Abstract
Capsule Arrival fuel load of migratory passerine birds after the Sahara crossing in spring varied between oases of different habitat quality.
Aims To test: Equation(1) if spring migrants reaching northern Africa with low fuel stopover in oases of species-specific suitable habitat; Equation(2)
whether migrants have sufficient fuel loads to reach southern Europe; and Equation(3)
if there is local variation.
Methods We collected data from passerines caught at three different oases in southeast Morocco during the spring of 2011.
Results In species typical of mesophilic vegetation in Europe, birds carrying lower fuel loads occurred at a stopover site with lusher vegetation than two oases located further south. By contrast, in species adapted to xerophilic vegetation, birds carrying high fuel loads were found in drier habitat.
Conclusion When arriving in northern Africa, migrants stopover in oases of different habitat quality in a species-specific fashion. Fuel load is, except for a few Mediterranean species, sufficiently high to reach the south of Europe without needing to refuel. Significant differences between oases suggests that fuel loads of birds from a single site are non-representative of the entire region.
Crossing geographic barriers such as oceans, mountain ranges or deserts is one of the main challenges which migrant birds must face, because fuelling in these areas is impossible or nearly impossible (Moreau Citation1972, Newton Citation2008). Migrants crossing such barriers have developed particular migratory strategies to cope with such challenges, commonly consisting of the accumulation of large fuel loads prior to these crossings (Fry et al. Citation1968, Citation1970, Hjort et al. Citation1996, Rubolini et al. Citation2002, Delingat et al. Citation2008, Fransson et al. Citation2008). The identification of target pre-migratory fuelling areas is thus of great importance from ecological, evolutionary and conservation perspectives (Ottosson et al. Citation2005, Julliard et al. Citation2006) as is the identification of those areas where migrants can refuel after crossing these barriers (Moore & Kerlinger Citation1987, Maitav & Izhaki Citation1994, Pilastro & Spina Citation1997, Maggini & Bairlein Citation2011). To understand the importance of these latter areas as stopover sites for migrants, two important issues have to be considered: firstly, how much fuel migrants have on arrival at the site and, secondly, how flexible migrants are in deciding whether or not to land at sites of species-specific suitable habitat.
The Sahara desert is one of the main eco-geographic barriers in the world and it occupies a remarkable position within the Afro-European system of bird migrations (Moreau Citation1972, Biebach Citation1990). When crossing this desert, passerines show an intermittent strategy of flying by night and resting during the day (Schmaljohann et al. Citation2007). Most European–Afrotropical passerines accumulate large fuel loads before the desert crossing, both in autumn (Izhaki & Maitav Citation1998, Schaub & Jenni Citation2000, Rubolini et al. Citation2002, Fransson et al. Citation2008) and spring (Fry et al. Citation1968, Citation1970, Bairlein Citation1991, Wood Citation1992). In spring, we do not know to what extent these fuel loads are high enough to reach the south of Europe without needing to refuel in northern Africa. Thus, while some studies carried out on several Mediterranean islands have suggested that Afrotropical migrants arrive in northern Africa carrying sufficient fuel loads to allow them to continue their migration north, without further fuelling until reaching favourable habitats in Europe (Pilastro & Spina Citation1997), other studies carried out in the first vegetated areas in northern Africa do not support this strategy (Ash Citation1969, Smith Citation1979, Gargallo et al. Citation2011, Maggini & Bairlein Citation2011). Furthermore, individuals of several bird species were reported to depart from an unfavourable site at one oasis north of the desert without enough fuel to reach the south of Europe, suggesting that they needed to stopover at favourable sites further north in order to refuel to before crossing the Mediterranean (Maggini & Bairlein Citation2011). By contrast, data from a nearby oasis have shown that migrants stopover for long enough to accumulate fuel (Gargallo et al. Citation2011). Such differences between studies carried out at oases in close proximity to each other make it hard to draw conclusions on the strategy of migrants that need to refuel in northern Africa.
If migrants with low fuel are forced to land at the first available areas, irrespective of habitat quality, while migrants in better condition are able to look for more species-specific suitable stopover sites (Yosef et al. Citation2006), it can be predicted that birds with low fuel loads will be proportionately more abundant at less suitable than at more suitable stopover sites (oases). However, studies carried out in the middle of the desert showed that migrants at small oases had more fuel than at large oases (Bairlein Citation1992, Salewski et al. Citation2010). This result was attributed to the fact that lean migrants were forced to look for stopover sites with better fuelling opportunities. If this rule is also true for sites close to the desert border in northern Africa, then we expect a higher proportion of migrants with large fuel loads in oases of habitat that might be considered as less suitable for them.
Another relevant factor to be considered for both predictions is that not all species follow similar fuelling strategies: some species seem to acquire most or all of the fuel required to reach Europe south of the Sahara, while others gain fuel both south and north of the desert (Ottosson et al. Citation2002, Ottosson et al. Citation2005, Gargallo et al. Citation2011). For migrants using the first strategy, birds in poorer condition should be expected to select more optimal sites when they reach northern Africa, when compared with conspecifics in better condition, which could be less selective than lean birds. Such birds are not forced to refuel, so they can just rest in the shade of any vegetation and wait for nightfall (Bairlein Citation1992, Jenni-Eiermann et al. Citation2011). By contrast, in migrants exhibiting the second strategy – staged refuelling, where all birds would still need to refuel to some degree – fuel load is expected to make less difference in habitat choice. This is because of the fact that, while lean birds need better areas, birds in good condition will also need to search for and refuel in these areas.
The main goal of the present study was to test: Equation(1) if migrants reaching northern Africa with low fuel loads tend to stopover mainly in oases of less or more species-specific suitable habitat, as well as whether such strategy differs across species; Equation(2)
to what extent European–Afrotropical migrants reaching the first oases in northern Africa after the Sahara have sufficient fuel loads to reach southern Europe; and Equation(3)
whether there is local variation (i.e. differences among oases), preventing conclusions from one site being extrapolated to the entire region. To test this, we collected data from captured birds at three different oases in southeast Morocco during the spring migration period of 2011.
METHODS
Sampling area and protocol
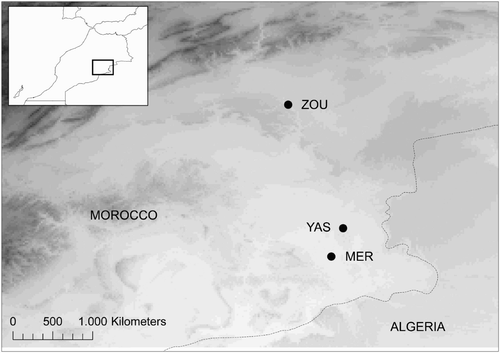
This study was carried out at three oases situated in southeast Morocco. These were (from north to south): Zouala (31°47′N 04°14′W, 932 m asl), Yasmina (31°13′N 03°59′W, 738 m asl), and Merzouga (31°05′N 04°02′W, 700 m asl) (). The main vegetation in all three sampling sites was tamarisk (Tamarix spp.). More specifically, Zouala consisted of a riparian forest of tamarisk situated within a large oasis with abundant Date Palm (Phoenix dactylifera) and orchards; Yasmina consisted of a small patch of tamarisks in a still water lake (which was dry in 2011), situated next to a dune area (sand desert); Merzouga consisted of quite extensive tamarisk woodland with a substratum of herbaceous plants (Hyoscyamus spp.) sited around the shores of a reservoir which was also dry in 2011. Thus, for species adapted to live in mesophilic (Eurosiberian) vegetation (for details see ), we considered Zouala as a ‘good’, more suitable stopover, Yasmina as a ‘poor’ stopover, and Merzouga (with a habitat very similar to Yasmina, but denser and spread over a larger area) as an ‘intermediate’ stopover. By contrast, for species adapted to xerophilic (Mediterranean) vegetation, Zouala is more likely to constitute a poor, less suitable stopover site, while Merzouga and, to a lesser extent, Yasmina, constitute good stopover sites.
Table 1. Number of captures and within-season recaptures (each bird counted only once) of the eight most abundant European–Afrotropical passerines at three oases in southeast Morocco during the spring migration in 2011.
Figure 1. Locations of the three oases where the study was carried out.
MER, Merzouga; YAS, Yasmina; ZOU, Zouala; altitudinal ranges in the area are represented in grey (higher areas shown in darker grey).
The sampling period lasted from 9 to 24 April 2011. European–Afrotropical migrants were caught with mist-nets (16 mm mesh; 2.5 m high), which were usually open from dawn to dusk, but closed on some days when temperatures were high during the midday and some afternoons and also in the presence of strong wind or rain storms. We placed 108–204 linear metres of mist-nets in Zouala, 213 m in Yasmina, and 84–180 m in Merzouga.
Once caught, each bird was ringed and its age and sex were determined when possible (Svensson Citation1996). We did not consider age and sex as additional factors for further analyses (i.e. the analysis was carried out purely at the species level) because many species undergo a complete moult while still in their wintering areas in tropical Africa (Svensson Citation1996), thus making it impossible to separate first-year birds from adults and because the sample sizes were small in some species. For each bird we also measured P3 (length of the third primary feather, numbered ascendently) with 0.5 mm accuracy and body mass with 0.1 g accuracy. No bird was kept for longer than an hour.
Statistical analyses
To examine whether the bird assemblage of European–Afrotropical migrants differed among oases we conducted a hierarchical analysis of clusters using Morisita's index of similarity and the Unweighed Pair Group Method (UPGM) algorithm. We used the Morisita's index since it is more robust than others in cases of unbalanced sample size, as well as more biased towards dominant taxa (Magurran & McGill Citation2011). This bias was irrelevant for our purposes since we were only interested in the most abundant species. Additionally, we calculated the H′ (Shannon) index of diversity and conducted paired t-tests in order to compare diversity indices between oases.
To test how the proportion of recaptures differed in relation to species and site, we used contingency tables.
To test whether fuel load differed between species and sites we used generalized linear models (glms), with dependent variable fuel load over lean body mass (lean body mass was mass when fat and muscle score were equal to zero), with site and species as predictive factors. Fuel load, f, was calculated as a proportion of actual mass, m, over lean body mass, m i : f = [(m – m i )/m i ]. To calculate m i we used the following equation:
Where m i is lean body mass, m 0 is the mean body mass of a sample of migrants with zero fat and muscle scores calculated for the mean P3 length, P3 i is the individual P3 length, P3 0 is the mean P3 length of the birds for which the m 0 values were obtained, and B is a coefficient. Data used to calculate the parameters m 0 , P3 0 , and B were obtained from a larger data set from migrants caught at Yasmina between 2008 and 2010 (Gargallo et al. Citation2011) (for details see ).
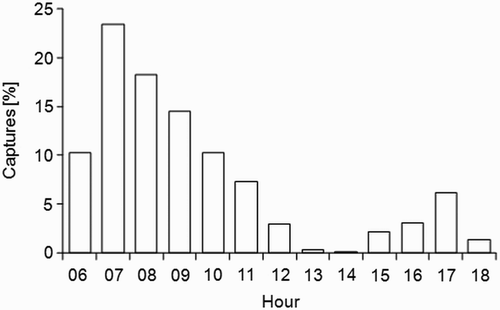
Fuel load differed among sites and species but there was an interaction between the two factors as well as an effect of catching time (anova: site, F 2 = 20.3, P < 0.001; species, F 7 = 22.7, P < 0.001; site × species, F 14 = 2.3, P = 0.004; catching hour, F 1 = 56.5, P < 0.001). Fuel load was positively correlated with catching time, although a species-by-species analysis revealed that such a correlation was significant only in five species (correlation tests not shown). To minimize such bias (e.g. one species may have variable daily patterns of capture for each site which may cause differences among sites), we repeated the correlations for the fraction of migrants caught before 10:00 (i.e. during a period of about four hours starting at dawn) and in this case the correlation was non-significant for all species (tests not shown; all r-values, P > 0.05). Overall, 66.3% of captures were made before 10:00 (). Therefore, we analyzed fuel load of first captures after dawn, which was hence considered as a proxy of arrival fuel load. This assumption is particularly useful for very small oases like Yasmina, where migrants often show very high recapture rates (Gargallo et al., unpubl. data), hence the first capture is likely to be of a bird recently landed. At larger sites, however, first captures are more likely to be made when the birds have already been present at the site for some time, since at these larger sites recapture rates are lower (Schaub et al. Citation2001). In the present study this might have been the case in Zouala or even Merzouga. Therefore, in these cases arrival fuel load may have been overestimated.
Figure 2. Hourly distribution of captures for the three study oases. Mist nets were opened from dawn to dusk.
n = 1066 (data considered here: first capture event of each individual; only birds with measured body mass and P3).
Flight ranges were calculated assuming that flying passerines lose mass at a constant rate of 1% per hour of flight (Hussel & Lambert Citation1980, Kvist et al. Citation1998). According to Delingat et al. Citation(2008), potential flight ranges Y (in km) can be assessed as:
Where U is groundspeed and f is fuel load.
Air speed of small passerines is reported to be of about 36 km h−1, i.e. 10 m s−1 (Bruderer & Boldt Citation2001). Wind profit during flight, however, can be of the same magnitude as birds' air speed. Thus, to obtain a more adjusted estimation of ground speed we calculated mean wind profit along the study period by adding it to birds' air speed. We obtained data on both wind velocity (m s−1) and direction the wing goes (degrees) from National Oceanic and Atmospheric Association (www.noaa.gov), at 925 mb, at Zouala. We calculated wind profit, b, as:
Where V is the wind velocity; αt is the (presumed) departure direction (in this case 0°), and αw is the direction of the wind (Akesson & Hedenstrom Citation2000). Thus, the groundspeed was calculated as:
Where g is the groundspeed, a is the air speed and b is the wind profit.
Two approaches were used to estimate flight ranges. First, we considered a basic starting model, assuming a continuous flight with no stopovers or different flight costs in relation to water or energy consumption during the day. Second, we considered an intermittent strategy with passerines flying at night while stopping-over during the day (Schmaljohann et al. Citation2007), so that energy is needed for the flight but also for the time on ground. Energy consumption during stopover is about 0.5% per hour (Meijer et al. Citation1994). Assuming that flights are carried out for a period of about 10 hours (from 20:00 to 06:00; Schmaljohann et al. Citation2007), stopover duration is about 14 hours. Therefore, we recalculated flight ranges assuming this intermittent strategy (Schmaljohann et al. Citation2008, Salewski et al. Citation2010). All analyses were run using the statistics packages spss 18.0 and past 1.97 (Hammer et al. Citation2001). All means are given ± se.
RESULTS
Bird assemblage and proportion of recaptures
Of the 61 species caught (), 27 (44.3%) were European–Afrotropical migrants. The mean number of captures per day (taking the nine most abundant European–Afrotropical migrant passerine species, with ≥ 10 captures at each site; we considered here both birds with and without all measurements) differed among sites (F 2,44 = 4.2, P = 0.022), with fewer captures at Yasmina (5.8 ± 1.0 captures per 100 m of mist-nets) than at the other two oases (Merzouga: 8.6 ± 1.0 captures per 100 m; Zouala: 10.8 ± 1.5 captures per 100 m, without significant differences between these two oases after a Duncan test, P > 0.05).
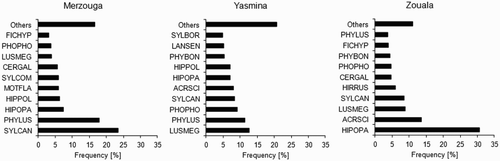
The bird assemblage of European–Afrotropical migrants differed among oases. In particular, the two oases further south were more similar to each other than to the oasis further north, where, in addition, the assemblage was more homogeneous because one species, the Olivaceous Warbler Hippolais opaca accounted for more than 30% of the abundance and thus was dominant over the rest () (H′ index ± sd: Merzouga, 2.54 ± 0.042; Yasmina, 2.74 ± 0.038; Zouala, 2.37 ± 0.048; all paired t-tests had P-values < 0.001).
Figure 3. Frequency distribution of the ten most abundant European–Afrotropical passerines captured at each of the three oases in Tafilalt, Morocco.
Each individual is counted only once; for abbreviations see Appendix 2.
Nine species overall had ≥ 10 captures at each sampling site (), representing 79.6% of the captures of European–Afrotropical passerines at the three oases and these were hence the focus of our more detailed analyses. One species' totals, however, the Nightingale (Luscinia megarhynchos), was composed of both local breeding birds and migrating ones at one of the sites (Zouala). Accordingly, this species was removed from further analyses.
The proportion of recaptured birds differed between species (χ2 = 41.3, df = 7, P < 0.001) ranging between 1.8% for Willow Warblers Phylloscopus trochilus and 16.4% for Olivaceous Warblers, but remained consistent among sites (contingency tables: all P-values > 0.05).
Fuel load
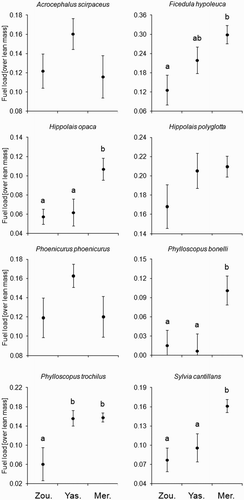
The anova of fuel load with site and species as factors revealed a significant interaction between species and site (). In five species, body mass was observed to differ among sites (), with higher values at Merzouga than at the other two oases ().
Table 2. Results from the anova used to test for the effect of site (study oasis) and species on fuel load.
Table 3. Species-specific anovas to test for differences in fuel load between sites.
Figure 4. Fuel loads (mean ± se) of eight European–Afrotropical migrant species caught at three oases in southern Morocco during spring migration in 2011.
In species where we detected significant differences among sites (), the same letters lump mean values without significant differences between them.
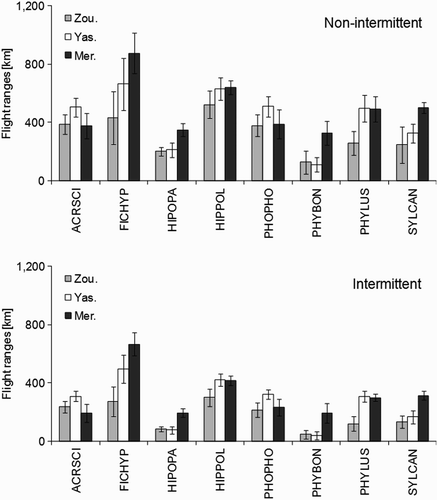
Mean wind profit along the study period was –0.46 ± 0.83 m/s, i.e. headwinds (range: –6.02 m/s to +3.63 m/s), so the groundspeed was 9.54 (i.e. 34 km/h). Potential flight ranges of migrants caught at Merzouga (the southernmost sampling site) varied from 328 ± 86 km for the Bonelli's Warbler Phylloscopus bonelli to 890 ± 140 km for the Pied Flycatcher Ficedula hypoleuca, while at Zouala (the northernmost sampling site) they varied from 54 ± 82 km for the Bonelli's Warbler to 530 ± 94 km for the Melodious Warbler Hippolais polyglotta. The majority of migrants arriving at the northern border of the Sahara had sufficient fuel to reach the coast of northern Africa and even to reach far into Iberia (). However, they did not carry sufficient fuel loads to reach the north of Iberia or the northern coast of the Mediterranean (). Assuming an intermittent flying strategy, however, flight ranges were found to be lower than assuming a non-stop flight ().
Figure 5. Potential flight ranges (mean ± se, assuming both a non-intermittent and intermittent flight strategy) of European–Afrotropical migrants caught at three oases on the northern border of the Sahara in southern Morocco during spring 2011.
Reference distances from Merzouga (in a straight line): southern Mediterranean coast, 460 km; southern coast of Spain, 630 km; western and eastern edge of the Pyrenees, 1380 and 1410 km, respectively.
DISCUSSION
We studied how the bird assemblage and fuel load of European–Afrotropical migrants caught during the spring migration period in northern Africa differed among oases of presumably different habitat quality. Although habitat quality was not tested specifically (e.g., food availability, vegetation characteristics, etc.), this seems likely (see Methods for details).
The bird assemblage was more homogeneous in Zouala than in Yasmina and Merzouga. This difference was mainly because of the proportionately higher number of captures of Olivaceous Warblers in Zouala, where tamarisk formations constitute a very similar habitat to the breeding one for this species (Cramp Citation1992). We have no evidence of Olivaceous Warblers breeding in Zouala, although this is a possibility that cannot be completely discarded. Anyhow, even if breeding, the number of local birds would be marginal compared with those still on migration to their breeding areas further north.
The Subalpine Warbler, a Mediterranean species adapted to arid bush (Shirihai et al. Citation2001), was much more abundant in Merzouga, the first oasis north of the Sahara border, and also one of the driest. Accordingly, our results partly support the hypothesis that migrants tend to land and stopover in species-specific preferred habitats (Bairlein et al. Citation1986), where they could have an advantage over other species which would tend to be less efficient at foraging in the dry environments (Jenni-Eiermann et al. Citation2011).
It has been reported that, when carrying large fuel loads, migrants in the desert are less likely to be captured because they are inactive until nightfall, when they resume their migration (Jenni-Eiermann et al. Citation2011). This potential methodological confound to our results cannot be ruled out, so it must be considered that our data are perhaps biased towards birds carrying lower fuel loads. The fuel load of five species (Pied Flycatcher and Olivaceous, Willow, Subalpine and Bonelli's Warblers) differed among sampling oases. In all cases the migrants caught in Merzouga had higher fuel loads than those caught in Zouala, as also did two species in Yasmina (Pied Flycatcher and Willow Warbler). For species adapted to mesophilic vegetation, it is arguable that Zouala is likely to be a better, more suitable stopover site than the arid oases of Yasmina and Merzouga. If this is true, our data show that for these mesophilic birds the presumably more suitable habitat was chiefly used by less-fuel-loaded migrants (certainly Pied Flycatcher, Willow Warblers; and probably also Reed Warblers, Melodious Warbler and Redstart; although in these latter cases their fuel loads did not differ among sampling sites). This agrees with the idea that leaner birds, which have greater need to refuel, tend to stopover in more species-specific suitable habitat. Birds with higher fuel load probably did not need to refuel and were, therefore, more likely to stopover at any site with good cover, since they only needed to rest, avoiding heat and/or predation, before continuing their migration on the subsequent night/nights (Biebach Citation1990). This rationale, however, is not valid for species adapted to xerophilic vegetation, such as the Subalpine Warbler, which has been documented consistently to refuel in northern Africa (Gargallo et al. Citation2011), and which was observed to have higher fuel loads in oases composed of habitat more suited to this species.
Our data also suggest that migrants at stopover sites located close to the desert border are not only fallouts unable to continue their migration, but also those that actively decide to land at those sites, dependent on their fuel loads, species-specific habitat requirements and migratory strategies (Salewski et al. Citation2010). This contrasts with what has been observed in the western Mediterranean islands, where lean birds were proportionately more abundant in low-quality stopover sites (Gargallo et al. Citation2011). This difference may be explained by the fact that migrants passing through northern Africa have, if necessary, more landing sites available to them, something that is not the case in the Mediterranean, where birds in poor condition probably have no other option than to land at the first available sites during the sea crossing, irrespective of habitat quality.
Longer flight ranges were mostly recorded from Merzouga. In some species, especially Olivaceous and Bonelli's Warblers, fuel load was not sufficiently high to reach the coast of northern Africa; in the remainder, the amount of fuel was sufficient to reach Iberia, although additional fuelling would be required to reach the breeding grounds in northern Europe. Thus, migrants caught in southeast Morocco need to refuel either in Iberia (southern Europe) or in northern Africa to reach their breeding areas in northern Europe (for similar results see Maggini & Bairlein [2011]). This result suggests that the hypothesis that migrants crossing the Mediterranean sea do not refuel in northern Africa (Pilastro & Spina Citation1997) cannot be considered as universal. On the contrary, northern Africa is likely to be used as a target refuelling area for a number of European–Afrotropical passerines, as suggested by other recent research (Gargallo et al. Citation2011).
Fuel load was only calculated for the first capture event of each bird during a period of four hours from dawn. This was done with the aim of estimating fuel load of migrants at arrival as accurately as possible (thus avoiding possible biases in fuel load estimation in cases of mass gain throughout the day). However, there are still some biases that must be considered when using this approach. In particular, a bird caught at a stopover site is likely to have already been settled in that site for some days before its first capture (Schaub et al. Citation2001), so that its fuel on that day is not necessarily the same as at arrival (especially if there is fuel accumulation). This eventuality is particularly applicable to large stopover sites, where the probability of capturing a bird even if present is normally low (Schaub et al. Citation2001, Arizaga et al. Citation2008). In our case this was especially true for Zouala and, to a lesser extent, Merzouga (both large stopover sites), but not for Yasmina. Arrival fuel load could thus have been overestimated for Zouala and, to a lesser extent, Merzouga, as compared to Yasmina.
In conclusion, the bird assemblage and fuel load at arrival of European–Afrotropical passerines in northern Africa differed among oases of presumed different habitat quality. In species adapted to mesophilic vegetation we observed a clear trend for birds carrying lower fuel loads to select a stopover site characterized by lusher vegetation and higher humidity levels than the two oases located further south. In contrast, in the Subalpine Warbler, a species adapted to xerophilic vegetation, birds carrying large fuel loads were found in the driest habitat most suited to this species. Dry habitats situated just north of the desert are more profitable for those species adapted to xerophilic vegetation, while riparian oases further north offer good refuelling opportunities to species adapted to mesophilic vegetation. Species-specific differences in stopover behaviour must be considered when evaluating the selection of a stopover site and the role of parameters such as stopover site size, habitat quality and season should also be considered.
ACKNOWLEDGEMENTS
We are very grateful to the people who assisted us during the field work, especially to J.F. Cuadrado, O. Gutiérrez, D. Mazuelas, and S. Claden. I. Maggini was funded by the Institute of Avian Research, Wilhelmshaven, Germany. We are also indebted to H. Rguibi, and the people from Maison Zouala and Hotel Yasmina, for their invaluable help. N. Richards, V. Salewski, H. Schmaljohann contributed with useful comments on a previous version of the manuscript.
REFERENCES
- Akesson , S. and Hedenstrom , A. 2000 . Wind selectivity of migratory flight departures in birds . Behav. Ecol. Sociobiol. , 47 : 140 – 144 . (doi:10.1007/s002650050004)
- Arizaga , J. , Barba , E. and Belda , E. J. 2008 . Fuel management and stopover duration of Blackcaps Sylvia atricapilla stopping over in northern Spain during autumn migration period . Bird Study , 55 : 124 – 134 . (doi:10.1080/00063650809461513)
- Ash , J. S. 1969 . Spring weights of trans-Saharan migrants in Morocco . Ibis , 111 : 1 – 10 . (doi:10.1111/j.1474-919X.1969.tb01598.x)
- Bairlein , F. 1991 . Body mass of garden warblers (Sylvia borin) on migration: a review of field data . Vogelwarte , 36 : 48 – 61 .
- Bairlein , F. 1992 . Recent prospects on trans-Saharan migration of songbirds . Ibis , 134 : 41 – 46 .
- Bairlein , F. , Leisler , B. and Winkler , H. 1986 . Morphologische Aspekte der Habitatwahl von Zugvögeln in einem SW-deutschen Rastgebiet . J. Ornithol. , 127 : 463 – 473 . (doi:10.1007/BF01640261)
- Biebach , H. 1990 . “ Strategies of trans-Saharan migrants ” . In Bird migration , Edited by: Gwinner , E. 352 – 367 . Berlin : Springer Heidelberg .
- Bruderer , B. and Boldt , A. 2001 . Flight characteristics of birds: I. Radar measurements of speeds . Ibis , 143 : 178 – 204 . (doi:10.1111/j.1474-919X.2001.tb04475.x)
- Cramp , S. 1992 . The Birds of the Western Palearctic, Vol. 6 , Oxford : Oxford University Press .
- Delingat , J. , Bairlein , F. and Hedenström , A. 2008 . Obligatory barrier crossing and adaptive fuel management in migratory birds: the case of the Atlantic crossing in Northern Wheatears (Oenanthe oenanthe) . Behav Ecol Sociobiol , 62 : 1069 – 1078 . (doi:10.1007/s00265-007-0534-8)
- Fransson , T. , Barboutis , C. , Mellroth , R. and Akriotis , T. 2008 . When and where to refuel before crossing the Sahara desert – extended stopover and migratory fuelling in first-year Garden Warblers Sylvia borin . J. Avian Biol. , 39 : 133 – 138 . (doi:10.1111/j.0908-8857.2008.04361.x)
- Fry , C. H. , Ash , J. S. and Ferguson-Lees , J. 1968 . Some aspects of pre-migratory weights of Palaearctic migrants in Nigeria . Ibis , 110 : 413
- Fry , C. H. , Ash , J. S. and Ferguson-Lees , J. 1970 . Spring weights of some Palaearctic migrants at Lake Chad . Ibis , 112 : 58 – 82 . (doi:10.1111/j.1474-919X.1970.tb00076.x)
- Gargallo , G. , Barriocanal , C. , Castany , J. , Clarabuch , O. , Escandell , R. , López , G. , Rguibi , H. , Robson , D. and Suarez , M. 2011 . Spring bird migration in the western Mediterranean: results from the Piccole Isole Project Monografies del Museu de Ciències Naturals, Vol. 6. Museu de Ciències Naturals, Barcelona
- Hammer , Ø. , Harper , D. A.T. and Ryan , P. D. 2001 . PAST: palaeontological statistics software package for education and data analysis . Palaentol. Electron. , 4 http://palaeo-electronica.org/2001_1/past/past.pdf
- Hjort , C. , Pettersson , J. , Lindström , Å. and King , M. B. 1996 . Fuel deposition and potential flight ranges of Blackcaps Sylvia atricapilla and Whitethroats Sylvia communis on spring migration in The Gambia . Ornis Svec , 6 : 137 – 144 .
- Hussel , D. J.T. and Lambert , A. B. 1980 . New estimates of weight loss in birds during nocturnal migration . Auk , 97 : 547 – 558 .
- Izhaki , I. and Maitav , A. 1998 . Blackcaps Sylvia atricapilla stopping over at the desert edge; inter- and intra-sexual differences in spring and autumn migration . Ibis , 140 : 234 – 243 . (doi:10.1111/j.1474-919X.1998.tb04384.x)
- Jenni-Eiermann , S. , Almasi , B. , Maggini , I. , Salewski , V. , Bruderer , B. , Liechti , F. and Jenni , L. 2011 . Numbers, foraging and refuelling of passerine migrants at a stopover site in the western Sahara: diverse strategies to cross a desert . J. Ornithol. , 152 ( suppl. 1 ) : S113 – S128 . (doi:10.1007/s10336-010-0572-2)
- Julliard , R. , Bargain , B. , Dubos , A. and Jiguet , F. 2006 . Identifying autumn migration routes for the globally threatened Aquatic Warbler Acrocephalus paludicola . Ibis , 148 : 735 – 743 . (doi:10.1111/j.1474-919X.2006.00573.x)
- Kvist , A. , Klaassen , M. and Lindström , Å. 1998 . Energy expenditure in relation to flight speed: what is the power of mass loss rate estimates? . J. Avian Biol. , 29 : 485 – 498 . (doi:10.2307/3677168)
- Maggini , I. and Bairlein , F. 2011 . Body condition and stopover of trans-Saharan spring migrant passerines caught at a site in southern Morocco . Ring. Migr. , 25 : 31 – 37 . (doi:10.1080/03078698.2011.586591)
- Magurran , A. E. and McGill , B. J. 2011 . Biological Diversity: Frontiers in Measurement and Assessment , Oxford : Oxford University Press .
- Maitav , A. and Izhaki , I. 1994 . Stopover and fat deposition by Blackcaps Sylvia atricapilla following spring migration over the Sahara . Ostrich , 65 : 160 – 166 . (doi:10.1080/00306525.1994.9639678)
- Meijer , T. , Möhring , F. J. and Trillmich , F. J. 1994 . Annual and daily variation and body mass of Starlings Sturnus vulgaris . J. Avian Biol. , 25 : 98 – 104 . (doi:10.2307/3677026)
- Moore , F. and Kerlinger , P. 1987 . Stopover and fat deposition by North American wood-warblers (parulinae) following spring migration over the Gulf of Mexico . Oecologia , 74 : 47 – 54 . (doi:10.1007/BF00377344)
- Moreau , R. E. 1972 . The Palaearctic–African Bird Migration Systems , London : Academic Press .
- Newton , I. 2008 . The Migration Ecology of Birds , London : Academic Press .
- Ottosson , U. , Bairlein , F. and Hjort , C. 2002 . Migration patterns of Palaearctic Acrocephalus and Sylvia warblers in north-eastern Nigeria . Vogelwarte , 41 : 249 – 262 .
- Ottosson , U. , Waldenstrom , J. , Hjort , C. and McGregor , R. 2005 . Garden Warbler Sylvia borin migration in sub-Saharan West Africa: phenology and body mass changes . Ibis , 147 : 750 – 757 . (doi:10.1111/j.1474-919X.2005.00460.x)
- Pilastro , A. and Spina , F. 1997 . Ecological and morphological correlates of residual fat reserves in passerine migrants at their spring arrival in southern Europe . J. Avian Biol. , 28 : 309 – 318 . (doi:10.2307/3676944)
- Rubolini , D. , Gardiazabal , A. , Pilastro , A. and Spina , F. 2002 . Ecological barriers shaping fuel stores in Barn Swallows Hirundo rustica following the central and western Mediterranean flyways . J. Avian Biol. , 33 : 15 – 22 . (doi:10.1034/j.1600-048X.2002.330104.x)
- Salewski , V. , Schmaljohann , H. and Liechti , F. 2010 . Spring passerine migrants stopping over in the Sahara are not fall-outs . J. Ornithol. , 151 : 371 – 378 . (doi:10.1007/s10336-009-0464-5)
- Schaub , M. and Jenni , L. 2000 . Body mass of six long-distance migrant passerine species along the autumn migration route . J. Ornithol. , 141 : 441 – 460 .
- Schaub , M. , Pradel , R. , Jenni , L. and Lebreton , J. D. 2001 . Migrating birds stop over longer than usually thought: an improved capture–recapture analysis . Ecology , 82 : 852 – 859 .
- Schmaljohann , H. , Liechti , F. and Bruderer , B. 2007 . Songbird migration across the Sahara: the non-stop hypothesis rejected! . Proc. R. Soc. B, Biol. Sci. , 274 : 735 – 739 . (doi:10.1098/rspb.2006.0011)
- Schmaljohann , H. , Bruderer , B. and Liechti , F. 2008 . Sustained bird flights occur at temperatures far beyond expected limits of water loss rates . Anim. Behav. , 76 : 1133 – 1138 . (doi:10.1016/j.anbehav.2008.05.024)
- Shirihai , H. , Gargallo , G. and Helbig , A. J. 2001 . Sylvia Warblers. Identification, Taxonomy and Phylogeny of the Genus Sylvia. Cristopher Helm, London
- Smith , G. A. 1979 . Spring weights of selected trans-Saharan migrants in north west Morocco . Ring. Migr. , 2 : 151 – 155 . (doi:10.1080/03078698.1979.9673748)
- Svensson , L. 1996 . Guía para la identificación de los paseriformes europeos , Madrid : Sociedad Española de Ornitología .
- Wood , B. 1992 . Yellow Wagtail Motacilla flava migration from West Africa to Europe: pointers towards a conservation strategy for migrants on passage . Ibis , 134 : 66 – 76 .
- Yosef , R. , Markovets , M. , Mitchell , L. and Tryjanowski , P. 2006 . Body condition as a determinant for stopover in bee-eaters (Merops apiaster) on spring migration in the Arava Valley, southern Israel . J. Arid Environ. , 64 : 401 – 411 . (doi:10.1016/j.jaridenv.2005.06.012)