Abstract
Capsule Key marginal habitat features maintain their importance even when they occur at very low density.
Aims To assess the importance for breeding birds of key habitat elements, such as isolated shrubs, hedgerows and untilled vegetation patches.
Methods We investigated the habitat preferences of Red-backed Shrikes Lanius collurio in an agricultural environment where key habitat elements have been dramatically reduced. Based upon data from territorial maps, a fine-scale model of habitat preference was developed and the results of three different analytical approaches (binary logistic regression, multimodel inference and hierarchical partitioning) were compared.
Results The occurrence of Red-backed Shrikes was positively influenced by the extent of non-grazed/mown grassland, isolated bushes and hedgerows, and negatively influenced by woodland cover. The model highlighted the importance of bushes/hedgerows despite their very scarce occurrence in the study areas (overall, average cover only 7.94%). Breeding densities were rather low, but mean densities were slightly higher in pasture zones in which bushes/hedgerows availability was higher than elsewhere.
Conclusion These results confirm the crucial importance of these key marginal elements even in depauperated farmland landscapes. Maintaining and increasing their availability is a key management option for the encouragement of breeding Red-backed Shrikes, and potentially for other species which are also declining in these farmland habitats.
In the second half of the 20th century the European populations of many bird species which utilize farmland have undergone rapid decline (Tucker & Evans Citation1997, Pain & Pienkowski Citation1997, Gregory et al. Citation2008). The most important causes of this decline have been recognized in the intensification of agriculture, encouraged by national and European policies (Common Agricultural Policy) to increase agricultural yields (Pain & Dixon Citation1997, Donald et al. Citation2001; Stoate et al. Citation2009), with population declines most marked in countries where agricultural intensification has been longer and more pervasive (Donald et al. Citation2001, Geiger et al. Citation2010). The increase of agriculture production has been reached through mechanization, use of chemical products and removal of many ‘unproductive’ marginal elements of the landscape such as hedgerows, bushes, trees and uncultivated or semi-natural lands. These marginal elements represent key habitats for many species and are probably more important for farmland bird populations than landscape heterogeneity per se (Heikkinen et al. Citation2004, Batáry et al. Citation2011, Kleijn et al. Citation2011). The availability of good foraging habitats close to safe nesting sites is a critical factor for birds breeding on farmland (Benton et al. Citation2002), and many farmland bird species are associated with marginal features (see e.g. Brambilla et al. Citation2008, 2009).
The other side of the deep changes in agriculture practices is represented by the abandonment of many areas where agricultural intensification was not possible or too expensive. Land abandonment is leading to a dramatic loss of habitat for many species tied to open and semi-open landscapes (Pain & Pienkowski Citation1997, Brambilla et al. 2010b).
The aim of this article is to investigate the importance of marginal habitat features for threatened species, by studying habitat preferences of Red-backed Shrikes Lanius collurio in Trentino, Italian Alps. The study region still hosts an appreciable population of Red-backed Shrikes; however, many habitat traits usually selected by this species are poorly represented, having been removed by intensification and/or abandonment in many areas. Therefore, our study region qualifies as an ideal site to assess the importance of residual, marginal features in depauperated landscapes.
Red-backed Shrikes are particularly sensitive to both agricultural intensification and land abandonment (Brambilla et al. Citation2007), and they appear to be tied to the marginal features accompanying cultivated or grazed areas (Brambilla et al. Citation2009, Brambilla & Ficetola Citation2012). The populations of this species underwent a strong decline between the 1970s and the 1990s, disappearing from large areas (Lefranc Citation1997), while a short-term slightly positive trend is reported from the Pan-European Common Bird Monitoring Scheme for the period 1990–2003 (PECBMS Citation2008). By investigating habitat preferences in a population inhabiting a landscape which has undergone impoverishment of those habitat resources that appear to be essential for Red-backed Shrikes, we hoped to assess the importance of marginal features. If such marginal features are key resources even if scarce, habitat selection in the study species should be driven by the availability of those features; otherwise, if marginal features lose their importance when severely depleted, habitat suitability would be affected by other elements.
METHODS
Study area
The Trento Province (N. Italy, 6206 km2, elevation 67–3769 m asl) includes part of the Central and Eastern Italian Alps. The terrain is mainly mountainous (20 % is over 2000 m asl) with extensive woodland (56 % of the total surface, Provincia Autonoma di Trento Citation2007). Urbanized and cultivated areas are concentrated in the valleys. In the last 50 years the land surface area exploited for farmland activities has declined at middle–high elevation; at the same time, agricultural intensification at low elevation has led to an uniform landscape of high-intensity vineyards and orchards in large parts of the valleys. The area of meadows and pastures decreased from 148 769 ha in 1970 to 120 161 ha in 2000, while arable land declined from 7662 ha to 3678 ha (Provincia Autonoma di Trento, in Pedrini et al. Citation2005). Intensification in valley floors and abandonment at middle elevations are probably the main causes of the decline of birds on farmland in Trentino.
Some hundreds of Red-backed Shrike pairs (Pedrini et al. Citation2005) still breed in the remaining grassland areas, within a broad altitudinal range, with important sites in valley floors, middle elevations and uplands.
Fourteen study areas were chosen within the Trento province (6 in 2009; 8 in 2010). These were situated in locations where species occurrence was already known from large-scale breeding bird distribution data (Pedrini et al. Citation2005). Study areas included open and semi-open habitats, such as grasslands, pastures, herbaceous cultivations, but also portions of orchards, vineyards, small woodlots, sparse buildings. Study areas were delimited by discontinuities in the landscape, such as edges of large woodlands, large roads, urban habitats and rivers. Each area (range: 18.8–116.9 ha) was large enough to host potentially from a few to several breeding pairs of Red-backed Shrikes (Sutherland Citation2006). Study areas include all the important habitats for the species in Trentino (meadows, pastures, cultivated lands) in a broad altitudinal range (200–1800 m asl).
Fieldwork
Territory mapping
Each study area was surveyed at slow pace two to five times. The number of visits varied according to area, terrain, number of contacts with the target species. All contacts were plotted on detailed aerial photographs, recording all territorial and breeding behaviours indicating site occupancy; these included nests, male and female seen together, singing male, aggressive encounters, calling of juveniles (Sutherland Citation2006). Simultaneous observations of different pairs or males are particularly useful for the definition of the position and shape of the breeding territories of the species (Karlsson Citation2004; Brambilla et al. Citation2007, Citation2009).
Surveys were carried out in the morning (06:00–12:00 hours), between 1 June and 15 July in both years, to avoid possible recording of migrant (passage) birds which are still common in May in northern Italy (Brambilla et al. Citation2009). A lower number of individuals was observed during the first ten days of June of both years, probably because of later arrivals and/or settlements, which were also recently reported for other areas (Brambilla et al. Citation2009), and in accordance with the ringing data available for Italy (Spina & Volponi Citation2008).
Recording habitat variables
Thirty-two detailed habitat variables () not available as geographic information system (GIS) layers were directly measured in the field. After territory mapping, the habitat variables were measured within 1 ha (100 m × 100 m square) territory and control squares; 1 ha approximates the average size of breeding territory of the Red-backed Shrikes in the study region (Brambilla et al. Citation2007, Citation2009). Squares were placed so as to cover most of the area defended and used by each pair. Within each study area, an equal number of control squares was randomly scattered over areas known to be unoccupied by the species and with tree cover lower than 50 % of the area; sites with a higher tree cover would be clearly unsuitable for the species, and were therefore unsuitable as control squares for modelling fine-scaled habitat preferences (Brambilla et al. Citation2009).
Table 1. Habitat variables measured in the field within the 100 m × 100 m territory and control squares.
Habitat variables were measured during the last territory mapping sessions and during dedicated surveys (mid-July–August). Topographic variables (elevation, aspect and slope) were calculated in a GIS environment as mean values for each square by using the Digital Terrain Model of the Trento Province.
Statistical analysis
The data collected during 2009 were used to develop a first fine-scale model. The data set was simplified by reducing the number of factors. Habitat variables which occurred in less than 8 sample squares considering both territory and control squares were removed. These were: coverage of dry grassland, maize, cultivations, bramble bushes, heath, uncultivated land, lucerne, vineyards, orchards, urbanized areas, rocky areas, bare soil, waterbodies, wetlands, tree plantations and coverage and length of tree rows, paved roads, fences.
Habitat cover variables were square-root-arcsine transformed (Fowler & Cohen Citation2002). Aspect (a circular variable) was reclassified for obtaining a linear factor using a score reclassification: 330–360° and 0–30°: north; 30–60°: northeast; 60–90°: east-northeast; 90–120°: east-southeast; 120–150°: southeast; 150–210°: south; 210–240°: south-southwest; 240–270°: west-southwest; 270–300°: west-northwest; 300–330°: northwest (Brambilla et al. Citation2009).
A t-test was carried out for a comparison between mean values of habitat variables in territory and control squares.
To assess the relative importance of habitat variables on the probability of species occurrence within a sample square (modelled by comparing occupied versus unoccupied squares), three alternative statistical procedures were carried out and compared: stepwise binary logistic regression, multimodel inference, and hierarchical partitioning. We avoided including strongly intercorrelated factors (r > 0.6) in model selection. The factors which were indicated to be the most important according to the three analyses were used to build two models through logistic regression analysis (procedure enter in SPSS).
Logistic regression was carried out in SPSS for Windows 17.0 (Norušis Citation2009) using a stepwise backward procedure, which sequentially excludes non-significant factors (P > 0.1) on the basis of the likelihood ratio test.
Multimodel inference, based on an information-theoretic approach (Anderson et al. Citation2000, Anderson Citation2008), was performed on a set of 12 models. The set was built including 2–4 habitat variables in each model in such a way that every variable was considered at least once (Merli & Meriggi Citation2006). Akaike's Information Criterion corrected for small samples (AICC), differences between AICC values (Δ i ) and Akaike weights w were calculated for each model in order to identify the ‘best’ models in the set. The area under the curve (AUC) of the Receiver Operating Characteristic (ROC) plots was used to assess the discrimination ability of the models (Pearce & Ferrier Citation2000). This value varies between 0.5 (discrimination ability no better than random) and 1.0 (perfect discrimination ability; Pearce & Ferrier Citation2000). Model averaging was carried out on the seven best models to assess the relative importance of the habitat variables. For each factor we calculated: the sum of the Akaike weights of the models which included the variable, the regression coefficient β weighted and averaged on the models and the unconditional sampling variance with 95 % confidence interval (Anderson et al. Citation2000).
Hierarchical partitioning analysis was used to check the results of the previous analysis (Brambilla et al. Citation2010a). This method was carried out in r 2.12 (R Development Core Team Citation2010) by means of hier.part package (Walsh & Mac Nally Citation2009) and allowed the estimation of both independent (I) and joint (J) explanatory power of each variable, considering all the 2K possible models (Mac Nally Citation2000).
Checking the validity of models
The data collected in 2010 in eight new areas were classified according to the models obtained from the 2009 data. The discrimination ability of the models over the new data set was tested by calculating the AUC of the ROC plots.
Breeding densities
Breeding densities were calculated as number of breeding pairs per 10 ha per study area (). To assess the potential effect of habitat variables on breeding density, we first checked for possible correlations between breeding densities and each predictor (calculated as mean percentage value over both control and territory squares of each study area), and second, checked for an effect of the cumulative percentages of hedgerows, bushes and isolated bushes (summarized to obtain one factor representing all nesting and perching elements) on the breeding density. Finally, as we noted that areas with pastures hosted on average a higher number of marginal elements than areas without pastures (11.0 % versus 5.2 %; t = –2.74, P = 0.018), we checked for a difference in breeding densities also between areas with and without pastures.
Table 2. Breeding densities of Red-backed Shrikes in the 14 study areas in Trento Province (Italian Alps) in 2009 and 2010.
RESULTS
Thirty-three breeding pairs were recorded in 2009 in 6 study areas, and 42 in 2010 in the other 8 study areas. Comparing the mean values of habitat variables in territory and control squares, there was a significant difference with respect to non-grazed/mown grassland (t = –2.59, P = 0.014) and woodland (t = 1.55, P = 0.042) in the 2009 data set, and for non-grazed/mown grassland (t = –2.53, P = 0.014), isolated bushes (t = –4.03, P < 0.001), unpaved roads (t = 2.10, P = 0.041) and length of unpaved roads (t = 2.27, P = 0.027) in the 2010 data set. In territory squares the mean values of non-grazed/mown grassland and isolated bushes were higher than in control squares, whereas the opposite was true for the mean values of woodland and unpaved roads, which were lower in territory squares.
The output of stepwise binary logistic regression was the following equation (8th step of the backward stepwise procedure):
Table 3. The set of 12 models used for multimodel inference ranked according to their AICc values. ngmg = non-grazed/mowed grassland, b = bushes, ib = isolated bushes, w = wood, mg = mowed grassland, h = hedgerows, a = aspect, e = elevation, ur = unpaved roads, s = slope, gg = grazed grassland, it = isolated trees.
Model averaging allowed us to determine the information of all models considered (i.e. the seven best models of the previous set in this specific case). Results showed a clear effect of four variables on the occurrence of the species: non-grazed/mown grassland, bushes and isolated bushes (positive effect), and woodland (negative effect; ). The influence of hedgerows is less clear and less strong, because of the low averaged β and the confidence interval which includes 0. Values of the other two variables (mown grassland and aspect) are much lower; therefore, the variables with appreciable influence are the same as those included in logistic regression equation.
Table 4. Effect of seven habitat variables on the occurrence of Red-backed Shrikes as estimated from multimodel inference. The effect of aspect and mown grassland is negligible (confidence intervals of β encompass 0). The u.s. var. is the unconditional sampling variance.
Hierarchical partitioning analysis substantially confirmed the results of the previous analyses (). The five variables with higher independent contribution are the same as those included in the logistic regression equation. It was necessary to exclude two variables because with 11 factors the analysis produced a rounding error (a potential bias already mentioned in hier.part package guide, Walsh & Mac Nally Citation2009). Isolated trees and slope were excluded because of their low importance as pointed out by the previous analyses.
Table 5. Independent contribution (I) of single habitat variables on the occurrence of Red-backed Shrikes as estimated from hierarchical partitioning
On the basis of the outcome of the three different procedures, two models were developed: the first, including all the five variables which were indicated to be important, is the same as that found from the logistic regression analysis (see above), and had an AUC equal to 0.84 (good discrimination power). A second and more synthetic model was built excluding hedgerows, because of the lower importance resulted in multimodel inference:
Model validation
Applying the 2009 models to the 2010 data set, their discrimination capacity was found to be lower with respect to the original data (2009 data set). Therefore, the coefficients of the variables were re-calculated by applying the model to the entire data set. Two new equations were thus obtained:
Breeding densities and residual marginal elements
The five variables included in the best model occupied a very low area of the squares (), with the exception of non-grazed/mown grassland which had high percentages in some partially abandoned areas. The other elements with positive effect on the occurrence of the species (isolated bushes, bushes and hedgerows) which are used as breeding sites and hunting perches, had a very low cover in both territory and in control squares.
Table 6. Mean values ± standard error of the habitat variables included in the best model calculated for territory squares, control squares and the entire data set.
No significant correlation was found between bird breeding density and the single habitat variables (all P > 0.12), while the correlation between breeding density and the factor representing the sum of nesting and perching elements was marginally significant (r = 0.49, P = 0.079).
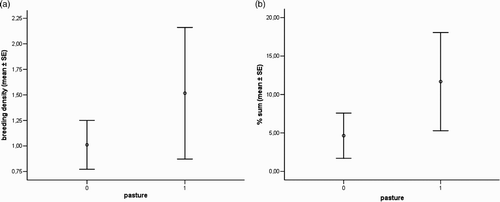
Consistent with the higher cover of marginal elements in pastures, breeding densities were marginally higher in study areas with pastures (mean = 1.52 pairs/10 ha) than without (mean = 1.01 pairs/10 ha; t = –2.06, P = 0.061) ().
Figure 1. Left: mean values (±1 se) of breeding density (pairs per 10 ha) of Red-backed Shrikes in study areas with (1) or without (0) pasture. Right: mean % values (±1 se) of hedgerows and bushes (sum) in study areas with (1) or without (0) pasture.
DISCUSSION
The fine-scale models obtained using three different approaches showed a good discrimination ability over the two completely independent data sets. They highlighted the importance of five habitat variables, four of which positively affected the probability of Shrike occurrence. Only one of the variables which are important for Shrikes, the non-grazed/mown grassland, was represented well within the whole area of the study. This habitat represents a source of invertebrate prey for Red-backed Shrikes (Tryjanowski et al. Citation2003, Casale & Brambilla Citation2009). Hedgerows, bushes and isolated bushes were included in the model of habitat preferences despite their scarcity in the whole area of the study (overall average cover = 7.94%), and all have a strong influence on the probability of the occurrence of Red-backed Shrikes according to the regression coefficients of the best ‘final’ model. Results showed that even a very limited availability of these marginal features is enough to influence the occurrence of breeding pairs of Red-backed Shrikes. Considering the surveyed areas, portions with bushes and hedgerows represent islands of suitable sites scattered within wider grassland areas (which are also islands at a different scale, given that in the region of the study below 2000 m asl the land is mainly covered by woodlands and intensively managed orchards and vineyards). The scarcity of bushes and hedgerows is the likely reason for the rather low breeding density. Breeding densities were higher in pasture areas, where the availability of the key habitat factors was significantly higher; grazed habitats often qualify as optimal habitats for Red-backed Shrikes, because of high food and nesting sites availability (Lefranc Citation1997; Brambilla et al. Citation2007). Similar studies carried out in a neighbouring region (Lombardy) reported much higher cover values of shrubs and hedgerows in both territory and control plots for Red-backed Shrikes (Brambilla et al. Citation2009, Citation2010b) in otherwise comparable habitats (same elevation and land use), with associated much higher breeding densities of Red-backed Shrikes (up to 10 pairs/10 ha; Casale & Brambilla 2009). This indirectly suggests how scarcity of these habitat features may actually limit breeding densities in our study areas.
These findings have potentially important conservation implications, as they suggest that even limited (and not expensive) efforts to ensure a minimal cover of these key factors in farmland areas (by maintaining or restoring bushes and hedgerows) could have positive impact on the density of Red-backed Shrikes. These findings suggest that even highly simplified grasslands could become attractive to Red-backed Shrikes with suitable management of these marginal habitat features. The importance of grassland areas with untilled herbaceous vegetation was also confirmed. Red-backed Shrikes positively respond to the introduction of management promoting untilled herbaceous vegetation (Casale & Brambilla Citation2009), and this would also have positive effects on the occurrence of many other farmland species (Casale & Brambilla Citation2009).
In conclusion, our work further confirms the importance of marginal habitat elements in farmed landscapes, suggesting that they can play a crucial function even though they may be scarce. We suggest that the maintenance or increase of such habitat elements should be encouraged and promoted also in depauperated landscapes.
ACKNOWLEDGEMENTS
We are grateful to C. Tattoni, F. Rizzolli and M. Segata for kind help with the work and to V. Fin and L. Sottovia for support. The present work has been partly financed by the Autonomous Province of Trento in the context of the Rural Development Programme and of the Natura 2000 network.
REFERENCES
- Anderson , D. R. 2008 . Model Based Inference in the Life Sciences: A Primer on Evidence , New York : Springer .
- Anderson , D. R. , Burnham , K. P. and Thompson , W. L. 2000 . Null hypothesis testing: problems, prevalence, and an alternative . J. Wildl. Manage. , 64 : 912 – 923 .
- Anderson , D. R. , Link , W. A. , Johnson , D. H. and Burnham , K. P. 2001 . Suggestions for presenting the results of data analyses . J. Wildl. Manage. , 65 : 373 – 378 .
- Batáry , P. , Báldi , A. , Kleijn , D. and Tscharntke , T. 2011 . Landscape-moderated biodiversity effects of agri-environmental management: a meta-analysis . Proc. R. Soc. Lond. B , DOI:10.1098/rspb.2010. 1923
- Benton , T. G. , Bryant , D. M. , Cole , L. and Crick , H. Q.P. 2002 . Linking agricultural practice to insect and bird populations: a historical study over three decades . J. Appl. Ecol , 39 : 673 – 687 .
- BirdLife International . 2004 . Birds in Europe. Population Estimates, Trends and Conservation Status , Cambridge : BirdLife International .
- Brambilla , M. and Ficetola , G. F. 2012 . Species distribution models as a tool to estimate reproductive parameters: a case study with a passerine bird species . J. Anim. Ecol. , DOI: 10.1111/j.1365-2656. 2012.01970.x
- Brambilla , M. , Rubolini , D. and Guidali , F. 2007 . Between land abandonment and agricultural intensification: habitat preferences of Red-backed Shrikes Lanius collurio in low-intensity farming conditions . Bird Study , 54 : 160 – 167 .
- Brambilla , M. , Guidali , F. and Negri , I. 2008 . The importance of an agricultural mosaic for Cirl Buntings Emberiza cirlus in Italy . Ibis , 150 : 628 – 632 .
- Brambilla , M. , Casale , F. , Bergero , V. , Crovetto , G. M. , Falco , R. , Negri , I. , Siccardi , P. and Bogliani , G. 2009 . GIS-models work well, but are not enough: habitat preferences of Lanius collurio at multiple levels and conservation implications . Biol. Conserv. , 142 : 2033 – 2042 .
- Brambilla , M. , Bassi , E. , Ceci , C. and Rubolini , D. 2010a . Environmental factors affecting patterns of distribution and co-occurrence of two competing raptor species . Ibis , 10 : 1 – 12 .
- Brambilla , M. , Casale , F. , Bergero , V. , Bogliani , G. , Crovetto , G. M. , Falco , R. , Roati , M. and Negri , I. 2010b . Glorious past, uncertain present, bad future? Assessing effects of land-use changes on habitat suitability for a threatened farmland bird species . Biol. Conserv. , 143 : 2770 – 2778 .
- Casale , F. and Brambilla , M. 2009 . Averla piccola. Ecologia e conservazione , Milano : Fondazione Lombardia per l'Ambiente e Regione Lombardia .
- Donald , P. F. , Green , R. E. and Heath , M. F. 2001 . Agricultural intensification and the collapse of Europe's farmland bird populations . Proc. R. Soc. Lond. B , 268 : 25 – 29 .
- Fowler , J. and Cohen , L. 2002 . Statistica per ornitologi e naturalisti , Roma : Franco Muzzio Editore .
- Geiger , F. , Bengtsson , J. , Berendse , F. , Weisser , W. W. , Emmerson , M. , Morales , M. B. , Ceryngier , P. , Liira , J. , Tscharntke , T. , Winqvist , C. , Eggers , S. , Bommarco , R. , Pärt , T. , Bretagnolle , V. , Plantegenest , M. , Clement , L. W. , Dennis , C. , Palmer , C. , Oñate , J. J. , Guerrero , I. , Hawro , V. , Aavik , T. , Thies , C. , Flohre , A. , Hänke , S. , Fischer , C. , Goedhart , P. W. and Inchausti , P. 2010 . Persistent negative effects of pesticides on biodiversity and biological control potential on European farmland . Basic Appl. Ecol. , 11 : 97 – 105 .
- Gregory , R. D. , Vořišek , P. , Noble , D. G. , Van Strien , A. , Klvaňová , A. , Eaton , M. , Gmeling Meyling , A. W. , Joys , A. , Foppen , R. P.B. and Burfield , I. J. 2008 . The generation and use of bird population indicators in Europe . Bird Conserv. Int. , 18 : S223 – S244 .
- Heikkinen , R. K. , Luoto , M. , Virkkala , R. and Rainio , K. 2004 . Effects of habitat cover, landscape structure and spatial variables on the abundance of birds in an agricultural–forest mosaic . J. Appl. Ecol. , 41 : 824 – 835 .
- Karlsson , S. 2004 . Season-depended diet composition and habitat use of Red-backed Shrikes Lanius collurio in SW Finland . Ornis Fenn. , 81 : 97 – 108 .
- Kleijn , D. , Rundlöf , M. , Scheper , J. , Smith , H. G. and Tscharntke , T. 2011 . Does conservation on farmland contribute to halting the biodiversity decline? . Trends Ecol. Evol. , 26 : 474 – 481 .
- Lefranc , N. 1997 . “ Shrikes and the farmland landscape in France ” . In Farming and Birds in Europe: The Common Agricultural Policy and its Implications for Bird Conservation , Edited by: Pain , D J and Pienkowski , M W . 236 – 268 . London : Academic Press .
- Mac Nally , R. 2000 . Regression and model-building in conservation biology, biogeography and ecology: The distinction between – and reconciliation of – ‘predictive’ and ‘explanatory’ models . Biodiversity Conserv. , 9 : 655 – 671 .
- Merli , E. and Meriggi , A. 2006 . Using harvest data to predict habitat–population relationship of the wild boar Sus scrofa in Northern Italy . Acta Theriol. , 51 : 383 – 394 .
- Norušis , M. J. 2009 . SPSS Statistics 17.0 Guide to Data Analysis , New York : Prentice Hall .
- Pain , D. J. and Dixon , J. 1997 . “ Why farming and birds in Europe? ” . In Farming and Birds in Europe: The Common Agricultural Policy and its Implications for Bird Conservation , Edited by: Pain , D. J. and Pienkowski , M W . 1 – 24 . London : Academic Press .
- Pain , D. J. and Pienkowski , M. W . 1997 . Farming and Birds in Europe: The Common Agricultural Policy and its Implications for Bird Conservation , Edited by: Pain , D. J. and Pienkowski , M.W . London : Academic Press .
- PECBMS 2008. Trends of common farmland birds in Europe. http://www.birdlife.cz/index.php?ID=1320. Accessed 12 October 2011.
- Pearce , J. and Ferrier , S. 2000 . Evaluating the predictive performance of habitat models developed using logistic regression . Ecol. Model , 133 : 225 – 245 .
- Pedrini , P. , Caldonazzi , M. and Zanghellini , S . 2005 . Atlante degli uccelli nidificanti e svernanti in provincia di Trento , Edited by: Pedrini , P. , Caldonazzi , M. and Zanghellini , S . Trento : Museo Tridentino di Scienze Naturali .
- Provincia Autonoma di Trento. 2007. Uso del suolo attuale e raffronto storico. http://www.foreste.provincia.tn.it/tabelle %20per %20internet/a_1 %20NEW.pdf. Accessed 6 August 2010.
- R Development Core Team . 2010 . R: A Language and Environment for Statistical Computing , Vienna : R Foundation for Statistical Computing .
- Spina , F. and Volponi , S. 2008 . Atlante della Migrazione degli Uccelli in Italia. 2. Passeriformi , Roma : Ministero dell'Ambiente e della Tutela del Territorio e del Mare, Istituto Superiore per la Protezione e la Ricerca Ambientale (ISPRA) .
- Stoate , C. , Báldi , A. , Beja , P. , Boatman , N. D. , Herzon , I. , Van Doorn , A. , De Snoo , G. R. , Rakosy , L. and Ramwell , C. 2009 . Ecological impacts of early 21st century agricultural change in Europe – a review . J. Environ. Manage. , 91 : 22 – 46 .
- Sutherland , W. J. 2006 . Ecological Census Techniques – A Handbook , 2 , Cambridge : Cambridge University Press .
- Tryjanowski , P. , Karg , M. K. and Karg , J. 2003 . Diet composition and prey choice by the Red-backed Shrike Lanius collurio in western Poland . Belg. J. Zool. , 133 : 157 – 162 .
- Tucker , G. M. and Evans , M. I. 1997 . Habitats for Birds in Europe: A Conservation Strategy for the Wider Environment , Cambridge : BirdLife International .
- Walsh, C. & Mac Nally, R. 2009. Package hier.part. http://cran.stat.unipd.it. Accessed 21 February 2010.