Abstract
Capsule The use of plasticine eggs coated with a thin layer of rubber instead of entirely plasticine eggs in dummy nests reduced the unnaturally high nest predation rate caused by attracted small mammals, while also enabling the identification of nest predators.
Artificial nests are widely used for estimating the predation pressure on avian nests. In many nest predation experiments with artificial nests, plasticine eggs are used beside real eggs in order to better identify nest predators based on their tooth or beak marks (Keyser et al. Citation1998, Batáry & Báldi Citation2004, Trnka et al. Citation2008). However, the use of plasticine eggs in ground nests is often heavily criticised, because the unnatural smell of plasticine is thought to affect the results by attracting nest predators searching using olfactory cues (Rangen et al. Citation2000, Maier & DeGraaf Citation2001). Plasticine eggs are usually aired to reduce or eliminate the smell of plasticine. Another method is to use painted or varnished coated clay eggs (Boulton & Cassey Citation2006). Haskell Citation(1999) coated plasticine eggs with a thin layer of rubber (PlastiDip®), but the attractiveness of these eggs to predators was not compared with that of real eggs and the method has not been used widely.
The aim of this study was to answer the following questions. How does the survival rate of ground nests vary if we place one plasticine (P) or one coated plasticine egg (C) next to one quail (Coturnix coturnix) egg (Q) using a quail–quail egg combination as control? What is the proportion of different predator types (small or large mammal or bird) in the predation events of different nest types?
The study was carried out in the interior of intensively managed winter wheat fields in south Hungary during the 2010 summer (June–July). A total of 150 artificial ground nests were placed in ten wheat fields south of the city of Pécs. Study sites were 29.05 ± 6.72 ha (mean ± se) and the distance to the nearest study site was 0.97 ± 0.38 km (mean ± se). Every nest contained two eggs using one of the following combinations: quail + quail egg (Q + Q), quail + plasticine egg (Q + P) and quail + coated (with PlastiDip®) plasticine egg (Q + C) of the same size. Artificial eggs were aired for at least two weeks before the beginning of the study (Bayne & Hobson Citation1997, Purger et al. Citation2004). Coated plasticine eggs remained flexible after drying, so they were able to conserve the marks of predators. In each experimental plot (30 m from the edge) there were 15 nests along three linear transects (transects were 20 m apart), with randomly placed egg combinations (5 Q + Q, 5 Q + P, 5 Q + C, hereafter nest type), 20 m apart from each other. Quails (Coturnix coturnix), typical ground-nesting birds of the area, were present in the study sites throughout the experiment. Their average incubation period is 14–16 days, so we monitored nests over two weeks, on days 1, 5, 9 and 13 after their initial placement, at the same time (in the mornings). A nest was considered to be depredated if one of the eggs was damaged or missing. For identification of nest predators we used the marks left on the eggs. Nest locations within transects were marked with yellow signal tape (0.5 × 20 cm), positioned at the same side and distance (2 m) from each nest (Butler & Rotella Citation1998).
We analysed the effect of nest types on daily probability of nest survival using a generalized linear mixed model (GLMM) with binomial error distribution, logit link function, and involving nest days as denominator. This method is known as Mayfield logistic regression (Hazler Citation2004, Pasinelli & Schiegg Citation2006). Nest days were rounded up to the nearest day (Hazler Citation2004). Wheat field was included in the model as a random factor. Difference between nest types, because of the significant effect in the mixed model, was further investigated using multiple comparisons of means by Tukey contrasts. Calculations were made using the lme4 (Bates et al. Citation2011) and the multcomp (version 1.2, Hothorn et al. Citation2008) packages for R 2.11.1 software (R Development Core Team Citation2010).
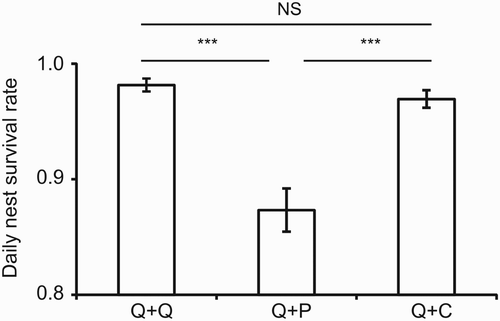
Almost half of the nests (45 %) were depredated. Analysing Mayfield nest survival rates we found that nests with quail and plasticine eggs (Q + P) suffered significantly higher predation than those containing two quail eggs (Q + Q) or those with quail and coated plasticine eggs (Q + C) (, ). The latter two combinations did not differ significantly. Furthermore, including the interaction term between nest type × site in the model did not result in a significant interaction effect (P > 0.117 for all possible nest type × site combinations), which means that there was no differential effect of treatment (nest type) according to wheat fields.
Table 1. Results of Tukey contrasts following GLMM comparing daily survival rates of artificial ground nests containing different egg types. Q, quail egg; P, plasticine egg; C, coated plasticine egg.
Figure 1. Daily nest survival rates ± se of artificial ground nests with different egg combinations.
*** Significant differences at P < 0.001. Q: quail egg; P: plasticine egg; C: coated plasticine egg.
From a total of 50 nests with two quail eggs (Q + Q), 11 (22 %) were depredated. In one case (9 %) eggs disappeared from the nest, in nine cases (82 %) eggs were broken (categorization in predator guilds was not possible), at one nest (9 %) beak marks were found on the eggs. From 50 nests containing quail and plasticine eggs (Q + P), 40 (80 %) were depredated. In two cases (5 %) eggs disappeared from the nests. We identified small mammal marks on the majority of plasticine eggs (n = 32, 80 %); a few eggs contained marks left by large mammals (n = 3, 7.5 %) and birds (n = 3, 7.5 %). Only 17 (34 %) nests with quail and coated plasticine eggs (Q + C) were depredated, mostly by small mammals (n = 10, 59 %). In four nests (23 %), quail eggs were broken and coated plasticine eggs disappeared or remained intact (hence categorization in predator guilds was not possible). In two nests (12 %), bird beak marks were found on the eggs, and in one case (6 %) eggs were taken away from the nests by unidentified predators.
Several previous studies suggested that predation rates might differ between real eggs and dummy eggs in artificial nests (e.g. Svagelj et al. Citation2003, Batáry & Báldi Citation2004, Alvarez & Galetti Citation2007). In the present study the predation rates of nests with different egg combinations differed significantly, with higher predation on nests containing plasticine eggs than those with only quail or quail with coated plasticine eggs. Based on the marks left by predators on the plasticine eggs, we found that small mammals were the main predators independently of nest type, whereas birds and large mammals were less important. It is likely that quail–quail nests were also discovered and mainly depredated by small mammals; however, identification based on marks on egg shells is more difficult. Several other studies found that the main predators of ground nests were mammals (e.g. Draycott et al. Citation2008, Padyšáková et al. Citation2010). Therefore, we think that the higher predation of nests containing plasticine eggs can be related to their smell, thereby attracting more small mammals than nests with only real (quail) and/or coated plasticine eggs. The survival rates of nests with two quail eggs and nests with quail and plasticine eggs coated with rubber layer were similar, thus we assume that the rubber layer reduced the unnatural smell of plasticine. Therefore, we recommend using real eggs with coated plasticine eggs instead of uncoated plasticine eggs, as the latter overestimate predation rates, while coated plasticine eggs still allow identification of predator types.
ACKNOWLEDGEMENTS
We would like to thank Krisztián Frank for the assistance in the field works, and Gavin Siriwardena and Gilberto Pasinelli for suggestions and corrections. We are grateful to Walter S. Svagelj and one anonymous referee, whose useful comments and linguistic corrections highly improved the manuscript. Péter Batáry was a Bolyai Research Fellow of the Hungarian Academy of Sciences.
REFERENCES
- Alvarez , A. D. and Galetti , M. 2007 . Artificial nest predation in Atlantic Forest Island: testing the place and the different types of egg . Rev. Bras. Zool. , 24 : 1011 – 1016 .
- Bayne , E. M. and Hobson , K. A. 1997 . Comparing the effects of landscape fragmentation by forestry and agriculture on predation of artificial nests . Conserv. Biol. , 11 : 1418 – 1429 .
- Batáry , P. and Báldi , A. 2004 . Evidence of an edge effect on avian nest success: a meta-analysis . Conserv. Biol. , 18 : 389 – 400 .
- Bates , D. , Maechler , M. and Bolker , B. 2011 . “ lme4: Linear mixed-effects models using S4 classes ” . R package version 0.999375-39. URL: http://CRAN.R-project.org/package=lme4
- Boulton , R. L. and Cassey , P. 2006 . An inexpensive method for identifying predators of passerine nests using tethered artificial eggs . New Zeal. J. Ecol. , 30 : 377 – 385 .
- Butler , M. A. and Rotella , J. J. 1998 . Validity of using artificial nests to assess duck-nest success . J. Wildlife Manage. , 62 : 163 – 171 .
- Draycott , R. A.H. , Hoodless , A. N. , Woodburn , M. I.A. and Sage , R. B. 2008 . Nest predation of Common Pheasants Phasianus colchicus . Ibis , 150 : 37 – 44 .
- Haskell , D. G. 1999 . The effect of predation on begging-call evolution in nestling wood warblers . Anim. Behav. , 57 : 893 – 901 .
- Hazler , K. R. 2004 . Mayfield logistic regression: a practical approach for analysis of nest survival . Auk , 121 : 707 – 716 .
- Hothorn , T. , Bretz , F. and Westfall , P. 2008 . Simultaneous inference in general parametric models . Biometrical J , 50 : 346 – 363 .
- Keyser , A. J. , Hill , G. E. and Soehren , E. C. 1998 . Effects of forest fragment size, nest density, and proximity to edge on the risk of predation to ground-nesting passerine birds . Conserv. Biol. , 12 : 986 – 994 .
- Maier , T. J. and DeGraaf , R. M. 2001 . Differences in depredation by small predators limit the use of plasticine and zebra finch eggs in artificial-nest studies . Condor , 103 : 180 – 183 .
- Padyšáková , E. , Šálek , M. , Poledník , L. , Sedláček , F. and Albrecht , T. 2010 . Predation on simulated duck nests in relation to nest density and landscape structure . Wildlife Res , 37 : 597 – 603 .
- Pasinelli , G. and Schiegg , K. 2006 . Fragmentation within and between wetland reserves: the importance of spatial scales for nest predation in reed buntings . Ecography , 29 : 721 – 732 .
- Purger , J. J. , Mészáros , L. A. and Purger , D. 2004 . Ground nesting in recultivated forest habitats – a study with artificial nests . Acta Ornithol , 39 : 141 – 145 .
- R Development Core Team . 2010 . “ R: a language and environment for statistical computing ” . Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0, URL: http://www.R-project.org
- Rangen , S. A. , Clark , R. G. and Hobson , K. A. 2000 . Visual and olfactory attributes of artificial nests . Auk , 117 : 136 – 146 .
- Svagelj , W. S. , Mermoz , M. E. and Fernandez , G. J. 2003 . Effect of egg type on the estimation of nest predation in passerines . J. Field Ornithol. , 74 : 243 – 249 .
- Trnka , A. , Prokop , P. and Batáry , P. 2008 . Dummy birds in artificial nest studies: an experiment with Red-backed Shrike Lanius collurio . Bird Study , 55 : 329 – 331 .