Abstract
Capsule The majority of female Ring Ouzels made two successful breeding attempts in a single season, but triple brooding was much less common.
Season-long reproductive success may be more dependent on the number of broods than on clutch size or the timing of breeding. Here we report on the frequency of multiple brooding in Ring Ouzels (Turdus torquatus), including the first documented cases of triple brooding in this species. Sixty-one out of 108 (56.5%) individually marked females made two successful breeding attempts in one or more years, but only two (1.8%) made three successful breeding attempts in one season, during 1999–2011. These triple-brooded females began breeding earlier in the season, and had shorter inter-brood intervals than other females. The timing of moult in preparation for migration may constrain the reproductive window at the end of the breeding period for long-distance migrants, meaning that such a reproductive strategy is uncommon for most female Ring Ouzels.
In birds, single brooding is common when breeding conditions remain suitable for a short time compared with the potential length of the breeding period (Martin Citation1987). However, if breeding conditions remain suitable for longer periods, species may become multi-brooded. In such cases, annual reproductive success may be dependent on the number of broods rather than on clutch size or the timing of breeding (e.g. Davies Citation1992, Holmes et al. Citation1992, Weggler Citation2006). However, direct measurement of the number of reproductive attempts that individuals make within a season requires continuous monitoring of a sample of marked animals. Accurate data on individual breeding frequency are therefore available from only a relatively few intensively studied species within restricted study areas (e.g. Browne & Aebischer Citation2004, Weggler Citation2006, Sim et al. Citation2011), although a method to estimate the annual number of reproductive attempts without using data from marked individuals has recently been developed (Cornulier et al. Citation2009). Thus, quantifying the number of breeding attempts made by marked individuals is important in understanding how breeding frequency influences annual reproductive success.
Double brooding (two broods reared in a single year) has been widely recorded among passerine bird species (e.g. Holmes et al. Citation1992, Smith & Marquiss Citation1995, Evans Ogden & Stutchbury Citation1996). However, triple brooding (three broods reared in a single year) is much less common (e.g. Pogue & Carter Citation1995, Friesen et al. Citation2001, Weggler Citation2006). Here we report on the frequency of multiple brooding, including two cases of triple brooding, in Ring Ouzels (Turdus torquatus), a medium-sized thrush breeding in upland areas of the UK and wintering in southern Spain and Morocco (Gibbons et al. Citation1993, Wernham et al. Citation2002). It is a red-listed species of high conservation concern in the UK due to recent population declines (Eaton et al. Citation2011), and recent research has focussed on understanding the underlying causes of these declines (Beale et al. Citation2006, Sim et al. Citation2007, Citation2010, 2011).
We studied Ring Ouzel breeding ecology in a population inhabiting approximately 18 km2 (9 × 2 km) of the southern part of Glen Clunie (56°56′N 3°25′W), in northeast Scotland, during 1998–2011. We aimed to locate all breeding pairs and nesting attempts annually by systematically walking through the study area, covering all ground to within 200 m, every 1–2 weeks between mid-April and mid-July, when breeding ceases. In each year, only one or two broods fledged from nests before we could find them; thus >90% of successful nests were located. Nest locations were recorded to the nearest 10 m using a Garmin GPS 12. British Ring Ouzels regularly make two breeding attempts per season, with each attempt taking on average 29–30 days from first egg laying (4–5 days egg laying, 13 days incubation per egg, beginning with the penultimate egg, and 13 days from hatching to fledging; Burfield Citation2002). Laying dates were recorded directly from nests found during laying or hatching, or back-calculated using known relationships between nestling age and wing length (Burfield Citation2002), or by comparison with photographs of known-age nestlings. We assumed that all breeding attempts in which egg-laying began before 11 May (the earliest known laying date for a second breeding attempt) were first attempts.
As part of an experiment to test if poor food supply for nestlings reduces body condition at fledging and thus subsequent post-fledging survival, Mealworms (Tenebrio molitor) were provided to 11 pairs in 2011, with a further 13 control pairs being left unfed. However, supplementary feeding did not improve nestling body condition at fledging, nor did it increase the percentage of second breeding attempts in fed pairs (54.5%) compared with control pairs (69.2%; unpubl. data).
During 1999–2011, 263 adult Ring Ouzels were marked with British Trust for Ornithology metal rings and three or four colour-rings to allow individual identification without need for recapture. These comprised 27 males and 27 females originally ringed as nestlings within or outside Glen Clunie, and 95 male and 114 female previously unringed individuals that were caught and ringed as breeding adults in Glen Clunie (Sim & Rebecca Citation2003). These adults were aged as 2nd year (hatched the previous year) or 2+ years (hatched two or more years previously) at capture, based on the shape of the tail feathers and contrast in colour of the greater coverts (Svensson Citation1992).
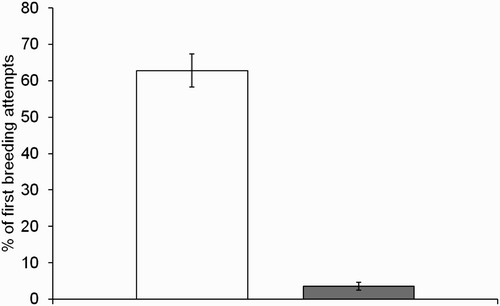
During 1999–2011, 108 of the marked females were recorded making 151 first breeding attempts (some females bred in multiple years and 33 caught at second breeding attempts were not seen in later years; ). Considering all 151 first breeding attempts, a mean of 62.8 ± 4.5% of females made second attempts (), and 45.0 ± 4.9% were double-brooded in a single year. A mean of 3.5 ± 1.1% of females (all 2+ years old) made third attempts (), and 2 (1.3%) of these successfully reared three broods to fledging in the same year. Considering only the 108 marked females, 72 (66.7%) made second attempts and 61 (56.5%) were double-brooded in one or more years. Only 6 (5.5%; all 2+ years old), made third attempts, and 2 (1.8%) of these successfully reared three broods to fledging in the same year.
Table 1. The number of occasions that 108 individually marked females were recorded making first, second and third breeding attempts in a season, the percentage of first breeding attempts that successfully reared a brood to fledging (S), failed at the egg (Fe) and nestling (Fn) stages, and the outcomes of triple breeding attempts, during 1999–2011. Only females in 2005 and 2011 successfully reared three broods to fledging within a single year.
Figure 1. The percentage (± 1 SE) of 151 first breeding attempts made by 108 individually marked females that resulted in second (unfilled bars) and third (filled bars) breeding attempts, during 1999–2011.
The first triple-brooded female was marked as a 2+ year old at her second breeding attempt in 2004, rearing a brood of 4. In 2005, she fledged broods of 4, 5 and 4 at her first, second and third breeding attempts, respectively, within 370 m of the 2004 nest. Estimated first-egg laying dates were 8 April, 11 May and 16 June, respectively. Distances between successive breeding attempts were 75 m and 87 m, and she was mated to the same marked male for all three attempts. In 2006, she moved 1.2 km to a new breeding site where she fledged a brood of 4 from her first breeding attempt. However, she was not seen again in 2006 or in later years.
The second triple-brooded female was marked as a 2nd year bird at her first breeding attempt in 2008, and fledged first and second broods of 3 and 4 young, respectively. In 2009, she fledged first and second broods of 4 and 5, respectively, within 250 m of the 2008 nest sites. In 2010, she moved 3.6 km to a new breeding site where she fledged a first brood of four young, but was not recorded making a second breeding attempt. In 2011, she fledged broods of 4, 5 and an unknown number of young from her first, second and third breeding attempts, respectively, within 230 m of the 2010 nest. The third nest was not found, but the same female was observed feeding at least three recently fledged juveniles within 100 m of the first two nests, and one of these was caught and measured (allowing estimated age and hence laying date to be calculated). Estimated first-egg laying dates were 18 April, 19 May and 19 June, respectively. The distance between first and second nests was 177 m, and she was mated to the same marked male for all three attempts.
We believe that these are the first two proven cases of triple-brooding by Ring Ouzels. Elsewhere, female Ring Ouzels have occasionally been reported to make three breeding attempts in one year, although none were individually marked or reared three broods (Appleyard Citation1994; Burfield Citation2002), and one marked pair made five unsuccessful breeding attempts in the same year (Smith Citation2003). With fewer than 2% of all marked females breeding in Glen Clunie successfully rearing three broods in one year, triple brooding is clearly uncommon in this species. In many birds the end of the breeding season is marked strikingly by reduced reproductive output and/or high mortality costs of late breeding for parents (e.g. Smith & Marquiss Citation1995, Verboven & Visser Citation1998), with unfavourable environmental conditions ultimately terminating the breeding season. However, for long-distance migrants, the timing of moult in preparation for migration may constrain the reproductive window, even if environmental conditions remain suitable (Evans Ogden & Stutchbury Citation1996, Dawson Citation2004). This appears to be the case for Ring Ouzels in Glen Clunie, where productivity was only marginally lower for second broods (Sim et al. Citation2011) and a suitable food supply was available until at least September (unpubl. data). The need to complete the post-breeding moult (Svensson Citation1992) in preparation for migration is likely to be a major constraint limiting an extended breeding season in Ring Ouzels (Porkert & Miloslav Citationin press).
The two triple-brooded females were estimated to have laid the first eggs of their first breeding attempts on 8 April (the earliest recorded laying date during our study) and 18 April, respectively. These dates are significantly earlier than the mean laying date (27 April ± 0.36 SE, n = 294) of all assumed first breeding attempts during 1998–2011. However, there was no evidence that mean laying dates of first breeding attempts showed a significant linear change across years (laying date = –0.09 × year, r = 0.15, P = 0.60, N = 13 years). In addition, the percentage of females making second breeding attempts within a year showed no significant linear change across years (percentage of females making second breeding attempts = 0.16 × year, r = 0.04, P = 0.90, N = 13 years). There was thus no apparent advancement of laying dates, or an increase in the percentage of females making second breeding attempts, and therefore no evidence that climate change was having a significant effect on these aspects of Ring Ouzels breeding ecology during our study.
The triple-brooded females had intervals of 33 and 36 days, and 31 and 31 days, between successive breeding attempts, respectively. Three of these inter-brood intervals are significantly shorter than the mean interval between first-egg laying date in first and second breeding attempts (36.4 days ± 0.45 SE, n = 87) for all marked females. The particularly short inter-brood intervals of the triple-brooded female in 2011 may have been at least partly due to the supplementary food provided to this pair during their first and second breeding attempts, as part of a larger supplementary feeding experiment. However, there was no evidence that the provision of supplementary food increased the rate of double brooding in fed pairs (see above).
These results show that triple brooding by Ring Ouzels is possible if females begin egg-laying early in the season, and re-nest rapidly. However, given the scarcity of triple brooding in this species, such a reproductive strategy is probably selected against. In particular, breeding three times in a season may reduce the probability of surviving to breed in subsequent years, and thus impact upon lifetime reproductive success (Newton Citation1989). Although the impact of triple brooding on population growth rate (λ) in our study population was low (Sim et al. Citation2011), the potential impact on λ of triple brooding in Ring Ouzels and other multi-brooded species is of high priority. Such studies should examine the number of breeding attempts made, and how this impacts upon season-long reproductive success and λ.
Acknowledgements
We thank Invercauld Estate for cooperation with access to Glen Clunie. Jerry Wilson, Staffan Roos and Jane Reid provided valuable comments on an earlier version of this Short Article. This study was funded by RSPB, the Scottish Ornithologists Club (to IMWS 2000-05 and 2010), Scottish Natural Heritage (2006-08) and the Cairngorms National Park Authority (2007).
References
- Appleyard . 1994 . Ring Ouzels of the Yorkshire Dales , Leeds : W.S. Maney & Son Ltd .
- Beale , C. M. , Burfield , I. J. , Sim , I. M.W. , Rebecca , G. W. , Pearce-Higgins , J. W. and Grant , M. C. 2006 . Climate change may account for the decline in British Ring Ouzels Turdus torquatus . J. Anim. Ecol. , 75 : 826 – 835 .
- Browne , S. J. and Aebischer , N. J. 2004 . Temporal changes in the breeding ecology of European turtle doves Streptopelia turtur in Britain, and implications for conservation . Ibis , 146 : 125 – 137 .
- Burfield , I. J. 2002 . “ The breeding ecology and conservation of the Ring Ouzel Turdus torquatus in Britain ” . In PhD dissertation , University of Cambridge .
- Cornulier , T. , Elston , D. A. , Arcese , P. , Benton , T. G. , Douglas , D. J.T. , Lambin , X. , Reid , J. , Robinson , R. A. and Sutherland , W. J. 2009 . Estimating the annual number of breeding attempts from breeding dates using mixture models . Ecol. Lett. , 12 : 1184 – 1193 .
- Davies , N. 1992 . Dunnock Behaviour and Social Evolution , New York : Oxford University Press .
- Dawson , A. 2004 . Control of the annual cycle in birds: endocrine constraints and plasticity in response to ecological variability . Phil. Trans. R. Soc. B , 363 : 1621 – 1633 .
- Eaton, M.A., Balmer, D.E., Cuthbert, R., Grice, P.V., Hall, J., Hearn, R.D., Holt, C.A., Musgrove, A.J., Noble, D.G., Parsons, M., Risley, K., Stroud, D.A. & Wotton, S. 2011. The state of the UK's birds 2011. RSPB, BTO, WWT, CCW, JNCC, NE, NIEA, and SNH, Sandy, Bedfordshire.
- Evans Ogden , L. J. and Stutchbury , B. J.M. 1996 . “ Constraints on double brooding in a neotropical migrant, the ” . Vol. 98 , 736 – 744 . Condor : Hooded Warbler .
- Friesen , L. E. , Cadman , M. D. and Allen , M. L. 2001 . “ Triple brooding by Southern Ontario Wood Thrushes ” . In Wilson Bull Vol. 113 , 237 – 239 .
- Gibbons , D. W. , Reid , J. B. and Chapman , R. A. 1993 . The New Atlas of Breeding Birds in Britain and Ireland: 1988–1991 , London : T. and A.D. Poyser .
- Holmes , R. T. , Sherry , T. W. , Marra , P. P. and Petit , K. E. 1992 . Multiple brooding and productivity of a neotropical migrant, the Black-throated Blue Warbler (Dendroica caerulescens) . in an unfragmented temperate forest. Auk , 109 : 321 – 333 .
- Martin , T. E. 1987 . Food as a limit on breeding birds: a life-history perspective . Ann. Rev. Ecol. Syst. , 18 : 619 – 636 .
- Newton , I. 1989 . Lifetime reproduction in birds , Edited by: Newton , I. London : Academic Press .
- Pogue , D. W. and Carter , W. A. 1995 . Breeding biology of secondary cavity-nesting birds in . Oklahoma. Southwest. Natur. , 40 : 167 – 173 .
- Porkert , J. and Miloslav , H. in press . A record of breeding-moulting overlap in the Common Redstart Phoenicurus phoenicurus , Bird Study . DOI: 10.1080/00063657.2012.683987
- Sim , I. M.W. and Rebecca , G. W. 2003 . “ Catching methods and biometrics of breeding Ring ouzels Turdus torquatus torquatus in northeast Scotland. Ringing & Migration ” . Vol. 21 , 163 – 168 .
- Sim , I. M.W. , Burfield , I. J. , Grant , M. C. , Pearce-Higgins , J. W. and Brooke , M.de L. 2007 . The role of habitat composition in determining breeding site occupancy in a declining Ring Ouzel Turdus torquatus population . Ibis , 149 : 374 – 385 .
- Sim , I. M.W. , Rollie , C. , Arthur , D. , Benn , S. , Booker , H. , Fairbrother , V. , Green , M. , Hutchinson , K. , Ludwig , S. , Nicoll , M. , Poxton , I. , Rebecca , G. , Smith , L. , Stanbury , A. and Wilson , P. 2010 . The decline of the Ring Ouzel in . Britain. Br. Birds , 103 : 229 – 239 .
- Sim , I. M.W. , Rebecca , G. W. , Ludwig , S. C. , Grant , M. C. and Reid , J. M. 2011 . Characterizing demographic variation and contributions to population growth rate in a declining population . J. Anim. Ecol. , 80 : 159 – 170 .
- Smith , R. D. and Marquiss , M. 1995 . Production and costs of nesting attempts in Snow Buntings Plectrophenax nivalis: why do they attempt second broods? . Ibis , 137 : 469 – 476 .
- Smith , L. 2003 . Report of the Long Mynd Breeding Project: Ring Ouzel 2004 . http://www.naturalshropshire.org.uk/LinkClick.aspx?filetic-ket=Cw6fMvUVP-8%3D&tabid=57&mid=395
- Svensson , L. 1992 . Identification Guide to European Passerines , Stockholm : L. Svensson .
- Verboven , N. and Visser , M. 1998 . Seasonal variation in local recruitment of Great Tits: the importance of being early . Oikos , 81 : 511 – 524 .
- Weggler , M. 2006 . Constraints on, and determinants of, the annual number of breeding attempts in the multi-brooded Black Redstart Phoenicurus ochruros . Ibis , 148 : 273 – 284 .
- Wernham , C. V. , Toms , M. P. , Marchant , J. H. , Clark , J. A. , Siriwardena , G. M. and Baillie , S. R . 2002 . The Migration Atlas: Movements of the Birds of Britain & Ireland , Edited by: Wernham , C. V. , Toms , M. P. , Marchant , J. H. , Clark , J. A. , Siriwardena , G. M. and Baillie , S. R . London : T. & A.D. Poyser .