Abstract
Capsule The Eurasian Marsh Harrier shows a leapfrog migration pattern with individuals breeding in Northern and Eastern Europe wintering further from their breeding grounds than individuals belonging to the populations of Western and Southern Europe.
Aims We analyse wintering distribution of Marsh Harriers in relation to their breeding grounds to identify differential wintering area selection strategies.
Methods We used ring recovery data of 320 individuals from the European population. Latitude and longitude of ringing sites as well as age-class were used to explain variation in the distances between breeding areas and wintering sites. Moreover, we analyse differences in migration distances performed by males and females and differences among individuals wintering in sub-Saharan Africa.
Results Distance covered to reach the wintering ground increases with increasing latitude and longitude of the breeding sites. Marsh Harriers breeding in Northern and Eastern Europe migrate farther than individuals belonging to the populations within Western and Southern Europe, with no effect of age-classes in relation to the latitude of wintering. Males show a tendency to migrate longer distances than females. Finally, when taking into account only birds wintering south of the Sahara desert, juveniles winter further west than adults.
Conclusions The results are consistent with the assumption that this species shows a leapfrog migration. We suggest that the difference in the longitudinal distribution of juveniles and adults wintering south of Sahara could be related to the different ability of experienced individuals to compensate for the wind drift of dominant easterly winds during migration over the desert in autumn.
Among migratory bird species, several factors can promote variation in migration behaviour (Newton 2008). Within a given population, migration timing and/or distance can vary according to age- and/or sex-classes (differential migration; Cristol et al. Citation1999, Bai & Schmidt Citation2012). Among different populations, geographical location of breeding areas can be another source of variation. The latter can result in a latitudinal segregation among wintering populations, as in the following patterns: ‘chain migration’, where winter quarters of different populations are situated in the same latitudinal sequence as the breeding areas and ‘leapfrog migration’, where winter ranges are in a reverse latitudinal arrangement compared with breeding areas, i.e. the most northern breeding population winter farthest south (Salomonsen Citation1955, Smith et al. Citation2003).
Although many different cases of leapfrog migration are reported among American raptors (Bildstein Citation2006), only a few cases are known in Europe: the Common Kestrel (Falco tinnunculus), the Eurasian Sparrowhawk (Accipiter nisus) and the Common Buzzard (Buteo buteo; Wallin et al. Citation1987, Ferguson-Lees & Christie Citation2001). The third case is the most documented one; in particular individuals breeding in Finland and Siberia migrate by leapfrogging over partially migratory and sedentary populations in central and southern Europe as well as in Japan (Alerstam Citation1990).
The Eurasian Marsh Harrier (Circus aeruginosus) is a good model species to investigate differential migration because the European breeding population is widely distributed, both from the longitudinal and latitudinal point of view. The wintering grounds of this species are also extensively distributed, from Western Europe to sub-Saharan Africa (Sternalski et al. Citation2008, Strandberg et al. Citation2008).
It is widely reported that European populations show a leapfrog migration (e.g. Brown et al. Citation1982), but this hypothesis has never been thoroughly investigated. Support is, however, provided by studies that show individuals from Western France are mostly resident (Bavoux et al. Citation1992, Citation1994) while populations breeding in Scandinavian are largely long-distance migrants (Strandberg et al. Citation2008). Some recent studies have investigated the movements of populations from limited parts of the marsh harriers breeding range by visual observations, traditional radio-telemetry and, more recently, by satellite tracking (Sternalski et al. Citation2008, Strandberg et al. Citation2008, Klaassen et al. Citation2011, Agostini & Panuccio Citation2010). Here we analyse migration patterns of Eurasian Marsh Harriers using ringing recovery data from a large portion of Europe. Although ringing studies may have biases – for example, towards regions with denser human populations and stronger hunting pressure – if we assume that the proportion of age- and sex-classes represented in recoveries is not biased, then interesting insights into migration patterns in relation to sex and/or age can be obtained. A previous study compared results of ringing recovery data with results provided by satellite tracking of some species of raptors including the Eurasian Marsh Harrier (Strandberg et al. Citation2009b) and suggested that the interpretation of the temporal patterns from ringing recovery data might be problematic. Nevertheless ring recoveries and satellite tracking have been shown to provide more or less the same geographical distribution. Furthermore, ringing recoveries provide a larger sample size of individuals than satellite tracking. Our aim was to compare the distances between breeding and wintering grounds of Eurasian Marsh Harriers in relation to longitude, latitude, age- and sex-class, in order to distinguish which factors predict variation in migration behaviour.
METHODS
Ringing data of Eurasian Marsh Harriers were obtained from the EURING database. In the analysis we considered only highly accurate data concerning dates and coordinates of capture and recovery with a maximum error of 50 km (EURING categories from 0 to 4). After filtering, we retained for analysis a total of 320 individuals ringed during the breeding season (10 May–10 July) and recovered during winter (15 October–28 February), selecting these periods according to the migratory phenology of the species (Génsbøl Citation2004, Strandberg et al. Citation2008, Agostini & Panuccio Citation2010). It has been reported that some Eurasian Marsh Harriers do not settle in the wintering grounds before late autumn (Strandberg et al. Citation2008); for this reason, we also repeated all the analysis using a subset of data (15 November–28 February). As the results were the same we chose to present and discuss the full data set. The time span of data is shown in and the largest amount of data comes from the birds breeding in central Europe (81.6% of the data included in the latitude band 49.00°–53.99°N). Numbers of juvenile and adult Eurasian Marsh Harriers (age at recapture) were not significantly different (χ2 = 2.1, d.f. = 1, n.s.). In order to investigate general patterns of migration distance, a Generalized Linear Model (GLM) with negative binomial distribution was applied using the ‘MASS’ package included in R software (Venables & Riley Citation2002). Latitude and longitude of ringing sites as well as age-class (juvenile, adult) were used as independent factors to explain variation in the loxodrome distance (km) between the ringing site (breeding area or birthplace) and the place of recovery (wintering site). In the analysis the age of birds was defined according to the recovery data, and therefore a bird ringed at the nest and recovered at least in its second or following winter was considered an adult. In the case of 43 individuals with known sex, we used a z-test to compare the mean migratory distances of the two sex categories (males, females) after logarithmic transformation of data, and an F test to compare variances (Fowler & Cohen Citation1992). We tested the hypothesis that climate change could affect the travel distances between breeding and wintering territories by correlating the year of recovery and the migratory distance (km). Age-related variation concerning the location of wintering sub-Saharan areas was investigated using a Mann–Whitney U-test to compare the longitudes of wintering sites. Because juveniles may be more likely to make extensive exploratory movements before they establish their wintering site, we considered the time effect as a possible source of sampling bias: i.e. juveniles might be recovered significantly earlier than adults in winter. For this reason we compared the median date of recovery using a Mann–Whitney U-test (Fowler & Cohen Citation1992). Finally, ArcGIS 9.1 software was used to project and visually inspect the data.
Table 1. Spatial and temporal characteristics of the ringing recoveries data set used in the present study.
RESULTS
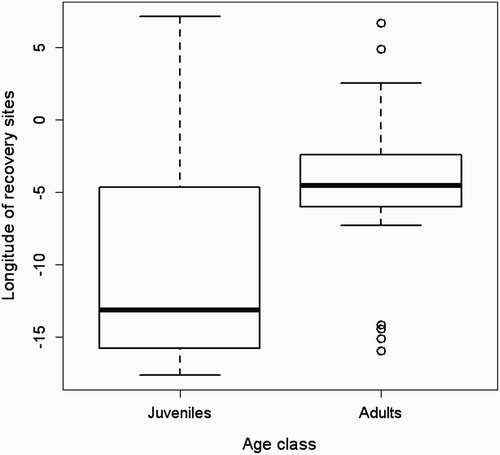
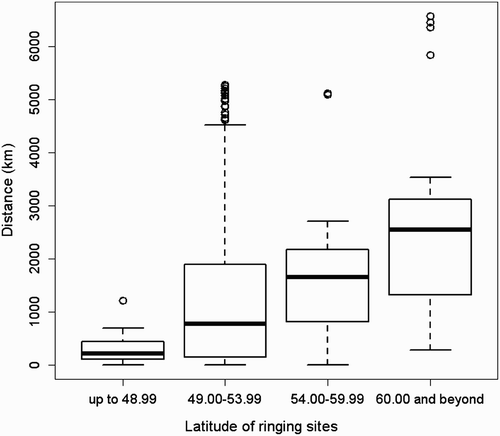
Our GLM shows a significant positive relationship between the length of the migratory journey and both latitude (F = 10.1; d.f. = 317; P < 0.01) and longitude (F = 14.9; d.f. = 316; P < 0.001) of the breeding sites but not with age-class (F= 0.0002; d.f. = 318; P > 0.05), showing that individuals belonging to northern and eastern populations migrate farther than individuals breeding in Western and Southern Europe (). Considering individuals ringed south of 49°N none wintered in Africa, while those from Great Britain and northern France did not cross the Sahara desert ( & 5). Eurasian Marsh Harriers belonging to the populations of Netherlands and Belgium and individuals breeding in central–eastern Europe (Germany, Poland and former Czechoslovakia) show a wide winter distribution both in Europe and Africa (, c). Finally those from Northern Europe show a tendency to migrate farther away than the other populations ( & 5). Among the two geographical factors, longitude (parameter estimate: 0.06) appeared to be stronger than latitude (parameter estimate: 0.008) in helping to explain the increases in the distances travelled. Among 43 sexed birds the mean migration distance was higher for males (1794.5 ± 713 km; n = 13) than for females (1118.4 ± 237.9 km; n = 30; & ), but this difference was not significant (z = 1.3, n.s.). We found no significant correlation between migratory distance and the recovery year (R = 0.06, n.s.). Forty-one individuals were recovered in Africa south of the Sahara Desert, and among them 20 were juveniles and 21 adults (48.8% vs. 51.2%; χ2 = 0.02, d.f. = 1, n.s.). Comparing the longitude of wintering sites in sub-Saharan Africa, juveniles wintered significantly further west than adults (; Mann–Whitney U-test; U = 121, P = 0.02), and this result was not due to juveniles being ringed more commonly in the west, because there were no differences in the longitude of breeding site (Mann–Whitney U-test; U = 164, n.s.). We did not find significant differences when comparing dates of recovery of juveniles and adults wintering south of Sahara (Mann–Whitney U-test = 166.5, n.s.).
Figure 1. Map showing ring and recovery sites of Eurasian Marsh Harriers breeding in Europe (black = ringing sites, white = recovery sites). A, circles = individuals coming from areas located south of 49°N; triangles = individuals from Great Britain and northern France. B, individuals from Belgium and Netherlands. C, individuals from Germany, Poland, former Czechoslovakia. D, circles = individuals coming from areas located north of 60°N; triangles = individuals breeding in the area included between 54° and 59.99°N.
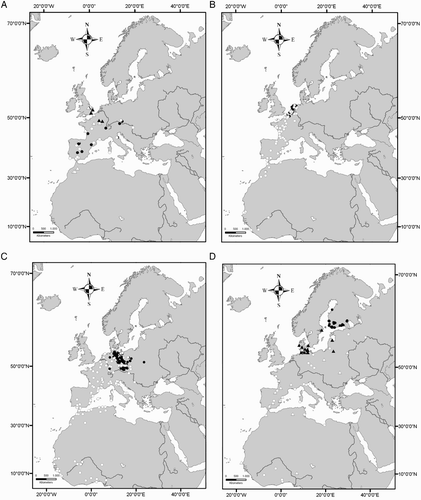
Figure 2. Migratory distances of male (n = 13) and female (n = 30) Eurasian Marsh Harriers. The bottom and top of boxes show the 25 and 75 percentiles, respectively. The horizontal line joined to the box by the dashed line shows the maximum and the minimum range of the data. Points are outliers.
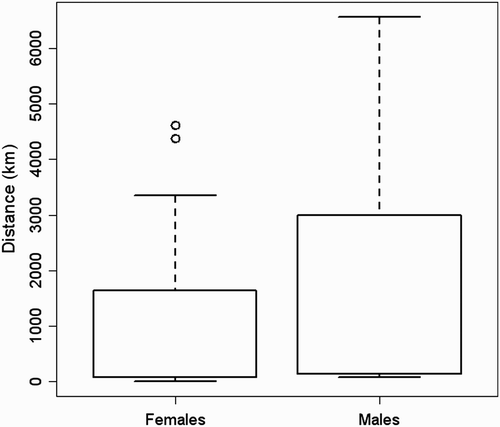
Figure 3. Ring and recovery sites of male (a) and female (b) Eurasian Marsh Harriers (black = ringing sites, white = recovery sites).
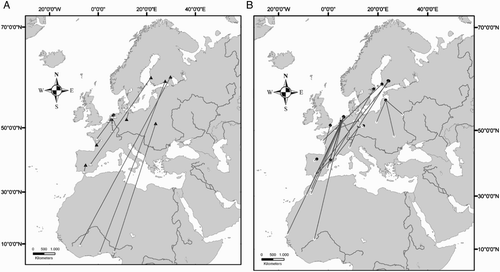
Figure 4. Longitudes of wintering sites located south of Sahara desert of juvenile (n = 20) and adult (n = 21) Eurasian Marsh Harriers. The horizontal bold lines show the median longitude of wintering site for each category. The bottom and top of boxes show the 25 and 75 percentiles, respectively. The horizontal line joined to the box by the dashed line shows the maximum and the minimum range of the data. Points are outliers.
DISCUSSION
Our results indicate that northern and eastern populations of Eurasian Marsh Harriers migrate further away from their breeding areas than southern and western ones. Starting from 49° of breeding latitude (), some individuals start to show very short migration distances or even residency. This migratory pattern could be largely explained by leapfrog migration that has been found in other systems (Alerstam Citation1990, Boland Citation1990, Marques et al. Citation2009, Kylin et al. Citation2011). Alerstam Citation(1990) gave two possible explanations of leapfrog migration. First, ‘competition and the timing of migration’: birds breeding in the southernmost part of the species range start and finish reproduction earlier than birds breeding in the northern areas. As a consequence, northern populations find the closest wintering areas are already occupied. For this reason they are forced to move further south. Secondly, ‘competition and the predictability in spring’: birds breeding in southern latitudes are more motivated to winter as close as possible to their breeding areas, within the same climatic region, in order to detect quickly the onset of spring and reach the breeding territories as soon as possible. On the other hand, northern breeders cannot take advantage of such a strategy because even overwintering in the first unoccupied wintering area means that they would always be outside the same climatic region of their breeding area. Therefore, they are forced to overshoot the first favourable wintering areas that they reach during migration, in order to avoid competition with southern breeders. Other hypotheses are based on optimal time allocation rather than effects of competition. Greenberg Citation(1980) showed that northern breeders, which have to spend the longest annual period away from the breeding range, may minimize their total mortality during the long non-breeding period by migrating to the wintering grounds furthest south. On the other hand, Bell (Citation1996, Citation1997) predicted that northern breeders with a late spring migration could utilize the surge of spring food at southern latitudes before starting the migration. All these assumptions are not mutually exclusive and could directly affect one another in influencing leapfrog migration in the Eurasian Marsh Harrier.
Figure 5. Graph showing the relationship between the lengths of the migratory journey in relation to the latitude of the breeding site. Latitude bands and sample size: up to 48.99°N = 13; 49.00–53.99°N = 261; 54.00–59.99°N = 20; 60.00°N and beyond = 26. The horizontal bold lines show the median migration distance of each latitudinal slot. The bottom and top of boxes show the 25 and 75 percentiles, respectively. The horizontal line joined to the box by the dashed line shows the maximum and the minimum range of the data. Points are outliers.
Marsh Harriers are strictly linked to wetlands that are mostly frozen during winter in north, central and eastern Europe, and therefore during this period feeding opportunities drastically decrease. The leapfrog pattern is more pronounced in individuals breeding in Eastern Europe compared to the western individuals probably because in Western Europe in winter there are more ice-free water bodies, probably resulting from the influence of the Atlantic Ocean. Individuals breeding in Western and Southern Europe can remain close to their breeding sites because wetlands do not freeze, especially on coastal areas (like western France). As a consequence Eurasian Marsh Harriers from northern and eastern Europe are forced to migrate further to avoid competition with these resident (or shorter-distance migrants) birds. This may entail costs, however. In many species of migrating birds, early arrival is associated with higher fitness (Myers Citation1981, Alerstam Citation1990). Eurasian Marsh Harriers breeding in Sweden that arrived later had lower breeding success and this was also linked to the difficulties in crossing ecological barriers such as the Sahara desert (Strandberg et al. Citation2009a). Migrants that have further to go may use several strategies to reduce the time needed to undertake their migratory journey (i.e. the use of more direct routes, limiting the use of stop-over sites and thus extending the time spent travelling during the night; see also Alerstam Citation2006).
Finally, it is important to consider the different starting dates for breeding: individuals with a northerly breeding distribution start their reproduction on average some weeks later than birds in western and southern Europe (Ferguson-Lee & Christie 2001, Génsbøl Citation2004). This pattern, with individuals coming from the eastern part of the breeding range moving further than the western ones, is also known for the eastern subspecies of Common Buzzard. Steppe Buzzards (Buteo buteo vulpinus) overwinter in Africa down to South Africa, while individuals belonging to the nominate subspecies remain mostly in Europe during winter (Alerstam Citation1990).
Considering the effect of the age-class, our GLM shows that there were no differences in migration distance among adults and juveniles. Satellite tracking data show that no juveniles from Sweden have wintered south of the Sahara, although the sample size (three individuals) was too small to draw any conclusion (Strandberg et al. Citation2008). More data on age proportions from different areas are needed to verify if there is a latitudinal segregation in relation to the age of the individuals.
Male Eurasian Marsh Harriers showed a higher mean migratory distance than females, but this result was not significant. This fits with the results of visual observations carried out in the Mediterranean basin, both at migration bottlenecks and at wintering sites, that showed a stronger tendency for adult male Eurasian Marsh Harriers to migrate over longer distances than adult females, because an imbalance towards males has been observed during migration en route and from Africa, with the opposite trend at wintering sites in Italy (Agostini & Logozzo Citation2000, Agostini et al. Citation2003, Panuccio et al. Citation2005a, Citation2005b, Citation2012, Agostini & Panuccio Citation2010). It has been suggested that this latitudinal segregation of the sexes is promoted by sexual dimorphism, because females, being larger, are more adept than smaller males in taking larger prey items and coping with colder temperature, on the basis of the Bergman's rule (Cristol et al. Citation1999, Panuccio et al. Citation2005a). As in other studies based on ringing recovery data, it is likely that the proportion of the population wintering in Africa is greater than that shown by our data set, because ring monitoring in Europe is more widespread compared to Africa (Bai & Schmidt Citation2012).
When considering individuals wintering south of Sahara desert, juveniles were distributed significantly more west during the winter than adults, even when accounting for the latitude of the breeding site. One possible explanation of this might be a differential response to the prevailing winds. Winds blowing towards west are dominant across the Sahara desert in autumn and this affects the migration paths of raptors, including Eurasian Marsh Harriers (Klaassen et al. Citation2010, Citation2011). In other species of raptors it has been observed that adults (experienced individuals) compensate for the drift of lateral winds during migration while juveniles (inexperienced individuals) do not (Thorup et al. Citation2003). This may occur because adults have better orientation abilities and are more skilled at compensating for wind drift. Alternatively, juveniles may be more susceptible to drifting in lateral winds because it is their first migration and they are without any specific winter site as a final destination prior to setting off. We suggest that juvenile Eurasian Marsh Harriers are more likely to be affected by the wind during their first flight across the Sahara desert and, as a result, their wintering sites are located more west than adults and their paths are differently oriented. It remains unclear if and how juveniles then shift their wintering grounds East during the subsequent winters. Juveniles could be more likely than adults to make extensive exploratory movements before they establish their wintering site (Alerstam Citation1990). However, within our data set, we found no differences in the dates of recoveries between juvenile and adult Eurasian Marsh Harriers and therefore we do not think that the exploratory behaviour of juveniles biased our analyses. An alternative explanation to the longitudinal differences could be that in coastal areas of Western Africa rice fields are widely distributed (Zwarts et al. Citation2009) and because juveniles are inexperienced in finding optimal and safe wintering sites it is possible that higher numbers of juveniles are killed in those sites, compared to areas located more inland (i.e. along the Niger River valley).
Our study confirms leapfrog migration patterns occur in Eurasian Marsh Harriers and suggests that migration distances vary by latitudinal and longitudinal gradients. Age-class did not influence the migratory distance while the slight difference among males and females suggests that further studies are recommended to investigate the hypothesis of partial sexual segregation during winter. Finally, this research confirms that is possible to perform large scale analyses on a large sample of individuals using ringing recovery data. This approach allows us to unravel differences in migration strategies and wintering locations among different age- and sex-classes that would be much more difficult, if not impossible, using satellite tracking techniques that currently have a very limited sample size.
ACKNOWLEDGEMENTS
We are grateful to Chris du Feu, EURING for the permission to use their data set. We also thank Will Cresswell and two anonymous reviewers for their comments on an earlier draft of the manuscript, and Jack Ashton-Booth for reviewing the English of the manuscript. Ugo Mellone is supported by FPU grant of the Spanish Ministry of Education (reference AP2008-0947). MEDRAPTORS (www.raptormigration.org) is a network of ornithologists and birdwatchers, and works to improve research and the protection of migrating birds of prey through specific projects and observation camps.
REFERENCES
- Agostini , N. , Coleiro , C. and Panuccio , M. 2003 . Autumn migration of Marsh Harriers across the central Mediterranean in 2002 . Ring , 25 : 47 – 52 . (doi:10.2478/v10050-008-0073-3)
- Agostini , N. and Logozzo , D. 2000 . Migration and wintering distribution of the Marsh Harrier (Circus aeruginosus) in southern Italy . Buteo , 11 : 19 – 24 .
- Agostini , N. and Panuccio , M. 2010 . Western Marsh harrier (Circus aeruginosus) migration through the Mediterranean sea: a review . J. Raptor Res. , 44 : 136 – 142 . (doi:10.3356/JRR-09-48.1)
- Alerstam , T. 1990 . Bird Migration , Cambridge , , UK : Cambridge University Press .
- Alerstam , T. 2006 . Strategies for the transition to breeding in time-selected bird migration . Ardea , 94 : 347 – 357 .
- Alerstam , T. and Hedenström , A. 1998 . The development of bird migration theory . J. Avian Biol. , 29 : 343 – 369 . (doi:10.2307/3677155)
- Bai , M. L. and Schmidt , D. 2012 . Differential migration by age and sex in central European Ospreys Pandion haliaetus . J. Ornithol. , 153 : 75 – 84 . (doi:10.1007/s10336-011-0697-y)
- Bavoux , C. , Burneleau , G. , Nicolau-Guillaumet , P. and Picard , M. 1992 . Le Busard des roseaux Circus a. aeruginosus en Charente Maritime. V – Déplacements et activité journalière des juvéniles en hiver . Alauda , 60 : 149 – 158 .
- Bavoux , C. , Burneleau , G. , Nicolau-Guillaumet , P. and Picard , M. 1994 . Le Busard des roseaux Circus a. aeruginosus en Charente-Maritime (France). VII – Déplacements et activité journalière des adultes en hiver . Alauda , 62 : 281 – 288 .
- Bell , C. P. 1996 . Seasonality and time allocation as causes of leapfrog migration in the Yellow Wagtail Motacilla flava . J. Avian Biol. , 27 : 334 – 342 . (doi:10.2307/3677265)
- Bell , C. P. 1997 . Leapfrog migration in the Fox Sparrow: minimizing the cost of spring migration . Condor , 99 : 470 – 477 . (doi:10.2307/1369953)
- Bildstein , K. L. 2006 . Migrating Raptors of the World , Ithaca : Cornell University Press .
- Boland , J. M. 1990 . Leapfrog migration in North American shorebirds: intra and interspecific examples . Condor , 92 : 284 – 290 . (doi:10.2307/1368226)
- Brown , L. H. , Urban , E. K. and Newman , K. 1982 . The Birds of Africa. Vol. 1: Ostriches to birds of prey , London : Academic Press .
- Cristol , D. A. , Mitchell , B. B. and Carbone , C. 1999 . “ Differential migration revisited: latitudinal segregation by age and sex class ” . In Current Ornithology Edited by: Nolan , V. Vol. 15: 33–38. Academic/Plenum Publisher, New York
- Ferguson-Lees , J. and Christie , D. A. 2001 . Raptors of the World , London : Christopher Helm .
- Fowler , J. and Cohen , L. 1992 . Statistics for Ornithologists , UK : BTO .
- Génsbøl , B. 2004 . Birds of Prey , 4 , London : Collins .
- Greenberg , R. 1980 . “ Demographic aspects of long-distance migration ” . In Migrant Birds in the Neotropics , Edited by: Keast , A. and Morton , E. S. 493 – 504 . Washington , DC : Smithsonian Institution Press .
- Klaassen , R. H.G. , Strandberg , R. , Hake , M. , Olofsson , P. , Tøttrup , A. and Alerstam , T. 2010 . Loop migration in adult Marsh Harriers Circus aeruginosus, as revealed by satellite telemetry . J. Avian Biol. , 41 : 200 – 207 . (doi:10.1111/j.1600-048X.2010.05058.x)
- Klaassen , R. H.G. , Hake , M. , Strandberg , R. and Alerstam , T. 2011 . Geographical and temporal flexibility in the response to crosswinds by migrating raptors . Proc. R. Soc. B. , 278 : 1339 – 1346 . (doi:10.1098/rspb.2010.2106)
- Kylin , H. , Bouwman , H. and Louette , M. 2011 . Distribution of the subspecies of Lesser Black-backed Gulls Larus fuscus in sub-Saharan Africa . Bird Study , 58 : 186 – 192 . (doi:10.1080/00063657.2011.555517)
- Marques , P. A.M. , Costa , A. M. , Rock , P. and Jorge , P. E. 2009 . Age-related migration patterns in Larus fuscus spp . Acta Ethol , doi: 10.1007/s10211-009-0060-y
- Myers , J. P. 1981 . A test of three hypotheses for latitudinal segregation of the sexes in wintering birds . Can. J. Zool. , 59 : 1527 – 1534 . (doi:10.1139/z81-207)
- Newton, I. 2008. Migration Ecology of Birds. Academic Press, London.
- Panuccio , M. , D'Amicis , B. , Canale , E. and Roccella , A. 2005a . Sex and age ratios of Marsh Harriers Circus aeruginosus wintering in central–southern Italy . Avocetta , 29 : 13 – 17 .
- Panuccio , M. , Agostini , N. and Mellone , U. 2005b . Autumn migration strategies of Honey Buzzards, Black Kites, Marsh and Montagu's Harriers over land and over water in the central Mediterranean . Avocetta , 29 : 27 – 32 .
- Panuccio , M. , Chiatante , G. and Tarini , D. 2013 . Two different migration strategies in response to an ecological barrier: Western Marsh Harriers and juvenile European Honey Buzzards crossing the central–eastern Mediterranean in autumn . J. Biol. Res.-Thessalon. , in press
- Salomonsen , F. 1955 . The evolutionary significance of bird migration . Dan. Biol. Medd. , 22 ( 6 )
- Smith , R. B. , Meehan , T. D. and Wolf , B. O. 2003 . Assessing migration patterns of Sharp-shinned Hawks Accipiter striatus using stable-isotope and band encounter analysis . J. Avian. Biol , 34 : 387 – 392 . (doi:10.1111/j.0908-8857.2003.03107.x)
- Sternalski , A. , Bavoux , C. , Burneleau , G. and Bretagnolle , V. 2008 . Philopatry and natal dispersal in a sedentary population of western marsh harrier . J. Zool. , 274 : 188 – 197 . (doi:10.1111/j.1469-7998.2007.00372.x)
- Strandberg , R. , Klaassen , R. H.G. , Hake , M. , Olofsson , P. , Thorup , K. and Alerstam , T. 2008 . Complex timing of Marsh Harrier Circus aeruginosus migration due to pre and post migratory movements . Ardea , 96 : 159 – 171 . (doi:10.5253/078.096.0202)
- Strandberg , R. , Klaassen , R. H.G. , Hake , M. and Alerstam , T. 2009a . How hazardous is the Sahara Desert crossing for migratory birds? Indications from satellite tracking of raptors . Biol. Lett. , 6 : 297 – 300 . (doi:10.1098/rsbl.2009.0785)
- Strandberg , R. , Klaassen , R. H.G. and Thorup , K. 2009b . Spatio-temporal distribution of migrating raptors: a comparison of ringing and satellite tracking . J. Avian Biol. , 40 : 500 – 510 . (doi:10.1111/j.1600-048X.2008.04571.x)
- Thorup , K. , Alerstam , T. , Hake , M. and Kjellén , N. 2003 . Bird orientation: compensation for wind drift in migrating raptors is age dependent . Proc. R. Soc. Lond. , series B 270: S8–S11
- Venables , W. N. and Ripley , B. D. 2002 . Modern Applied Statistics with R , 4 , New York : Springer .
- Wallin , K. , Wallin , M. L. , Jaras , T. and Standvick , P. 1987 . Leapfrog migration in the Swedish Kestrel Falco tinnunculus population . Proc. 5th Nordic Ornithol. Congr. , 1985 : 213 – 222 .
- Zwarts , L. , Bijlsma , R. G. , van der Kamp , J. and Wymenga , E. 2009 . Living on the Edge: Wetlands and Birds in a Changing Sahel , Zeist : KNNV Publishing .