested spatial scales on habitat choice and breeding performance of barn swallow . Evol. Ecol. , 24 : 491 – 508 .
- Ambrosini , R. , Bolzern , A. M. , Canova , L. , Arieni , S. , Møller , A. P. and Saino , N. 2002 . The distribution and colony size of barn swallows in relation to agricultural land use . J. Appl. Ecol. , 39 : 524 – 534 .
- Ambrosini , R. , Bolzern , A. M. , Canova , L. and Saino , N. 2002 . Latency in response of barn swallow Hirundo rustica populations to changes in breeding habitat conditions . Ecol. Lett. , 5 : 640 – 647 .
- Ambrosini , R. , Ferrari , R. P. , Martinelli , R. , Romano , M. and Saino , N. 2006 . Seasonal, meteorological, and microhabitat effects on breeding success and offspring phenotype in the barn swallow . Hirundo rustica. Ecosci. , 13 : 298 – 307 .
- Bani , L. 2008 . “ Rondine – Swallow ” . In Hirundo rustica, [Wildlife in Lombardy. 2008 Report on Distribution, Abundance and Conservation Status of Birds and Mammals]: 120 Edited by: Vigorita , V. and Cucè , L. Regione Lombardia – DG Agricoltura, Università degli Studi di Milano-Bicocca, Università degli Studi dell'Insubria, Università degli Studi di Pavia, Milan. (In Italian)
- Bani , L. , Massimino , D. , Orioli , V. , Bottoni , L. and Massa , R. 2009 . Assessment of population trends of common breeding birds in Lombardy, Northern Italy, 1997–2007 . Ethol. Ecol. Evol. , 21 : 27 – 44 .
- Bretz, F., Genz, A. & Horton, L. 2001. On the numerical availability of multiple comparison procedures. Biometrical J. 43: 645–656.
- Butler , S. J. , Vickery , J. A. and Norris , K. 2007 . Farmland biodiversity and the footprint of agriculture . Science , 315 : 381 – 383 .
- Cadotte , M. W. and Fukami , T. 2005 . Dispersal, spatial scale, and species diversity in a hierarchically structured experimental landscape . Ecol. Lett. , 8 : 548 – 557 .
- Denny , M. W. , Helmuth , B. , Leonard , G. H. , Harley , C. D. , Hunt , L. J.H. and Nelson , E. K. 2004 . Quantifying scale in ecology: lessons from a wave-swept shore . Ecol. Monog. , 74 : 513 – 532 .
- Donald , P. F. , Green , R. E. and Heath , M. F. 2001 . Agricultural intensification and the collapse of Europe's farmland bird populations . Proc. R. Soc. Lond. Ser. B: Biol. Sci. , 268 : 25 – 29 .
- Evans , K. L. , Wilson , J. D. and Bradbury , R. B. 2007 . Effects of crop type and aerial invertebrate abundance on foraging barn swallows Hirundo rustica . Agric. Ecosyst. Environ. , 122 : 267 – 273 .
- Fielding , A. H. and Haworth , P. F. 1995 . Testing the generality of bird–habitat models . Conserv. Biol. , 9 : 1466 – 1481 .
- Fornasari , L. , Bani , L. , de Carli , E. and Massa , R. 1998 . Optimum design in monitoring common birds and their habitat . Gibier Faune Sauvage Game Wildl , 15 ( 2 ) : 309 – 322 .
- Fornasari , L. , de Carli , E. , Brambilla , S. , Buvoli , L. , Maritan , E. and Mingozzi , T. 2002 . [Breeding bird distribution in Italy. First report of the MITO2000 monitoring program] . Avocetta , 26 : 59 – 115 . (In Italian)
- Grüebler , M. U. , Korner-Nievergelt , F. and von Hirschheydt , J. 2010 . The reproductive benefits of livestock farming in barn swallows Hirundo rustica: quality of nest site or foraging habitat? . J. Appl. Ecol. , 47 : 1340 – 1347 .
- Landis , J. R. and Koch , G. G. 1993 . The measurements of observer agreement for categorical data . Biometrics , 33 : 159 – 174 .
- Møller , A. P. 1994 . Sexual Selection and the Barn Swallow , Oxford , , UK : Oxford University Press .
- Møller , A. P. 2001 . The effect of dairy farming on barn swallow Hirundo rustica abundance, distribution and reproduction . J. Appl. Ecol. , 38 : 378 – 389 .
- Pebesma , E. J. and Bivand , R. S. 2005 . Classes and methods for spatial data in R . R News , 5 : 9 – 13 .
- R Development Core Team . 2008 . R: a Language and Environment for Statistical Computing , Vienna , , Austria : R Foundation for Statistical Computing .
- Rossetti , G. , Pieri , V. and Martens , K. 2005 . Recent ostracods (Crustacea, Ostracoda) found in lowland springs of the provinces of Piacenza and Parma (Northern Italy) . Hydrobiologia , 542 : 287 – 296 .
- Tucker , G. M. and Heath , M. F. 1994 . Birds in Europe: Their Conservation Status , Cambridge , , UK : Birdlife International .
- Turner , A. 2006 . The Barn Swallow , London : T & A D Poyser .
- Vaugan , I. P. and Ormerod , S. J. 2003 . Improving the quality of distribution models for conservation by addressing shortcomings in field collection of training data . Conserv. Biol. , 17 : 1601 – 1611 .
- Wiens , J. A. , Rotenberry , J. T. and Van Horne , B. 1986 . A lesson in the limitation of field experiments: shrub-steppe birds and habitat alteration . Ecology , 67 : 365 – 376 .
- Yee , T. W. 2009 . VGAM: Vector Generalized Linear and Additive Models. R package version 0 . : 7 – 9 . http://www.stat.auckland.ac.nz/~yee/VGAM
- Zuur , A. F. , Ieno , E. I. , Walker , N. J. , Saveliev , A. A. and Smith , G. M. 2009 . Mixed Effects Models and Extensions in Ecology with R , New York : Springer .