Abstract
Capsule The presence of farm animals increases the amount of larger insect prey (mainly large Diptera and Coleoptera) and decreases the proportion of smaller prey (Hymenoptera).
Aims To determine the relationships between presence of farm animals and diversity of diet of nestling Barn Swallows from 52 colonies located at sites with and without livestock.
Methods Diet was determined on the basis of faecal analysis. We used two approaches to quantify livestock based on their spatial distribution – a breeding site-focused approach and a village-focused approach.
Results Our results show a relationship between the presence of livestock and the size, number and composition of insect prey detected in the diet of nestling Barn Swallows. The results from the two approaches we used in our study were broadly consistent, although effects varied according to the number and types of animal being raised. The presence of domestic animals increases the amount of larger prey (mainly larger Diptera and Coleoptera) and decreases the proportion of smaller prey (Hymenoptera species). The effect of individual livestock species, mainly pigs and poultry alone, was considerably higher than the effects of pooling livestock species from Principal Component Analysis.
Conclusion Our results imply that Barn Swallows were foraging beyond the immediate area of the farm buildings in which they nest, suggest considerable adaptability in the species' diet and further emphasize the ecological role that livestock play in providing a source of prey of a required size rather specific identity. Furthermore, a statistically significant positive effect of average mass of prey (which is a direct consequence of the presence of cattle at a breeding site) on colony size suggests that livestock farming provides a more profitable prey community, which may ultimately increase local populations of Barn Swallow.
Agricultural environments impact populations of wild plant and animals, with farm animals in agricultural areas interacting with the environment in several different ways (Buckingham & Peach Citation2005). In general, the presence of livestock may be considered to function at two levels in an ecosystem. At one level there are concerns about chemical impacts such as the input of nutrients and other substances, including toxins to the soil and atmosphere. At a second level, livestock are a component of the environment and interact with other species (Söderstrom et al. Citation2001; Buckingham & Peach Citation2005). Some of these associations between domestic and wild animals, such as birds, can be commensal and mutualistic (sensu Holland & Bronstein Citation2008, Källander Citation1993). In many studies the presence of livestock has been identified as a key factor affecting the number and usage of different agricultural habitats by birds, including ground-and aerial-feeding bird guilds (Chamberlain & Fuller Citation2000, Ambrosini et al. Citation2002, Henderson et al. Citation2007), although to date there has been no study of the importance of different species of farm animals as a factor explaining variation in diet in birds.
The Barn Swallow Hirundo rustica belongs to a group of birds closely associated with humans (Cramp Citation1998, Turner Citation2006, Kragten et al. Citation2009). This species breeds mainly in buildings where farm animals – mainly cattle or pigs – are kept (Møller Citation1983, Citation1987, Citation2001, Ambrosini et al. Citation2002, Lubbe & De Snoo Citation2007). According to Ambrosini et al. (Citation2002, Citation2011) and Møller Citation(1983), livestock farming and the architecture of rural buildings are the main factors affecting the distribution and abundance of breeding Barn Swallows in Europe. On farms with cattle, Barn Swallows breed significantly earlier than on farms with no cattle; this difference can be accounted for by higher food abundance and higher temperature present in cowsheds which serve as breeding sites (Møller Citation1987, Ambrosini et al. Citation2002). Furthermore, Grüebler et al. Citation(2010) showed that the presence of livestock in the building with the nest and large numbers of manure heaps around breeding sites increased nestling survival in second broods, which increases the annual output of the Barn Swallow.
Recent changes in agricultural practices, especially the local termination of dairy farming and changes in the distribution of cattle, have been shown to be the main factors affecting a decline in number of Barn Swallows in most European countries (Turner Citation1994, Citation2006; Møller & Vansteenwegen Citation1997, Møller Citation2001, Evans et al. Citation2003a, Robinson et al. Citation2003, PECBMS Citation2007). Furthermore, recent data also demonstrate that the presence of cattle affects the abundance of Barn Swallows feeding in agricultural areas (Henderson et al. Citation2007). In some circumstances, the elimination of non-cropped habitats from the agricultural landscape and the disappearance of pastures may be additional reasons for the decline of the Barn Swallow in Europe (Evans et al. Citation2003b, Citation2007, Grüebler et al. Citation2008).
Møller (Citation1987, Citation2001) suggested that the decline of Barn Swallow populations is caused mainly by the reduction of food resources, notably large Diptera associated with cattle (mostly horse flies Tabanidae, hover flies Syrphidae and muscid flies Muscidae) which may constitute over 90% of the diet of Barn Swallows (Turner Citation2006). The reduction of these groups of insects is thought to be the cause of the decrease of reproductive success and the quality of offspring produced by Barn Swallows (Møller Citation1987, Citation2001, Ambrosini et al. Citation2002).
In general, the average size of prey taken by swallows is always larger than the average size of prey available in a given habitat (Mengelkoch et al. Citation2004; McCarty & Winkler Citation1999, Turner Citation1982, Bryant Citation1973, review in Hespenheide Citation1971). In the Barn Swallow, the inclusion of an item in the diet is dependent on the absolute abundance of large prey, although small items are included when relatively abundant (Turner Citation1982). However, there are no studies describing changes in the food composition of Barn Swallows, other hirundines or other aerial-feeding birds in relation to the presence of farm animals and there is little evidence of the preference that swallows may have for particular taxa. Besides, most data on diet composition and potential food resources of Barn Swallows come from only a few studies (for review see Cramp Citation1998).
Our key research question was to identify the relationship between livestock husbandry and diet composition of nestling Barn Swallows. We studied breeding sites of Barn Swallows characterized by a traditional system of animal rearing – i.e. small numbers of animals representing a few species (mainly poultry, cattle and pigs) that often remained in the open around the buildings – which is still extensive in southwest Poland. Therefore, our study characterized Barn Swallow diet under environmental and agricultural conditions which ceased to exist in Western Europe after the 1960s and 1970s. We hypothesized that the presence of farm animals could enhance certain insect populations associated with livestock husbandry. We quantified several variables of Barn Swallow nestling diet – e.g. biomass and number of different groups of main prey (flying insects) – and used these data to assess the effect of different species of farm animals. We also tested for any relationship between the size of breeding colonies and diet, in terms of the size or biomass of the prey caught.
METHODS
The diet of nestling Barn Swallows was investigated by identification of prey items from faecal sacs. Faecal sacs were collected from under occupied Barn Swallow nests during single visits at 52 sites (= buildings), including farm buildings used to breed various species (or mixed groups) of livestock (piggeries, cowsheds, chicken coops) located in 15 villages in southwestern Poland (extreme locations: 51°10′57″N, 16°57′18″E; 51°21′41″N, 18°41′01″E). At each breeding site we collected faeces once between 30 June and 4 August 2005; however, in the case of large accumulation of faeces under nests it is possible that we collected material from both early and late broods. We collected faecal samples only from a single nest in each building. Nests sampled were in farmstead buildings used for rearing of animals, mainly cattle and pigs (n = 34), or farm buildings in use where no livestock had been kept (n = 18), including one unused cowshed where cattle breeding had ceased a few years earlier. A feature of rural areas of Poland, as in most areas of central Europe, is that livestock are often bred in small farmstead buildings located in villages.
The number of nests and farm animals at breeding sites were established through counts and direct interviews with farm owners, respectively. Data on number of livestock for particular villages were collected from local authorities.
Diet characteristics
Faecal sacs were collected from under 52 occupied nests (= buildings) of barn swallows, located within 15 villages. There were 291 occupied swallow nests in all colonies from which faecal sacs were collected; colony sizes ranged from 1 to 25 breeding pairs (distribution of the number of nests/number of colonies: 1/12, 2/8, 3/4, 4/3, 5/8, 6/6, 8/3, 10/3, 17/2 and 25/3). Within individual villages faecal sacs were collected from between 1 and 8 nests; in three villages there were single nests; in the rest of the villages the number of nests/village was 2/5, 3/2, 6/3, 7/1 and 8/1. We pooled five faecal sacs randomly selected from all collected beneath a nest to quantify food items for each nest. Detailed description of methods used to identify faecal components and list prey items are presented elsewhere (Orłowski & Karg Citation2011).
We used 22 variables to describe nestling diet (hereafter ‘diet variables’): e.g. average number of prey items, average number of genera or species (taxa), average number of families, average mass of individual prey items, average mass of prey and total mass of prey, or average number, proportion and mass of prey and total mass of prey representing the four main insect orders. The variable ‘average mass of individual prey items’ was calculated on the basis of the mass of a single individual insect and is a synthetic measure, whereas ‘average mass of prey’ refers to the average mass for a given group of insects. Proportions of the four main orders of insects – namely Coleoptera, Diptera, Hymenoptera and Hemiptera – were calculated based on the actual number of representatives of these particular orders. The order Heteroptera was treated jointly with the order Homoptera, although the latter were scarcely represented (Heteroptera + Homoptera = Hemiptera; after McCarty & Winkler Citation1999). Mass of prey was calculated as dry mass, i.e. mg d.w.; these values were obtained from detailed measurements of insect weights based on analysis of 479 087 individuals of different taxa of insects (more details presented in Orłowski & Karg Citation2011).
Statistical analyses and diet modelling
Adult Barn Swallows collect food within 400–600 m of the nest to feed nestlings (Bryant & Turner Citation1982, Møller Citation1987, Ambrosini et al. Citation2002). Similarly, in our study area the village buildings typically did not extend beyond a diameter of about 500 m. Hence, we hypothesize that the diet of Barn Swallows might be related to the presence of farm animals not only at a small (= individual breeding site or building) scale but also larger (= village) spatial scale (). For this reason we modelled the effects of presence of farm animals on various diet properties (after arbitrarily selecting the 14 most key dietary variables) of the Barn Swallow at these two scales: (i) for an individual breeding site or building and (ii) for a village. We decided to omit from the analyses the proportions and average mass of main type of prey because they were combinations of the numbers and the total mass.
Table 1. Descriptive statistics of the number of farm animals at breeding sites (farmsteads) and villages located in southwestern Poland where faeces of nestling Barn Swallows Hirundo rustica were gathered in 2005; values given are means with 95% confidence intervals (95% CI).
Initially we performed a Canonical Correspondence Analysis (CCA) using MVSP software (Kovach Citation2005) in order to plot and so visualize the environmental gradient associated with farming of various livestock species onto the ordination of the diet properties. The CCA biplot was constructed with log-transformed variables, including the full set of 22 dependent dietary variables and 10 explanatory variables, including 9 livestock variables (i.e. cows, pigs, other large animals and poultry × two spatial scales, and sites without livestock, which had a value of 0 [absent] or 1 [present]) and number of nests in a colony.
Because of the high multicolinearity among numbers of some livestock species, both at the farm and village scale, which resulted from a few similar species of farm animals being reared at individual farmhouses, we conducted two independent Principal Component Analyses (PCA) with a varimax normalized rotation for breeding sites (farmhouses) and for villages including only four species of livestock (i.e. cows, pigs, other large animals and poultry). PCA variables derived from these analyses () were used later as explanatory variables for the modelling of diet properties.
Table 2. Component values and factor loadings of the Principal Component Analysis (PCA) of the characteristic farm animals at breeding sites and villages used as environmental predictors explaining the diet variables of nestling Barn Swallows Hirundo rustica; factor rotation: varimax normalized; figures in bold indicate the variable for which each factor indicated the most variability.
We hypothesized that the effects of individual species of livestock on the diet properties of nestling Barn Swallow might be more pronounced than for livestock variables derived from a PCA. Therefore we built two different multivariate Generalized Linear Models (GLZs) in order to assess all potential effects of individual species of farm animals as well as the livestock PCA variables on diet properties. In both models the number of farm animals bred at a breeding site was nested within variables describing number of animals reared at a village scale. We also included the ‘number of nests’ in a colony as a potential covariate in this model. This variable was nested within number of animals at a village scale as with number of farm animals at a breeding site. We applied a normal distribution and log-link function in GLZ models. Due to the fact that the main goal of our analyses was to assess the real response of the diet properties to environmental conditions associated with livestock farming, we presented one overall full model for each of 14 analysed dietary variables. To validate the models, the parameter estimates, standard errors and Wald (χ2) statistics were used to check the significance of the regression coefficients for each explanatory variable. Models were checked for over-dispersion using a likelihood ratio test (LRT). A goodness-of-fit of the overall model was assessed as residual deviation/residual degrees of freedom (res. dev./res. d.f.).
Finally, we tested the effect of the number of four species of farm animals bred at a farmhouse (breeding site) on the number of nests in a colony by fitting a multivariate general linear model (GLM) with normal distribution and log-link function.
All data were log-transformed and percentage data were square root–arcsine-transformed prior to analysis (Zar Citation1984) when necessary to satisfy assumptions of normality and homoscedasticity. The residuals of all models were approximately normally distributed. Statistical analyses were performed using GLM and GLZ modules of Statistica 7.0 (StatSoft 2006) and Excel software. The accepted significance level was P = 0.05.
RESULTS
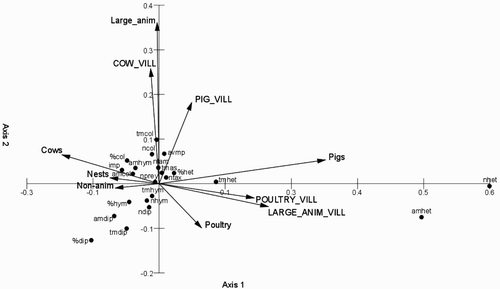
A CCA biplot used for grouping all variables measured (n = 32) showed that in the upper right quarter of two explanatory variables were grouped, pigs at breeding sites and villages, and five dietary variables (average number of taxa, total mass of prey, average mass of prey, total mass of Coleoptera, total mass of Hemiptera). The upper left quarter had four livestock variables grouped together (cows and other farm animals at breeding sites, cows at villages and number of nests) and eight dietary variables quite closely associated with them: average number of prey items, average mass of Coleoptera, average individual mass of prey, average mass of Hymenoptera, proportion of Coleoptera, average number of families, number of Coleoptera and total mass of Coleoptera. In the two lower quarters of the plot the remaining four explanatory variables and nine diet variables were grouped, respectively (see ), which generally indicated the relatively poor association of these dependent variables with farm animals reared both at breeding sites and at a village scale.
Table 3. General comparison of diet variables (mean ± 1 se) of nestling Barn Swallows Hirundo rustica from all 52 breeding sites (farmhouses), where there were various farm animals compared to sites without farm animals; large animals comprises cows, pigs and horses. Different letters indicate statistically significant differences (P ≤ 0.05) obtained in ANOVA GLM on paired comparisons between the two data sets.
Figure 1. CCA ordination biplot showing the locations of the 22 diet variables (explanation of abbreviations given in ) of nestling Barn Swallows Hirundo rustica in relation to different farm animals (cows, pigs; other large animals (= large animals) and poultry) reared in breeding sites (small letter) and villages (= vill; large letter); 10 explanatory variables, including 8 variables representing various species of livestock, number of nests in a colony (‘Nests’) and sites without livestock (‘Non-anim’) are depicted by arrows.
Farm animals
The number of farm animals at the 52 breeding sites was highly diverse. In general, cattle were most numerous at breeding sites (58.2% of total number of livestock) and poultry were most common in villages (44.5%) ().
The principal component analysis conducted with farm animals at breeding sites gave two main factors. PCA1 represented pigs and poultry and accounted for 37% of the variance contained in the original data set; PCA2 represented cattle, and encompassed 26% of the remaining variance (). Similarly, the PCA analysis with farm animals at the village scale revealed also two factors: PCA1 representing cattle and poultry, explaining 34% of total data variance, and PCA2 representing pigs and other large farm animals, explaining 33% of total data variance ().
General composition of the diet at the breeding sites
In total, 3152 prey items representing 98 taxa of invertebrates were identified from faecal sacs; a detailed taxonomical list of prey is presented separately elsewhere (Orłowski & Karg Citation2011). The proportions of the four dominant orders of insects were highly diverse across the 52 breeding sites (). On average, at all sites, the highest number, proportion and total mass was made up by Coleoptera (recorded in all sites). Diptera (recorded at 50 sites) and Hymenoptera (recorded at 51 sites) characterized the highest and smallest average mass of prey, respectively. Hemiptera (28 sites) were the least common prey by number and frequency at sites ().
Diet composition in relation to farm animals
Results of GLZ models for livestock variables derived from PCA conducted for the four species of farm animals () showed that at breeding sites PCA1 (= pigs and poultry) scores have a significant positive association with two diet variables: number and total mass of Hemiptera. PCA2 (= cows) showed a significant positive association with average mass of prey and total mass of Coleoptera. At the village scale, of the two PCA scores considered, only PCA1 (= cows and poultry) showed a positive association with average number of genera / species and number and total mass of Hymenoptera ().
Table 4. Results of Generalized Linear Models (GLZs) exploring the influence of two PCA scores of farm animals reared in breeding sites (= farmhouses) and villages on diet variables of nestling Barn Swallows Hirundo rustica; 1in mg dry weight (mg d.w.); statistical significance: * P < 0.05, ** P < 0.01, *** P < 0.001; bold indicates statistically significant effects.
The second set of GLZ models testing the effects of individual species of livestock showed that 11 of the 14 diet variables were significantly affected by the presence of farm animals. In total we found 19 significant effects (13 positive and 6 negative) between diet variables and number of farm animals at both spatial scales, including 13 significant ones for breeding sites and 6 for villages (). Considering the breeding sites, the highest number of significant effects (both n = 5) were found for pigs (always positively correlated with average number of genera/species, total mass of prey, number and total mass of Hemiptera and total mass of Coleoptera). The number of pigs at the village scale also significantly positively affected the average individual mass of prey and number of Hymenoptera. The results for poultry were less consistent and there was only one positive (average individual mass of prey) and four negative effects (average number of prey items, average number of genera/species, average number of families of prey, number of Diptera). The only negative effects were found for cattle at the village scale (total mass of prey and number of Hymenoptera). In the case of the other large farm animals there were positive associations: at the breeding sites, the average mass of prey and total mass of Coleoptera, and at the village scale, average number of genera/species and number and total mass of Hymenoptera ().
Table 5. Results of Generalized Linear Models (GLZs) exploring the influence of number of nests and various farm animals reared at breeding sites (= farmhouses) and villages on diet variables of nestling Barn Swallows Hirundo rustica. Statistical significance: * P < 0.05, ** P < 0.01, *** P < 0.001; bold indicates statistically significant effects.
Diet variation in relation to the size of the Barn Swallow colony
The multivariate GLM model testing the effect of four species of farm animals bred at a farmhouse (breeding site) scale on the number of nests in a colony showed a significant overall result (GLM, R = 0.82, F 4,47 = 24.2, P < 0.0001); however, only the number of cattle had a significant individual effect on the size of a colony (F = 70.5, P < 0.0001); the number of pigs, other large animals and poultry did not affect the colony size (F = 0.01, 2.47 and 1.48, P ≥ 0.12, respectively). Statistically significant (positive) effects of colony size on diet variables were found only in the case of average mass of prey ().
DISCUSSION
Five major conclusions arise from our study. First, the presence of farm animals is an important factor that shapes some of the diet variables of nestling Barn Swallows. Secondly, both our breeding site-focused approach and a village-focused approach show an impact on the diet properties depending on the type and numbers of domestic animals being raised. Thirdly, individual species of farm animals (from GLZ models; ) showed greater effects than pooled combinations of livestock species from PCA (). Fourthly, in general, the presence of farm animals increases the importance of larger prey (mainly large Diptera and Coleoptera) and decreases the share of less profitable (= smaller) prey (Hymenoptera). Fifthly, the average mass of prey is significantly positively correlated with the colony size of Barn Swallows.
In general, the PCA based on the number of different species of farm animals revealed variation in the livestock being kept in rural areas of Poland. At both spatial scales of this analysis, most variation exists in the number of pigs and cattle, and secondarily in the number of poultry and other large animals. It seems that these differences resulted from varying proportions of the four species of livestock at both spatial scales, especially in the case of poultry and cows, which were most numerous in villages and breeding sites, respectively (). Our results match the traditional system of animal rearing as described in the introduction, i.e. the keeping of a small number of animals representing a few species of livestock, especially pigs and poultry. Both of the two scales used in our study are complementary in explaining the dietary properties of the Barn Swallow, although effects vary according to the number and types of animal being raised at the different scales. However, earlier findings on the quality of Barn Swallow nestlings showed that the environmental conditions at the scale of nesting microhabitat are more influential than macro-environmental conditions at the scale of foraging range (Ambrosini & Saino Citation2010).
Our first assessment of interactions of diet properties with characteristics of farm animals from the PCA with the use of GLZ models showed that the effect of farm animals on diet of the Barn Swallow is a scale-dependent phenomenon. The models also showed importantly that the average mass of prey was significantly positively related to the colony size of Barn Swallows. At breeding sites, pigs and poultry (PCA1) were significantly and positively associated with the number and total mass of Hemiptera, whereas cows (PCA2) increased the average mass of prey and total mass of Coleoptera. At the village scale, only the cows and poultry (PCA1) showed a positive association with average number of taxa and number and total mass of Hymenoptera. It seems that our positive relation between the size of colony of Barn Swallow and the number of cows (GLM model with four livestock species) and average mass of prey (GLZ model with PCA livestock variables) matches earlier studies of the ecology of Barn Swallow, possibly suggesting a direct link between livestock farming and abundance of larger prey types near the breeding site, which may translate into their exploitation near farm buildings where cattle are kept (Waugh Citation1978, Møller Citation1987, Citation2001, Ambrosini et al. Citation2002, Grüebler et al. Citation2010). Furthermore, our village-scale analysis – showing a positive association between cows and poultry and number of taxa and total mass of Hymenoptera – could imply that Barn Swallows were foraging beyond the immediate area of the farm buildings, utilizing a more diverse prey community (although swallow foraging was not measured directly) when these species of livestock were reared outside the breeding sites. These results agree with earlier relationships detected by Bryant and Turner Citation(1982), who found that both food abundance and prey size were negatively correlated with foraging distance, which suggests considerable adaptability in the species' diet and further emphasizes the important role that cattle and pigs play in providing a source of prey of a required size, rather than necessarily supporting prey of a particular species. These aspects of the feeding ecology of the Barn Swallow have never been described before.
The impact of livestock on the diet of nestling Barn Swallows is evident from comparisons of real values of dietary variables between breeding sites with and without farm animals (). These comparisons show a significant increase of average individual mass of all prey and Dipteran prey at breeding sites with pigs, large animals and cattle, and decrease in the total number and average mass of Hymenoptera (up to more than threefold) at breeding sites without farm animals. These findings clearly imply that farm animal presence does influence diet selectivity in Barn Swallows; in the presence of livestock, larger Diptera become a more important component of the diet and smaller (lighter) Hymenoptera much less.
The second set of GLZ models testing the effects of individual species of livestock shows higher complexity and variability of interactions between diet properties and individual species of farm animals. In general, our previous linkages of PCA results with diet variables showed fewer significant results in comparison to GLZs with individual species of livestock. It seems likely that the pooled characteristics of different farm animals masked a degree of complexity – each species has different effects (i.e. features of farm buildings, management of livestock faeces and differentiated insect fauna).
Overall, the results from both sets of GLZ models might suggest a fundamental trade-off between the presence of different species of livestock and the size, number and composition of prey that best provide optimal foraging opportunities for swallows and the scale over which they will forage optimally (Turner Citation1982). This is confirmed by the effects shown in the GLZ models with individual species of livestock, where the highest number of significant effects was found for pigs, poultry and other large farm animals, which generally increase the contribution of larger prey (Coleoptera, Hemiptera) in the diet, with the exception of pigs at the village scale which affected the number of smaller prey, namely Hymenoptera. The four negative effects of poultry at a breeding site scale (affecting the average number of prey items, average number of taxa, average number of families of prey, and number of Diptera) and only negative effects in the case of the cattle at the village scale (which affected the total mass of prey and number of Hymenoptera), suggest that Barn Swallows likely feed outside the breeding sites dependent on the livestock present in the area. In contrast, and importantly, these findings apparently showed that the presence of livestock negatively affect the contribution in the diet of the smallest prey items (Hymenoptera; see ). Turner Citation(1982) also found that large prey (≥ 1.5 mg d.w.) were incorporated into Barn Swallow diet when their abundance in the environment increases, whereas the number of small prey items (< 1.5 mg d.w.) found in the diet was not correlated with their abundance in the air. Moreover, large prey items were generally heavily selected when they were rare in the environment while small prey were often avoided (Turner Citation1982). One result from the present study that differed from previous studies was the relatively small proportion of Diptera and predominant proportion of Coleoptera present in the diet of nestling Barn Swallows (; and see Orłowski & Karg Citation2011 for a comment on potential biases in assessment of diet composition base on analysis of faeces). In the case of our study, the positive association between proportion of Coleoptera, mass of Diptera and number of livestock is probably a result of food selectivity by Barn Swallow, i.e. selection of less mobile and heavier prey (cf. Bryant & Turner Citation1982), particularly species linked with presence of dung and manure of large farm animals (i.e. from the high densities and biomass of these insects, mainly Coleoptera, cf. Hanski & Koskela Citation1977, Jones Citation1990, Sanchez-Pinero & Avila Citation2004). Furthermore, the statistically significant positive effect of colony size (which is likely to be a direct consequence of the presence of cattle at a breeding site) on the average mass of prey shown in GLZ model with PCA livestock variables could suggest that the livestock farming provides a more profitable prey community. This then results in higher food resources near farm buildings where cattle or other domestic animals are kept and ultimately increases the number of local populations of Barn Swallow (Møller Citation1987, Ambrosini et al. Citation2002). However, our results are only correlations and do not show direct causation.
Dung and manure on pastures and near farm buildings constitute nutrient-rich food resources that provide a diverse community not only of coprophagous species of Coleoptera, but also other trophic groups of these insects (i.e. omnivores, predators and detrivores) (Hanski & Koskela Citation1977, Horgan Citation2002, Sanchez-Pinero & Avila Citation2004, Pimsler Citation2007). Although some earlier results indicate that dunghills may serve as the main foraging sites and manure facilities positively influence colony size and increase survival of nestling Barn Swallows (Møller Citation1983, Ahnström et al. Citation2008; Grüebler et al. Citation2010), so far the invertebrate fauna at such places has not been associated with diet of any of the swallows foraging at such sites. In our data set, however, coleopterans associated with dung and manure made up jointly 23.5% of the number and 24.3% of the total biomass of all representatives of the order (Orłowski & Karg Citation2011). The breeding sites from which faeces of Barn Swallows were collected were located mostly within areas of organic small-scale farming, where organic fertilizers (i.e. manure and slurry) are commonly applied (Bengtsson et al. Citation2005, Lubbe & De De Snoo 2008, Kragten et al. Citation2011), a practice which enhances the species richness and biomass of many groups of insects, including Carabidae. Furthermore, it is noteworthy that the use of organic fertilizers significantly increases the abundance of aerial insects over crop fields (discussed in Evans et al. Citation2007, Kragten et al. Citation2011).
CONCLUSIONS AND RESEARCH IMPLICATIONS
Our results considerably extend earlier studies on the feeding ecology of Barn Swallows, showing how diet changes in relation to the presence of livestock. For example, Evans et al. Citation(2007) describe Barn Swallow diet across broadly defined farmland habitats across Europe, but there have been no detailed studies describing the diet of this species in relation to the presence of livestock. Our results suggest that the preservation of a diverse agricultural landscape is not as essential as the maintenance of livestock, which may be the most important and consistent factor associated with presence of the bird species in agricultural areas.
Owing to the fact that our analysis covered breeding sites where several species of farm animals were raised simultaneously, it is difficult to disentangle the effects of the different species of livestock. Our results from the region of more traditional methods of animal husbandry in southwestern Poland are also difficult to compare with regions with more industrialized livestock production that can be found in Western Europe. Despite the correlational character of our study, we probably do characterize the diet of Barn Swallows as a function of the agricultural practices in Eastern European, where farmland bird populations have declined considerably in recent years (PECBMS Citation2007). Future research would profit from comparisons with other populations of Barn Swallows from rural areas characterized by more diverse systems of agricultural management and methods of animal breeding. Finally, there is likely to be a direct link between the reproductive success in Barn Swallows and food availability due to the presence of livestock found on dairy farms (Møller Citation2001, Grüebler et al. Citation2010), which suggest that additional investigations on the breeding ecology and prey selection by Barn Swallows in central and eastern Europe on these particular livestock farms are especially needed.
ACKNOWLEDGEMENTS
We are very grateful for Will Cresswell, Michael A. Patten, Simon Gillings, Piotr Tryjanowski, Krzysztof Kujawa and anonymous reviewers for valuable comments and discussion, which greatly improved the value of the article; and Joanna Czarnecka for statistical advice. We appreciate the improvements in English usage made by Peter Lowther through the Association of Field Ornithologists' programme of editorial assistance and Wenesa Synowiec. We thank Józef, Witek, Agnieszka and Viola Ciach and Stanisław Wojaczek for accommodation, technical support and assistance during the field study.
REFERENCES
- Ambrosini , R. , Bolzern , A. , Canova , L. , Arieni , S. , Møller , A. P. and Saino , N. 2002 . The distribution and colony size of Barn Swallow in relation to agricultural land use . J. Appl. Ecol. , 39 : 524 – 534 . (doi:10.1046/j.1365-2664.2002.00721.x)
- Ambrosini , R. and Saino , N. 2010 . Environmental effects at two nested spatial scales on habitat choice and breeding performance of barn swallow . Evol. Ecol. , 24 : 491 – 508 . (doi:10.1007/s10682-009-9321-5)
- Ambrosini , R. , Bani , L. , Massimino , D. , Fornasari , L. and Saino , N. 2011 . Large-scale spatial distribution of breeding Barn Swallows in relation to cattle farming . Bird Study , 58 : 495 – 505 . (doi:10.1080/00063657.2011.609883)
- Ahnström , J. , Berg , A. and Söderlund , H. 2008 . Birds on farmsteads – effects of landscape and farming characterisrics . Ornis Fenn. , 85 : 98 – 108 .
- Bengtsson , J. , Ahnström , J. and Weibull , A. 2005 . The effects of organic agriculture on biodiversity and abundance: a meta analysis . J. Appl. Ecol. , 42 : 261 – 269 . (doi:10.1111/j.1365-2664.2005.01005.x)
- Bryant , D. M. 1973 . The factors influencing the selection of food by the house martin (Delichon urbica (L.)) . J. Anim. Ecol. , 42 : 539 – 564 . (doi:10.2307/3123)
- Bryant , D. M. and Turner , A. K. 1982 . Central place foraging by swallows (Hirundinidae): the question of load size . Anim. Behav. , 30 : 845 – 856 . (doi:10.1016/S0003-3472(82)80158-9)
- Buckingham , D. L. and Peach , W. J. 2005 . The influence of livestock management on habitat quality for farmland birds . Anim. Sci. , 81 : 199 – 203 . (doi:10.1079/ASC50700199)
- Chamberlain , D. E. and Fuller , R. J. 2000 . Local extinctions and changes in species richness of lowland farmland birds in England and Wales in relation to recent changes in agricultural land-use . Agric. Ecos. Environ. , 78 : 1 – 17 . (doi:10.1016/S0167-8809(99)00105-X)
- Cramp , S. 1998 . The Complete Birds of the Western Palaearctic on CD-ROM , Edited by: Cramp , S. Oxford , , UK : Oxford University Press .
- Evans , K. L. , Wilson , J. D. and Bradbury , R. B. 2003a . Swallow Hirundo rustica population trends in England: data from repeated historical surveys . Bird Study , 50 : 178 – 181 . (doi:10.1080/00063650309461310)
- Evans , K. L. , Bradbury , R. B. and Wilson , J. D. 2003b . Selection of hedgerows by Swallows Hirundo rustica foraging on farmland: the influence of local habitat and weather . Bird Study , 50 : 8 – 14 . (doi:10.1080/00063650309461284)
- Evans , K. L. , Wilson , J. D. and Bradbury , R. B. 2007 . Effects of crop type and aerial invertebrate abundance on foraging barn swallows Hirundo rustica . Agric. Ecos. Environ. , 122 : 267 – 273 . (doi:10.1016/j.agee.2007.01.015)
- Grüebler , M. U. , Korner-Nievergelt , F. and Von Hirschheydt , J. 2010 . The reproductive benefits of livestock farming in barn swallows Hirundo rustica: quality of nest site or foraging habitat? . J. Appl. Ecol. , 47 : 1340 – 1347 . (doi:10.1111/j.1365-2664.2010.01873.x)
- Grüebler , M. U. , Morand , M. and Naef-Daenzer , B. 2008 . A predictive model of the density of airborne insects in agricultural environments . Agric. Ecos. Environ. , 123 : 75 – 80 . (doi:10.1016/j.agee.2007.05.001)
- Hanski , I. and Koskela , H. 1977 . Niche relations among dung-inhabiting beetles . Oecologia , 28 : 203 – 231 .
- Henderson , I. , Holt , C. and Vickery , J. 2007 . National and regional patterns of habitat association with foraging Barn Swallows Hirundo rustica in the UK . Bird Study , 54 : 371 – 377 . (doi:10.1080/00063650709461497)
- Hespenheide , H. A. 1971 . Food preference and the extent of overlap in some insectivorous birds, with special reference to the Tyrannidae . Ibis , 113 : 59 – 72 . (doi:10.1111/j.1474-919X.1971.tb05123.x)
- Holland , J. N. and Bronstein , J. 2008 . “ Mutualism ” . In Population Dynamics. Population Dynamics Edited by: Jorgensen , S. E. and Fath , B. D. Vol. 3 , of Encyclopedia of Ecology: 2485–2491. Elsevier, Oxford
- Horgan , F. G. 2002 . Shady field boundaries and the colonisation of dung by coprophagous beetles in Central American pastures . Agric. Ecos. Environ. , 91 : 25 – 36 . (doi:10.1016/S0167-8809(01)00261-4)
- Jones , G. 1990 . Prey selection by the greater horseshoe bat (Rhinolophus ferrumequinum): optimal foraging by echolocation? . J. Anim. Ecol. , 59 : 587 – 602 . (doi:10.2307/4882)
- Källander , H. 1993 . Commensal feeding relationships between yellow wagtails Motacilla flava and cattle . Ibis , 135 : 97 – 100 . (doi:10.1111/j.1474-919X.1993.tb02815.x)
- Kovach , W. L. 2005 . MVSP – A MultiVariate Statistical Package for Windows, ver. 3.1 , Pentraeth, Wales , , UK : Kovach Computing Services .
- Kragten , S. , Reinstra , E. and Gertenaar , E. 2009 . Breeding Barn Swallows Hirundo rustica on organic and conventional arable farms in the Netherlands . J. Ornithol , 150 : 515 – 518 . (doi:10.1007/s10336-009-0383-5)
- Kragten , S. , Ramiro , S. M. , van der Poll , R. , Wang , J. and de Snoo , G. R. 2011 . Abundance of invertebrate prey for birds on organic and conventional arable farms in the Netherlands . Bird Conserv. Intern. , 21 : 1 – 11 . (doi:10.1017/S0959270910000079)
- Lubbe , S. K. and De Snoo , G. R. 2007 . Effect of dairy farm management on Swallow Hirundo rustica abundance in The Netherland . Bird Study , 54 : 176 – 181 . (doi:10.1080/00063650709461473)
- McCarty , J. P. and Winkler , D. W. 1999 . Foraging ecology and diet selectivity of tree swallows feeding nestlings . Condor , 101 : 246 – 254 . (doi:10.2307/1369987)
- Mengelkoch , J. M. , Niemi , G. J. and Regal , R. R. 2004 . Diet of nestling tree swallow . Condor , 106 : 423 – 428 . (doi:10.1650/7341)
- Møller , A. P. 1983 . Breeding habitat selection in the Swallow Hirundo rustica . Bird Study , 30 : 134 – 142 . (doi:10.1080/00063658309476788)
- Møller , A. P. 1987 . Advantages and disadvantages of coloniality in the swallow . Hirundo rustica. Anim. Behav. , 35 : 819 – 832 . (doi:10.1016/S0003-3472(87)80118-5)
- Møller , A. P. 2001 . The effect of dairy farming on barn swallow Hirundo rustica abundance, distribution and reproduction . J. Appl. Ecol. , 38 : 378 – 389 . (doi:10.1046/j.1365-2664.2001.00593.x)
- Møller , A. P. and Vansteenwegen , C. 1997 . “ Barn Swallow Hirundo rustica ” . In The EBCC Atlas of European Breeding Birds , Edited by: Hagemeijer , W. J. and Blair , M. 478 – 479 . London , , UK : T & AD Poyser .
- Orłowski , G. and Karg , J. 2011 . Diet of nestling Barn Swallows Hirundo rustica in rural areas of Poland . Cent. Eur. J. Biol. , 6 : 1023 – 1035 . (doi:10.2478/s11535-011-0070-4)
- PECBMS . 2007 . State of Europe's Common Birds, 2007 , Prague , , Czech Republic : CSO/RSPB . 23 pages
- Pimsler , M. L. 2007 . “ A survey of the dung beetles in cattle manure on pastures of a organic and a conventional dairy farms in New York state ” . Honors Thesis, Department of Entomology, Cornell University, New York. Available from http://ecommons.library.cornell.edu (accessed 15 December 2009)
- Robinson , R. A. , Crick , H. and Peach , W. 2003 . Population trends of Swallows Hirundo rustica breeding in Britain . Bird Study , 50 : 1 – 7 . (doi:10.1080/00063650309461283)
- Sanchez-Pinero , F. and Avila , J. M. 2004 . Dung–insects community composition in arid zones of south-eastern Spain . J. Arid Environ. , 56 : 303 – 327 . (doi:10.1016/S0140-1963(03)00057-0)
- Söderstrom , B. , Pärt , T. and Linnarsson , E. 2001 . Grazing effects on between-year variation of farmland bird communities . Ecol. Appl. , 11 : 1141 – 1150 . (doi:10.1890/1051-0761(2001)011[1141:GEOBYV]2.0.CO;2)
- Turner , A. K. 1982 . Optimal foraging by the swallow (Hirundo rustica, L.): prey size selection . Anim. Behav. , 30 : 862 – 872 . (doi:10.1016/S0003-3472(82)80160-7)
- Turner , A. K. 1994 . “ Swallow Hirundo rustica ” . In Birds in Europe: Their Conservation Status , Edited by: Tucker , G. M. and Heath , M. 370 – 371 . Cambridge , , UK : BirdLife International .
- Turner , A. K. 2006 . The Barn Swallow , London : T & AD Poyser .
- Waugh, D.R. 1978. Predation strategies in aerial feeding birds. PhD thesis. Department of Biology, University of Stirling.
- Zar , J. H. 1984 . Biostatistical Analysis , 2 , London : Prentice Hall .