Abstract
Capsule The breeding season diets of White-tailed Eagles and Golden Eagles in western Scotland were different, and there was no evidence of competition between the two species.
Aim To test the hypothesis that the reintroduced White-tailed Eagles will have an adverse effect on Golden Eagles through competition for food.
Methods Collections of prey remains at nests and regurgitated pellets during the breeding season were analysed. Temporal change in Golden Eagle territory occupancy and breeding productivity where the influence of White-tailed Eagles should be most evident was also examined.
Results Diet breadth was similar between species but diet composition was significantly different, with White-tailed Eagles taking more sheep and aquatic or coastal food items, whereas Golden Eagles took more gamebirds (Galliformes), lagomorphs and other terrestrial prey. Diet overlap comparisons, between species-pairs that nested close together or far apart, rejected a hypothesis that there was competition for food, but partially supported an alternative hypothesis that diet overlap indicated abundant shared food. There was no indication that White-tailed Eagles have had any long-term effect on the breeding productivity or abundance of territorial Golden Eagles.
Conclusions There is no evidence that reintroduced White-tailed Eagles are having an adverse effect on Golden Eagles through competition.
White-tailed Haliaeetus albicilla and Golden Eagles Aquila chrysaetos both bred in Scotland historically (Evans et al. Citation2012), until the extinction of the former in the early 20th century, principally due to human persecution (Baxter & Rintoul Citation1953, Love Citation1983). Since their reintroduction to western Scotland in 1975 numbers of White-tailed Eagles have been increasing (Green et al. Citation1996, Evans et al. Citation2003, Citation2009). Several authors have suggested that the continued spread of the White-tailed Eagle in Scotland will lead to displacement of Golden Eagles through competitive effects as coastal ranges are reclaimed (Thom Citation1986, Watson et al. Citation1992, Watson Citation1997, Halley & Gjershaug Citation1998, Halley Citation1998).
Interspecific competition arises when the interaction of two species has an adverse effect on one or both species. Competition may occur due to: (a) exploitation, when individuals from different species share the same limited resource(s) and compete with each other through ‘scramble’ or indirect competition; or (b) interference, when individuals from different species fight or the presence of one species directly denies or reduces access to resources to a second species (Pontin Citation1982).
The primary resources for raptors, over which competition is most likely to occur, are nest-sites and food (Newton Citation1979). The two eagle species currently typically nest in different habitat in Scotland (Evans et al. Citation2010). This difference reflects historical reconstructions of distribution in Scotland (Evans et al. Citation2012) and contemporary distributions elsewhere where co-existence has had a longer history and no competitive influence has been inferred (e.g. Halley Citation1998). Therefore, if competition occurs in Scotland, and reintroduced White-tailed Eagles are to displace Golden Eagles (Thom Citation1986, Watson et al. Citation1992, Watson Citation1997, Halley & Gjershaug Citation1998, Halley Citation1998), the limiting resource is most likely to be food, and White-tailed Eagles should be the superior competitor.
Halley & Gjershaug Citation(1998) showed that the Golden Eagle is the superior interference competitor over food, based on observations of behavioural interactions at carcasses in Norway. This runs counter to the postulated competitive advantage of reintroduced White-tailed Eagles over Golden Eagles. Therefore, if this postulated advantage has substance then it infers that White-tailed Eagles should be the superior ‘scramble’ competitor over shared food supplies (Halley Citation1998). Watson et al. Citation(1992) reported, from samples collected soon after White-tailed Eagles became re-established in western Scotland, that the two species utilized a similar food resource. Halley Citation(1998) has argued that the White-tailed Eagle is better able to exploit these shared food supplies by greater digestive efficiency (thereby providing an indirect competitive advantage). Moreover, Halley Citation(1998) also suggested that the shared food resource in western Scotland may be depauperate through historical over-exploitation by humans, compared to other ‘food-rich’ areas such as Norway. In other words, the arguments that underpin the prediction that reintroduced White-tailed Eagles will displace Golden Eagles include: (1) a shared food resource; (2) this food resource is very poor intrinsically; and (3) being a poor resource, sharing of it will favour reintroduced White-tailed Eagles as the superior ‘scramble’ competitor and will thereby lead to a reduction in Golden Eagle territories.
These arguments have been criticized in a study which found no consistent evidence for adverse effects of White-tailed Eagles on Golden Eagles in western Scotland (Whitfield et al. Citation2002; see also Whitfield et al. Citation2008). Evans et al. Citation(2010) have also shown that the two species currently tend to be segregated by habitat, as they were historically in Scotland (Evans et al. Citation2012) and as they are elsewhere, currently (e.g. Halley Citation1998). So, even if food types are the same between species they are likely gathered in different areas. Hence, diet similarity by food type does not necessarily reflect exploitation of the same food resource (see also Katzner et al. Citation2003). Watson et al.'s (1992) early study, inferring diet similarity, was based on prey samples from only two pairs of White-tailed Eagles. A later dietary study, based on more samples, found substantially less dietary overlap between the two species and cast doubt on the suggestion that there was interspecific competition for food (Madders & Marquiss Citation2003). However, no formal testing of the hypothesis that there is interspecific competition for food has yet been undertaken.
For two species on simple environmental gradient(s), MacArthur & Levins Citation(1967) noted that, if the niches (or utilization of resources reflected by the environmental gradient) of two species overlapped too much, one or the other species would be excluded through interspecific competition from the environment and, more importantly, selection would operate most strongly on those individuals located in the zone of overlap. Hence, if interspecific competition affects resource utilization, overlap should be reduced in the presence of the potential competitor (Alatalo et al. Citation1985).
Nilsson Citation(1984) and Korpimäki Citation(1987) both found evidence of competition in birds of prey species that prey on small mammals, as diet similarity was lower in birds which had close interspecific neighbours than in those with no near-neighbours. In Korpimäki's Citation(1987) study, individuals of both subject-species had lower breeding success with interspecific neighbours than without such neighbours, and Nilsson Citation(1984) showed reduced breeding success in the smaller of his two subject-species.
Therefore, in examining the diet of White-tailed and Golden Eagles in western Scotland, we hypothesized that: (1) if there is interspecific competition for food, then the extent of diet overlap should be less in ‘close species-pairs’ of White-tailed and Golden Eagles that breed in close proximity than in ‘distant species-pairs’ which do not breed in close proximity (prediction 1: ‘proximity’ being governed by the range limits of Golden Eagle, as the more overtly territorial species: Cramp & Simmons Citation1980, McGrady Citation1997, Halley Citation1998); and (2) that diet overlap between close species-pairs should be negatively related to geographical proximity of the nearest interspecific neighbour (prediction 2). An alternative hypothesis is that high overlap in resource use can be a reflection of the general absence of pressure to subdivide resources rather than an indication of severe competition (Menge Citation1979, Katzner et al. Citation2003), and thus a sign of high resource availability. This conversely predicted: greater overlap in diet between close-proximity species-pairs than in distant species-pairs (alternative prediction 1); and a positive relationship between interspecific diet overlap and interspecific geographical proximity within close species-pairs (alternative prediction 2).
The primary objective of the present study was to test the competing alternative predictions of these two hypotheses. In a seasonal environment, such as western Scotland, food abundance will be greater during the breeding season, when our primary tests were conducted (Methods), and so interspecific competition (and its detection) in this season may be reduced. In our study area, however, Golden Eagle breeding productivity can be very low in several pairs, with failure to lay eggs not being uncommon in many years (Whitfield et al. Citation2001, Citation2008, R. Reid unpubl. data), indicating that food is not abundant for all pairs of Golden Eagles when breeding. Moreover, it was valid to use breeding season data for our hypothesis-tests when: (1) even slight changes in food availability can adversely affect breeding Golden Eagles (Whitfield et al. Citation2001, Citation2007); (2) interspecific competition has been documented in breeding raptors that probably have access to higher food abundance (Nilsson Citation1984, Korpimäki Citation1987); (3) Watson et al. Citation(1992) found little seasonal difference for either species in the proportions of terrestrial and aquatic prey; and (4) a previous suggestion of interspecific competition has been based partly on diet overlap during the breeding season (Watson et al. Citation1992) and a presumed depauperate food supply (Halley Citation1998).
In this article we: (1) present new dietary data complementing Madders & Marquiss Citation(2003), covering a wider geographical area and including more eagle pairs than earlier studies; (2) reassess diet composition, breadth and overlap for the two species in western Scotland; and (3) consider whether these constitute evidence for competition sufficiently serious to give cause for conservation concern.
Finally, we test the competition hypothesis directly, by examining if there has been any change in Golden Eagle territory occupation or breeding productivity on the island of Mull during the period of the White-tailed Eagle re-colonization. Mull was first re-colonized by breeding White-tailed Eagles in 1981; one of the first areas colonized by reintroduced birds. Expansion resulted in nine pairs by 2006. We therefore predicted that over the period 1981–2006, under the competition hypothesis, there should have been a reduction in Golden Eagle territory occupation and/or breeding productivity at those Golden Eagle territories where increased White-tailed Eagle activity has been most pronounced.
METHODS
Our study areas were two islands of the Inner Hebrides of western Scotland (Skye and Mull) and the northernmost islands in the Outer Hebrides of western Scotland, Lewis and Harris. The period of study for diet encompassed the years 1992, 1998–2002 and 2004–2006, with most data being collected 1998–2002. Our primary datum used in analyses was the dietary information gathered at a territory of each species. Whenever possible, we tried to match temporally the collection of data on the diet of the two species. As Golden Eagles leave fewer prey remains than White-tailed Eagles (Madders & Marquiss Citation2003), we supplemented dietary data for Golden Eagles using collections from additional years whenever possible to give greater confidence through sample size. Diet of individual pairs/territories can vary across years in western Scotland (Madders & Marquiss Citation2003), but with an absence of periodically abundant temporally cyclical prey species (Steenhof & Kochert Citation1988, Steenhof et al. Citation1997) we did not consider occasional non-contemporaneous sampling a serious inherent bias in our study, as these represented less than 20% of samples and we had confidence in repeatability of interannual estimates. Data collections in more recent years (2004–2006) reflected the geographical expansion of White-tailed Eagles over time and consequent opportunities for additional interspecies comparisons.
We collected two types of dietary data: from regurgitated pellets; and from prey remains (e.g. feathers, bones, fur) gathered at nests (Madders & Marquiss Citation2003, Marquiss et al. Citation2003). Pellets were collected from April to August at and immediately around nest-sites, and at roost sites known to be used by the territorial pair in question. Pellets were attributed to eagle species on the basis of moulted feathers found simultaneously below the same perches. In the few instances where there was uncertainty, pellets were not attributed to one or the other eagle species and not included in the study.
Prey remains were collected from White-tailed Eagle nests on Mull every week during the period when eaglets were 3–9 weeks old (early May to early July), and again after they had fledged (August). In these circumstances, any large remains which still provided a meal for pre-fledged young were left in situ and logged to avoid any duplication with later collections, or, if required for analysis of lamb predation/scavenging (Marquiss et al. Citation2003), were substituted with equivalent food mass. Elsewhere, White-tailed Eagle nests were cleared of remains once each year after eaglets had fledged. The more intensive effort on Mull was due to a detailed study on predation of lambs (Marquiss et al. Citation2003). ‘End of season’ collections bias diet estimates towards items which are robust and less subject to deterioration (e.g. large bones) and against items which are subject to rapid deterioration (e.g. small mammals, soft-bodied items such as fish) (Mersmann et al. Citation1992). As our use of Mull White-tailed Eagle data was spread between ‘neighbouring’ close pairs and ‘non-neighbouring’ distant species-pairs, this bias should not have affected our hypothesis-testing. In all Golden Eagle pairs, collections of prey remains were made at nests after any young had fledged: as noted by Madders & Marquiss Citation(2003) these produced fewer prey items than comparable searches at White-tailed Eagle nests.
Prey remains were identified to species, ensuring that the collected parts of an item did not result in the same individual being counted twice. A total of 106 classes were used, encompassing birds, mammals, reptiles and amphibians, fish and cephalopod. Materials in pellets were identified down to the lowest practical taxonomic level. When necessary, feather fragments were classified after examination of the downy barbules at 200× magnification, using a reference collection and Brom Citation(1986). Mammalian hair was classified using the medullary pattern, using Teerink Citation(1991). Due to inconsistency between individual pellets in the taxonomic level at which prey could be identified, we adopted fewer classification categories than for prey remains. Moreover, dietary indices can be sensitive to misidentification of prey class (Greene & Jaksic´ Citation1983), and so we used fewer classes in analyses of regurgitated pellets than in analyses of remains, to reflect the relative difficulties in identification to the lowest taxonomic level by method. The seven classes used for pellets were: (1) deer, (2) sheep and lambs, (3) lagomorphs (i.e. rabbits and hares), (4) other mammals, (5) seabirds, (6) other birds and (7) fish. The presence of any material in a pellet which was assigned to one of the seven prey classes was considered as one item for that prey class.
Diet breadth was indexed using the formula of Levins Citation(1968):
Several formulae are available to index diet breadth (Krebs Citation1989, Margurran Citation2003). Levins' index tends to give weight towards abundant resource states (prey items, in this case) (Krebs Citation1989, Margurran Citation2003), and so was considered more appropriate when examining potential competition than, for example, the Shannon index , which tends to weight rare resources (Krebs Citation1989, Margurran Citation2003). Nevertheless, choice of index probably made little difference to our conclusions because, for example, using Golden Eagle prey remains with pairs/territories as replicates there was a high correlation between Levins' and Shannon indices (r = 0.92, n = 15).
Diet overlap was estimated using Pianka's Citation(1973) index:
Pianka's Citation(1973) index is more sensitive to sample size than, for example, Morisita's Citation(1959) index (Smith & Zaret Citation1982), but has broader utility (Krebs Citation1989, Gotelli & Entsminger Citation2006). To avoid bias associated with low sample size (Katzner et al. Citation2005, Ontiveros et al. Citation2005) we used only samples involving at least 10 prey items to estimate the diet of each pair/territory (Korpimäki Citation1987). The sample size necessary to estimate a consistent picture of diet will depend on diet breadth and, hence, also on the extent to which different lower prey taxa (e.g. species) are combined by the researcher into categories of higher taxa (e.g. genera). For prey remains, which had the higher number of prey classes (i.e. 106 species-classes, and so for which any inconsistency in diet estimation through sample size should have been most apparent), we found that with a minimum of 10 items per pair/territory (= replicate) there was no correlation between sample size and Levins' index for both White-tailed Eagle (r = –0.24, n = 16, P = 0.36) and Golden Eagle (r = 0.17, n = 18, P = 0.51). We were therefore confident that, despite the wide range in sample size per pair/territory (prey remains from a territory: White-tailed Eagle, 37–356, mean 121 (± 99 sd), n =16; Golden Eagle, 10–69, mean 31 (± 21 sd), n = 18) sample size did not influence our estimation of diet breadth and, hence, overlap.
By the same token, estimates of diet breadth and diet overlap are respectively under- and overestimated by the extent to which prey species or distinct food types are combined into groups or higher taxa (Greene & Jaksic´ Citation1983). Therefore, we preferred using diet indices derived from prey remains, with a much higher number of prey classes, to avoid predisposing our results towards the competition hypothesis. In addition, the fact that our collections of pellets were more restricted in geographical range, with most coming from the island of Mull, led us to base our dietary hypothesis-testing on data from prey remains and not from pellets. Nevertheless, although we rely on prey remains for hypothesis-testing, we also present results from pellet analyses, to provide additional comparisons, both with our results from prey remains and with previous studies which used this method in the same locale (Watson et al. Citation1992).
The nest-sites of each pair/territory of each species used in the years of study were entered in a Geographical Information System (GIS: ArcView) and the average location taken as the territory centre (McGrady et al. Citation2002, McLeod et al. Citation2002). The distance from the territory centre of each species to the nearest territory centre of the other species was then calculated. We used these distances to form ‘species-pairs’, with each species' territory centre being paired with the nearest other species' territory centre. ‘Neighbouring’ close pairs were those within 3 km of each other, and ‘non-neighbouring’ distant pairs were those more than 3 km apart. We used 3 km as a threshold because this encompasses all or most of the range use of territorial Golden Eagles in our study areas (McLeod et al. Citation2002, Haworth et al. Citation2006, R. Reid unpubl. data). Hence, in neighbouring ‘close’ pairs, each species had a nest-site of the other species in close proximity, and in non-neighbouring ‘distant’ pairs they did not. We analysed data by 3 km-distance categories (using non-parametric Mann–Whitney tests) rather than using a more complex generic geographical distance versus dietary distance matrix correlation (Mantel) test (Mantel Citation1967, Sokal & Rohlf Citation1995) because our hypotheses assumed a discontinuous influence (due to Golden Eagle territoriality) rather than a continuous influence as assumed by Mantel tests. As we did not have dietary data for all nearest neighbours, sample sizes vary between the presentation of generic results and those specifically concerned with ‘close’ or ‘distant’ species-pairs. Statistical packages used in analysis involved SPSS v.15 and R 2.4.1.
To test the competition hypothesis using data from the island of Mull, which was one of the first areas colonised by reintroduced White-tailed Eagles (in 1981) and where nine breeding pairs were present in 2006,we divided the period 1981–2006 in to two equal component periods, 1981–1993 and 1994–2006. This division categorized the increasing temporal influence of an expanding White-tailed Eagle presence. The expectation, under the competition hypothesis, was that Golden Eagle demographic metrics (occupation of territories and/or breeding productivity) should be lower in the later period, when the influence of White-tailed Eagles was greater.
Following on from the methods described by Whitfield et al. Citation(2002), White-tailed Eagle activity was classed on an annual basis as high (score = 2) if a pair nested within 3 km of a Golden Eagle territory centre (McLeod et al. Citation2002, Haworth et al. Citation2006) or low (score = 1) if a pair nested within 3–6 km distance of the territory centre of a Golden Eagle, or if there was at least one record of an immature White-tailed Eagle within the presumed Golden Eagle territory. No records of White-tailed Eagles within a Golden Eagle territory, either as breeding birds or as sightings of immatures, gave a null score (= 0) for that year. Annual White-tailed Eagle activity scores were averaged within the two periods for each of the known 32 Golden Eagle territories on Mull to derive a measure which was then contrasted across the two periods as either an increase or no change/decrease. We also documented whether average Golden Eagle breeding productivity (fledglings per year) on each territory had declined or increased across the same two periods. This gave us a 2 × 2 contingency table of White-tailed Eagle activity (increase or decrease/no change) and Golden Eagle productivity (increase or decrease/no change) with the total of all cells being 32, the total number of Golden Eagle territories on Mull. We also categorized Golden Eagle territories as having low or high overall levels of White-tailed Eagle activity depending on whether the cumulative activity score for each was higher or lower than the mean cumulative score for all Golden Eagle territories, and tested whether there was any difference in Golden Eagle breeding productivities between these two classes.
RESULTS
Median (range) diet breadth as estimated by Levins' index for prey remains was similar in both species: White-tailed Eagle, 4.81 (1.70–9.92), n = 16; Golden Eagle, 4.35 (1.14–9.10), n = 18 (Mann–Whitney U = 134, P = 0.73). Comparable statistics for Levins' indices of diet breadth using pellets, restricted to Mull due to limited and heterogeneous sampling elsewhere, were also similar between the two species: White-tailed Eagle, 2.51 (1.69–3.03), n = 9; Golden Eagle, 2.66 (1.78–3.53), n = 12 (Mann–Whitney U = 41, P = 0.59).
Similarity in Levins' indices, however, masked interspecific differences in diet composition as estimated by both methods. Diet as assessed by prey remains differed significantly between eagle species (MANOVA, F 12,21 = 7.2, P < 0.001) with Golden Eagles apparently taking fewer seabirds, waterfowl and fish, and White tailed Eagles apparently taking fewer gamebirds, ‘terrestrial’ birds (mainly passerines), lagomorphs and ‘other mammals’ (the class comprised mostly of predatory mustelids and Hedgehogs Erinaceus europaeus) (). Fish were probably underestimated by prey remains collections, as illustrated by the finding of 49 bottle corks in a White-tailed Eagle nest. The corks had been used to keep afloat Mackerel Scomber scombrus which had been thrown behind tourist boats viewing the eagles, but no remains of Mackerel were found in the nest. To avoid comparative bias this vicarious information on contribution of Mackerel to this pair's diet was ignored, but it still served to illustrate that while, from directly comparable methods, White-tailed Eagles ate fish more often than Golden Eagles, this interspecific difference was probably severely underestimated by our methods.
Table 1. Mean percentage composition of diets of Golden Eagles (n = 18) and White-tailed Eagles (n = 16) as assessed by analysis of prey remains gathered at nests. Seabirds = Procellariidae, principally Fulmar Fulmarus glacialis, and also Northern Gannet Morus bassanus, Phalacrocoracidae, Laridae, Alcidae; Waterbirds = mostly Grey Heron Ardea cinerea; Waterfowl = Anseriformes; Waders = Haematopodidae, Scolopacidae and Charadriidae; Diurnal raptor and owl = Accipitridae, Falconidae and Strigidae; Gamebirds = Red Grouse Lagopus lagopus and Pheasant Phasianus colchicus; Terrestrial birds = Passeriformes, mainly Raven Corvus corax and Hooded Crow C.cornix, and other birds not covered by other classes; Lagomorphs = Rabbit Oryctolagus cuniculus and Mountain Hare Lepus timidus; Deer and goat = Cervidae (mostly Red Deer Cervus elaphus) and Feral Goat Capra hircus; Sheep = Ovis aries mostly lambs; Other mammals = including voles, shrews, Mustelidae and Hedgehog Erinaceus europaeus; Fish = including Lesser Spotted Dogfish Scyliorhinus canicula, Lumpfish Cyclopterus lumpus, Labridae. Total items = 1937 (WTE) and 560 (GE). Asterisks denote significance level of differences between species using F tests in a MANOVA: * P < 0.05, ** P < 0.01, *** P < 0.001 (df = 1,32 in each case).
Fish were found in pellets from four of nine White-tailed Eagle pairs and none were found in any Golden Eagle pellets, but as most pellets contained no detectable fish remains, we excluded fish as an item from our pellet analysis. Diet as assessed by pellets differed significantly by eagle species (MANOVA, F 6,14 = 6.7, P = 0.002): Golden Eagle pellets contained fewer sheep (probably mostly lambs: P < 0.001), and fewer seabirds (P = 0.015) but more lagomorphs (mostly Mountain Hare Lepus timidus: P = 0.005) and more ‘other birds’ (P = 0.008) (df = 1,18 in each case) ().
Figure 1. Mean (+ se) proportions of six food classes in White-tailed Eagle (WTE, n = 9) and Golden Eagle (GE, n = 12) pellets collected April to August on the island of Mull. Total items = 1286 (WTE) and 610 (GE).
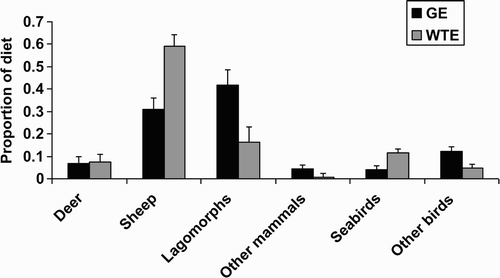
Under the test of prediction 1 of the competition hypothesis, we expected that there should be less dietary overlap in ‘close’ species-pairs which nested close together than in ‘distant’ species-pairs which did not nest close together. There was no difference in the extent of diet overlap (as estimated by Pianka's index using prey remains) between close ‘neighbouring’ pairs (n = 7) and distant ‘non-neighbouring’ pairs (n = 6) (Mann–Whitney U = 20, P = 0.87) (). Thus, prediction 1 of the competition hypothesis was rejected.
Table 2. Pianka’s Citation(1973) index of diet overlap using prey remains for ‘species-pairs’ of White-tailed Eagles (WTE) and Golden Eagles (GE) according to whether individuals of the two species nested close (< 3 km between territory centres) to each other (‘neighbours’ or close pairs) or not close to each other (> 3 km apart: ‘non-neighbours’ or distant pairs).
There was a negative correlation between interspecific territory centre distance and Pianka's index, within close neighbouring pairs (Spearman's r = –0.86, n = 7, P = 0.014). This disagreed with the test of prediction 2 of the competition hypothesis which expected that, within close neighbouring species pairs, there should be a positive correlation between territory centre distance and Pianka's index. Rather, the result pointed to support for the alternative hypothesis, that overlap in resource use reflected a general absence of selection pressure to subdivide resources.
There was no consistency in the food type which generated higher diet overlap across the two eagle species that nested in close proximity. The largest Pianka's index involved a shared heavy dependency on Fulmars Fulmarus glacialis, but in other species-pairs with high diet overlap several other prey species were involved. Hence, there was no consistent type of food that was shared by both species, even when they nested in close proximity.
There was no indication that increased White-tailed Eagle activity between the two periods, 1981–1993 and 1994–2006, had a detrimental effect on Golden Eagle breeding productivity (Fisher's Exact Test, P = 0.34), or that Golden Eagle productivity varied between ranges that experienced high or low White-tailed Eagle activity (Mann–Whitney U = 112.5, P = 0.56). The annual number of occupied Golden Eagle territories (mean ± sd) also did not differ between the two periods: 1981–1993, 27 ± 4; 1994–2006, 27 ± 3; t = 0.12, df = 24, P = 0.91).
DISCUSSION
We found no evidence of competition for food resources between the two eagle species as judged by their respective breeding season diets. Both our tests rejected the competition hypothesis on the basis of dietary overlap: the diets of the two species were substantially different. Our second test supported the alternative hypothesis, that diet overlap reflected abundant shared resources because pairs of White-tailed and Golden Eagles that nested in the closest proximity had the greatest extent of dietary overlap.
Broadly speaking, simple comparisons of diet revealed significant interspecies differences, with Golden Eagles more likely to take gamebirds (principally Red Grouse Lagopus lagopus), lagomorphs (rabbits and hares) and other non-ungulate mammals, whereas White-tailed Eagles more often fed on seabirds (principally Fulmars, but also including gulls and auks), waterfowl and waders, sheep and fish. In general, therefore, Golden Eagles tended to take prey more typical of higher terrestrial altitudes, whereas White-tailed Eagles fed more on aquatic species or low altitude species with some affinity for water. This reiterates the findings of Madders & Marquiss Citation(2003) and is not surprising given the typical nest-site and range-use habitats of the two species (Cramp & Simmons Citation1980, Whitfield et al. Citation2002, Madders & Marquiss Citation2003, Evans et al. Citation2010). In this light, the shared habit of feeding on sheep which has been emphasized previously as evidence of competition (Watson et al. Citation1992, Halley Citation1998) is probably likely also to have involved a large element of spatial (altitudinal) segregation, because sheep are found at all altitudes in the study areas (Evans et al. Citation2010). Moreover, on the occasions when we did document diet overlap in species-pairs nesting in close proximity, it did not primarily involve sheep.
Pairs of Golden Eagles nesting at low altitudes, more typical of White-tailed Eagle nest-sites (Evans et al. Citation2010), either tended to specialize on seabirds, like White-tailed Eagles, or took a broad spectrum of prey (although lagomorphs often predominated). Our results suggested that, when diet did overlap substantially, it involved situations when both species nested close together and shared abundant food resources. In general, however, we did not find the large degree of diet overlap originally reported for western Scotland (Watson et al. Citation1992), although that study involved only two pairs of White-tailed Eagles on Mull, and also had other difficulties (Madders & Marquiss Citation2003).
Biases in the methods used to study raptor diet have been the subject of considerable research (e.g. Mersmann et al. Citation1992, Redpath et al. Citation2001, Rutz Citation2003): the generic conclusion is that no method is ideal, and the extent of bias depends substantially on the predator and prey involved. As has been found previously (Wille & Kampp Citation1983, Mersmann et al. Citation1992), both methods that we used (especially pellet analysis) underestimated fish in diets. Undoubtedly this bias affected our estimates of White-tailed Eagle diet, but probably had little effect on diet estimates for Golden Eagles, judging by our own and other authors' findings (Wille & Kampp Citation1983, Willgohs Citation1984, Sulkava et al. Citation1997, McGrady Citation1997, Watson Citation1997). In the context of our study objectives, this probably served to make our conclusions conservative with regard to rejection of the competition hypothesis. Use of prey remains is also not immune to bias (e.g. Rutz Citation2003), but on balance our preference for this method to provide our main hypothesis-tests seemed amply justified, given the greater inherent resolution of prey item classification in remains, and greater difficulty in differentiation between predator-origin in pellets (see also Rutz et al. Citation2006).
Our dietary studies were restricted to the breeding season when, in a seasonal environment, food abundance may be expected to be higher, thereby reducing the prospect of competition. Nevertheless, the interspecific competition hypothesis we were testing was proposed (Watson et al. Citation1992) using data involving this season, and was based on dietary data from only two pairs of White-tailed Eagles. As we have noted earlier, there are several other reasons why our restriction of dietary studies to the breeding season should not have seriously compromised our testing of the interspecific competition hypothesis. In addition, the contribution of altitudinal habitat differences towards breeding season differences in diet (Evans et al. Citation2010) do not appear to be affected by any altitudinal shift in the two species' activity during the non-breeding season (Whitfield & McLeod Citation2007, R. Reid unpubl. data) in keeping with observations in arctic Norway in winter (Pálsdóttir Citation2012). Moreover, as both populations of our study species are resident, any cross-seasonal competitive influence of White-tailed Eagles should have been manifest in Golden Eagle breeding success and territory occupation, because even small changes in food availability for Golden Eagles in our study areas can have detrimental impacts on these important demographic parameters (Whitfield et al. Citation2001, Citation2007). We found no such effects based on surrogate measures of temporal change in White-tailed Eagle influence, however (reiterating Whitfield et al. Citation2002). Our analyses of such demographic effects covered a time span from when there were no White-tailed Eagles present to when there were several nesting pairs of White-tailed Eagles and a large non-breeding ‘floating’ population.
The premise for an expectation of adverse effects on Golden Eagles through interspecific competition following the reintroduction of White-tailed Eagles to our study area has its roots in a comparison of maps showing 19th-century distribution of each species (Love Citation1983). These have been interpreted by, for example, Watson Citation(1997) as indicative of substantial historic separation followed by Golden Eagles moving into former White-tailed Eagle locales once White-tailed Eagles had been persecuted to extinction. Based on this assumption, it has been suggested that with the return of White-tailed Eagles, Golden Eagles should revert to their former historic distribution and, hence, decline in overall abundance. This fundamental historic expectation for interspecific competition has been criticized by Whitfield et al. Citation(2002) because both species were persecuted in and before the 19th century, but Golden Eagles were intrinsically more likely to be found over a wider area, including in upland deer forests, where any large raptor would be more tolerated by land managers. Love Citation(2003) has also noted that interpretation of his historic ‘distribution’ maps (Love Citation1983) to infer competitive exclusion was unjustified and inappropriate: mapped White-tailed Eagle records involved all available nest-site records including some before the 19th century, whereas Golden Eagle records included only clutches subsequently available in museum zoological collections. Moreover, many Golden Eagle clutches were taken when White-tailed Eagles were almost extinct at the height of cumulative persecution effects. The two distribution maps presented by Love Citation(1983) were thus dissimilar both temporally and by type, with consequent implications for differential bias. In particular, some 19th-century commentators deliberately omitted Golden Eagle sites from avifaunal records in order to protect them from collectors, while consciously identifying many White-tailed Eagle sites that had been abandoned with no prospect of reoccupation (Evans et al. Citation2012).
A thorough examination of the available evidence of the historical distributions of the two species (Evans et al. Citation2012) has shown that the notion that the two species had substantially separate distributions in the past is fallacious, and that Golden Eagles were not restricted to mountainous inland areas, but also nested close to or on the coast, while White-tailed Eagles were absent from the high hills and largely restricted to the coastal zone or near large freshwater bodies, with both species present in landscapes with mountains close to low-lying wetlands and coasts. In other words, the picture of the two species' historic distribution, before effects of persecution had taken hold fully in western Scotland, seems to have been similar to recent distributions in relation to broad habitat type, both in our study area (Evans et al. Citation2010) and in western Norway, where both species have occurred together more continuously (Halley Citation1998). We suggest, therefore, that the fundamental hypothetical assumption for interspecific competition is also not supported by a rigorous consideration of the historical data.
We would conclude that, based on current evidence, there is no indication that White-tailed Eagles are having an adverse effect on Golden Eagles, although as noted by Whitfield et al. Citation(2002), future monitoring and research on this issue should continue.
ACKNOWLEDGEMENTS
In writing this article we owe a great debt of gratitude towards the late Mike Madders: a great friend and respected scientific colleague who was the major driver behind the origins of this study. His humour, insight and intellectual contribution are sorely missed. We are grateful to the landowners and farmers in our study areas for their cooperation and support. We thank Derek Hayward, Paul Haworth, Roger Broad, Fiona Harmer, Justin Grant, Rob Forrest, Alison MacLennan, Martin Carty, Steven MacDonald, Viv de Fresnes, David Miller, David Sexton and Brian Etheridge for assistance in collecting samples, and Ellen Wilson and David McLeod for database collation and GIS analyses, respectively. Alan Fielding provided helpful comments which improved an earlier draft. Comments from the editor and referees also improved a previous draft.
REFERENCES
- Alatalo , R. V. , Gustafsson , K. , Lundberg , A. and Ulfstrand , S. 1985 . Habitat shift of the Willow Tit Parus montanus in the absence of the Marsh Tit Parus palustris . Ornis Scandinavica , 16 : 121 – 128 . (doi:10.2307/3676477)
- Baxter , E. V. and Rintoul , J. V. 1953 . The Birds of Scotland , Edinburgh : Oliver & Boyd .
- Brom , T. G. 1986 . Microscopic identification of feathers and feather fragments of Palearctic birds . Bijdragen tot de Dierkunde , 56 : 181 – 204 .
- Cramp , S. and Simmons , K.E.L . 1980 . The Birds of the Western Palearctic Vol. 1 , Edited by: Cramp , S. and Simmons , K.E.L . Oxford : Oxford University Press .
- Evans , R. J. , Broad , R. A. , Duffy , K. , MacLennan , A. M. , Bainbridge , I. P. and Mudge , G. P. 2003 . “ Re-establishment of a breeding population of White-tailed Eagles in Scotland ” . In Sea Eagle 2000. Proceedings from an International Conference at Björkö, Sweden, 13–17 September 2000 , Edited by: Helander , B. , Marquiss , M. and Bowerman , W. 397 – 403 . Stockholm : Swedish Society for Nature Conservation .
- Evans , R. J , Wilson , J. D. , Amar , A. , Douse , A. , MacLennan , A. , Ratcliffe , N. and Whitfield , D. P. 2009 . Growth and demography of a re-introduced population of White-tailed Eagles Haliaeetus albicilla . Ibis , 151 : 244 – 254 . (doi:10.1111/j.1474-919X.2009.00908.x)
- Evans , R. J. , Pearce-Higgins , J. , Whitfield , D. P. , Grant , J. R. , MacLennan , A. and Reid , R. 2010 . Comparative nest habitat characteristics of sympatric White-tailed Haliaeetus albicilla and Golden Eagles Aquila chrysaetos in western Scotland . Bird Study , 57 : 473 – 482 . (doi:10.1080/00063657.2010.489317)
- Evans , R. J. , O'Toole , L. and Whitfield , D. P. 2012 . The history of eagles in Britain and Ireland: an ecological review of placename and documentary evidence from the last 1500 years . Bird Study , DOI:10.1080/00063657.2012.683388
- Gotelli , N. J. and Entsminger , G. L. 2006 . EcoSim: Null Models Software for Ecology , Version 7. Acquired Intelligence Inc. & Kesey-Bear, Jericho, VT, USA. Available online at: http://garyentsminger.com/ecosim.htm (accessed 27 November 2006)
- Green , R. E. , Pienkowski , M. W. and Love , J. A. 1996 . Long-term viability of the re-introduced population of the White-tailed Eagle Haliaeetus albicilla in Scotland . J. Appl. Ecol. , 33 : 357 – 368 . (doi:10.2307/2404757)
- Greene , H. W. and Jaksić , F. M. 1983 . Food–niche relationships among sympatric predators: effects of level of prey identification . Oikos , 40 : 151 – 154 . (doi:10.2307/3544212)
- Halley , D. J. 1998 . Golden and White-tailed Eagle in Scotland and Norway: co-existence, competition and environmental degradation . Br. Birds , 91 : 171 – 179 .
- Halley , D. J. and Gjershaug , J. O. 1998 . Inter- and intra-specific dominance relationships and feeding behaviour of Golden Eagles Aquila chrysaetos and Sea Eagles Haliaeetus albicilla at carcasses . Ibis , 140 : 295 – 301 . (doi:10.1111/j.1474-919X.1998.tb04392.x)
- Haworth , P. F. , McGrady , M. J. , Whitfield , D. P. , Fielding , A. H. and McLeod , D. R.A. 2006 . Ranging distance of Golden Eagles Aquila chrysaetos in western Scotland according to season and breeding status . Bird Study , 53 : 265 – 273 . (doi:10.1080/00063650609461442)
- Katzner , T. , Bragin , E. , Knick , S. and Smith , A. 2003 . Coexistence in a multispecies assemblage of eagles in central Asia . Condor , 105 : 538 – 551 . (doi:10.1650/7140)
- Katzner , T. E. , Bragin , E. A. , Knick , S. T. and Smith , A. T. 2005 . Relationship between demographics and diet specificity of Imperial Eagles Aquila heliaca in Kazakhstan . Ibis , 147 : 576 – 586 . (doi:10.1111/j.1474-919X.2005.00443.x)
- Korpimäki , E. 1987 . Dietary shifts, niche relationships and reproductive output of coexisting Kestrels and Long-eared Owls . Oecologia , 74 : 277 – 285 . (doi:10.1007/BF00379371)
- Krebs , C. J. 1989 . Ecological Methodology , New York : Harper & Row .
- Levins , R. 1968 . Evolution in Changing Environments , Princeton : Princeton University Press .
- Love , J. A. 1983 . The Return of the Sea Eagle , Cambridge : Cambridge University Press .
- Love , J. A. 2003 . “ A history of the White-tailed sea eagle in Scotland ” . In Sea Eagle 2000. Proceedings from an International Conference at Björkö, Sweden, 13–17 September 2000 , Edited by: Helander , B. , Marquiss , M. and Bowerman , W. pp. 39 – 50 . Stockholm : Swedish Society for Nature Conservation .
- McGrady , M. J. 1997 . Aquila chrysaetos Golden Eagle . Birds of the Western Palearctic Update , 1 : 99 – 114 .
- McGrady , M. J. , Grant , J. R. , Bainbridge , I. P. and McLeod , D. R.A. 2002 . A model of golden eagle (Aquila chrysaetos) ranging behavior . J. Rapt. Res. , 36 ( 1 Supplement ) : 62 – 69 .
- McLeod , D. R.A. , Whitfield , D. P. , Fielding , A. H. , Haworth , P. F. and McGrady , M. J. 2002 . Predicting home range use by golden eagles Aquila chrysaetos in western Scotland . Avian Sci. , 2 : 183 – 198 .
- MacArthur , R. H. and Levins , R. 1967 . The limiting similarity, convergence, and divergence of coexisting species . Am. Nat. , 101 : 377 – 385 . (doi:10.1086/282505)
- Madders , M. and Marquiss , M. 2003 . “ A comparison of the diet of White-tailed Eagles and Golden Eagles breeding in adjacent ranges in west Scotland ” . In Sea Eagle 2000. Proceedings from an International Conference at Björkö, Sweden, 13–17 September 2000 , Edited by: Helander , B. , Marquiss , M. and Bowerman , W. pp. 289 – 295 . Stockholm : Swedish Society for Nature Conservation .
- Mantel , N. 1967 . The detection of disease clustering and a generalized regression approach . Cancer Res. , 27 : 209 – 220 .
- Margurran , A. E. 2003 . Ecological Diversity and its Measurement , Princeton : Princeton University Press .
- Marquiss , M. , Madders , M. and Carss , D. N. 2003 . “ White-tailed Eagles and lambs: predation or scavenging? ” . In Sea Eagle 2000. Proceedings from an International Conference at Björkö, Sweden, 13–17 September 2000 , Edited by: Helander , B. , Marquiss , M. and Bowerman , W. pp. 297 – 305 . Stockholm : Swedish Society for Nature Conservation .
- Menge , B. A. 1979 . Coexistence between the seastars Asterias vulgaris and A. forbesi in a heterogeneous environment: a non-equilibrium explanation . Oecologia , 41 : 245 – 272 . (doi:10.1007/BF00377430)
- Mersmann , T. J. , Buehler , D. A. , Fraser , J. D. and Seegar , J. K.D. 1992 . Assessing bias in studies of bald eagle food habits . J. Wildl. Manage , 56 : 73 – 78 . (doi:10.2307/3808792)
- Morisita , M. 1959 . Measuring interspecific association and similarity between communities . Mem. Fac. Sci. Kyoto Univ. Ser. E (Biol.) , 3 : 65 – 80 .
- Newton , I. 1979 . Population Ecology of Raptors , Berkhamstead : Poyser .
- Nilsson , I. N. 1984 . Prey weight, food overlap, and reproductive output of potentially competing Long-eared and Tawny Owls . Ornis Scand. , 15 : 176 – 182 . (doi:10.2307/3675959)
- Ontiveros , D. , Pleguezuelos , J. M. and Caro , J. 2005 . Prey density, prey detectability and food habits: the case of Bonelli's eagle and the conservation measures . Biol. Cons. , 123 : 19 – 25 . (doi:10.1016/j.biocon.2004.10.004)
- Pálsdóttir , E. S. 2012 . Demographic patterns in winter carcass use by Golden Eagle and White-tailed Eagle in Finnmark, Norway. Unpublished Masters' Thesis, University of Tromsø, Norway .
- Pianka , E. R. 1973 . The structure of lizard communities . Ann. Rev. Ecol. Syst. , 4 : 53 – 74 . (doi:10.1146/annurev.es.04.110173.000413)
- Pontin , A. J. 1982 . Competition and Coexistence of Species , London : Pitman .
- Redpath , S. M. , Clarke , R. , Madders , M. and Thirgood , S. J. 2001 . Assessing raptor diet comparing pellets, prey remains, and observational data at Hen Harrier nests . Condor , 103 : 184 – 188 . (doi:10.1650/0010-5422(2001)103[0184:ARDCPP]2.0.CO;2)
- Rutz , C. 2003 . Assessing the breeding season diet of Goshawks Accipiter gentilis: biases of plucking analysis quantified by means of continuous radio-monitoring . J. Zool., Lond. , 259 : 209 – 217 . (doi:10.1017/S0952836902003175)
- Rutz , C. , Biljsma , R. , Marquiss , M. and Kenward , R. E. 2006 . Population limitation in the Northern Goshawk in Europe: a review with case studies . Stud. Avian Biol. , 31 : 158 – 197 .
- Sokal , R. R. and Rohlf , F. J. 1995 . Biometry , 3 , New York : Freeman .
- Smith , E. P. and Zaret , T. M. 1982 . Bias in estimating niche overlap . Ecology , 63 : 1248 – 1253 . (doi:10.2307/1938851)
- Steenhof , K. and Kochert , M. N. 1988 . Dietary responses of three raptor species to changing prey densities in a natural environment . J. Anim. Ecol. , 57 : 37 – 48 . (doi:10.2307/4761)
- Steenhof , K. , Kochert , M. N. and McDonald , T. L. 1997 . Interactive effects of prey and weather on golden eagle reproduction . J. Anim. Ecol. , 66 : 350 – 362 . (doi:10.2307/5981)
- Sulkava , S. , Tornberg , R. and Koivusaari , J. 1997 . Diet of the White-tailed Eagle Haliaeetus albicilla in Finland . Ornis Fenn. , 74 : 65 – 78 .
- Teerink , B. J. 1991 . Hair of West European Mammals , Cambridge : Cambridge University Press .
- Thom , V. M. 1986 . Birds in Scotland , Carlton : T. & A.D. Poyser .
- Watson , J. 1997 . The Golden Eagle , London : Poyser .
- Watson , J. , Leitch , A. F. and Broad , R. A. 1992 . The diet of the Sea Eagle Haliaeetus albicilla and Golden Eagle Aquila chrysaetos in western Scotland . Ibis , 134 : 27 – 31 . (doi:10.1111/j.1474-919X.1992.tb07225.x)
- Whitfield , D. P. and McLeod , D. R.A. 2007 . “ Range use in relation to nest site altitude in Golden Eagles. Report to Scottish Natural Heritage. Battleby, SNH ” .
- Whitfield , D. P. , McLeod , D. R.A. , Fielding , A. H. , Broad , R. A. , Evans , R. J. and Haworth , P. F. 2001 . The effects of forestry on golden eagles on the island of Mull, western Scotland . J. Appl. Ecol. , 38 : 1208 – 1220 . (doi:10.1046/j.0021-8901.2001.00675.x)
- Whitfield , D. P. , Evans , R. J. , Broad , R. A. , Fielding , A. H. , Haworth , P. F. , Madders , M. and McLeod , D. R.A. 2002 . Are reintroduced White-tailed Eagles in competition with Golden Eagles? . Scot. Birds , 23 : 36 – 45 .
- Whitfield , D. P. , Fielding , A. H. , Gregory , M. J.P. , Gordon , A. G. , McLeod , D. R.A. and Haworth , P. F. 2007 . Complex effects of habitat loss on golden eagles Aquila chrysaetos . Ibis , 149 : 26 – 36 . (doi:10.1111/j.1474-919X.2006.00591.x)
- Whitfield , D. P. , Fielding , A. H. , McLeod , D. R.A. and Haworth , P. F. 2008 . A conservation framework for Golden Eagles: implications for their conservation and management in Scotland , Scottish Natural Heritage Commissioned Report No. 193 (ROAME No. F05AC306). Scottish Natural Heritage, Battleby
- Wille , F. and Kampp , K. 1983 . Food of the White-tailed Eagle Haliaeetus albicilla in Greenland . Holarc. Ecol. , 6 : 81 – 88 .
- Willgohs , J. F. 1984 . White-tailed Eagles in Norway: Food, Breeding Biology, Competition and Enemies , WWF Norway, Trondheim [in Norwegian]