Abstract
Capsule Red-necked Nightjars' (Caprimulgus ruficollis) choice of road surface type is significantly affected by air temperature, which may lead to increased traffic-induced mortality under cool weather conditions.
Aims To assess how seasonality and weather conditions affect road surface use by Red-necked Nightjars and, if so, to identify possible patterns in human-induced mortality as a result of Nightjars' decisions.
Methods In 2009–2011, the number of sitting birds were counted along gravel and paved roads in southwestern Spain to evaluate environmental correlates of surface use by Nightjars. Thermal benefits from using each surface type were determined based on local weather data, while seasonality in road casualty rates was assessed at the regional scale.
Results Nightjars' occurrence on roads varied seasonally with the nature of the surface. Nightjars' decisions were significantly affected by the thermal regime, with birds being attracted towards the warmth of paved roads during migration, when cool weather (< 20°C) usually prevailed, or at low temperatures (< 14°C). Paved roads provided birds with a significantly warmer substrate than nearby gravelled or sandy areas. However, traffic-induced mortality significantly increased when surface use shifted from gravel to paved roads.
Conclusion Plasticity in surface choice presumably helps Nightjars to minimize thermoregulatory costs during adverse weather conditions, although the adaptive value of thermal-related decisions appears to incur mortality costs for Nightjar populations.
Birds generally adjust foraging patterns based on physiological sensitivity to microclimate to reduce energy expenditure in thermally stressful environments (Grubb Citation1975, Walsberg Citation1985, Wolf Citation2000). Short-term responses to microclimate may be of particular importance after migratory movements, when birds may face unexpected energy deficits at their destination due to adverse weather conditions or reduced food supply (Dawson & Hudson Citation1970, Ashdown & McKechnie Citation2008).
Nightjars (Caprimulgiformes) are nocturnal and crepuscular aerial insectivores for which some aspects of behavioural responses to microclimate have been widely explored. While facultative heterothermy appears to be a widespread among caprimulgids to reduce energy demand in cold weather, only a few species have been shown to behaviourally thermoregulate (i.e. to selectively use microhabitats and microclimates; Hickey Citation1993, Poulin et al. Citation1999, Fisher et al. Citation2004, Doucette et al. Citation2011).
Red-necked Nightjars (Caprimulgus ruficollis, average body mass = 96.3 g, C. Camacho unpubl. data), hereafter called ‘Nightjars’, are long-distance migrants that breed in Mediterranean open scrublands of the Iberian Peninsula and northwest Africa (Holyoak Citation2001). From dusk to dawn, Nightjars often forage on roads, hawking insects during short flights or taking prey on the ground (Jackson Citation2003c). Foraging style and selection of foraging site are known to strongly influence individuals' heat balance in adverse weather conditions (Hickey Citation1993, Wolf & Walsberg Citation1996). However, thermal-related effects on site choice by Nightjars at the road network scale seem to have been largely unstudied.
Because of increased traffic volume in recent decades, road collisions have emerged as a major source of mortality for numerous caprimulgid species (Jackson Citation2002). Nevertheless, there are a few ways that roads can still benefit Nightjars. For example, Jackson Citation(2003b) suggested that Nightjars probably use roads to forage for flying insects that have been attracted there. Open road surfaces may facilitate detection of flying insects silhouetted against the sky (Jackson Citation2003a), and roadside vegetation might facilitate their effective escape when attacked (Camacho 2012). In addition, the heat stored during the day that is radiated at night (Trombulak & Frissell Citation2000) could moderate fluctuations in the availability of insect prey as air temperature drops or even reduce metabolic costs for birds resting on road surfaces (Whitford Citation1985). However, given that numerous interacting parameters can produce a wide range of road surface temperatures across a road network (Shao et al. Citation1997), density of aerial insect prey and associated Nightjar occurrence might also vary among different road types in response to seasonality or weather conditions. Like some congeners (see Jackson Citation2003c), Red-necked Nightjars are attracted to both gravel and paved roads (Camacho Citation2012). However, mortality rates are usually higher on paved roads (Jackson Citation2002), which may counteract any potential thermal benefits in poor weather conditions. The main aim of this study was to assess how seasonality and weather conditions affect surface use by road-sitting Nightjars and to identify possible spatio-temporal patterns in human-induced mortality as a result of Nightjars' choice for different road types.
If Nightjars' occurrence on roads is determined by physiological thermal preferences, then the species' choice of road attributes in temperate regions should be seasonally affected by phenology or weather conditions. On the other hand, it might be predicted that high bird occurrences on paved roads could then lead to locally increased mortality risk due to the increased traffic volume and speed relative to unpaved roads. To address both hypotheses, I first analysed the effect of seasonality and weather variables on road use by Red-necked Nightjars during their time on the breeding grounds. Then, I explored the role of variation in road use at the study site on traffic-induced mortality at the regional scale.
METHODS
Study area
Selection of foraging sites by Nightjars was studied along roads surrounding the northwestern border of Doñana National Park in southwestern Spain (37°7′N, 6°33′W). The study plot consisted of a 24-km road circuit of gravel (17 km) and paved (7 km) roads crossing a private hunting reserve. This flat sandy area is partially covered by a mixture of Mediterranean scrublands with cattle-grazed grasslands and pine tree plantations, while the road verge plant community consisted of sparse tall (> 2 m) and medium height (1–2 m) shrubs and numerous dwarf stands and weeds. Access to the hunting reserve is highly restricted, so that traffic volume is considerably reduced (< 10 vehicles/24 h). To avoid possible bias in estimates of surface choice by Nightjars due to relocating birds disturbed by vehicles, roads with low traffic density were required. For this reason, together with the high occurrence of road-sitting Nightjars (up to 3 birds/km) and regular road widths (3.5 − 4 m), this road circuit was selected to assess the spatial responses of birds to microclimate.
Road occurrence and surface choice
During August–November 2009 and 2010 and March–November 2011, weekly transect counts of Nightjars were conducted along the 24-km road circuit. To account for a possible hourly variation in use of roads, all nocturnal censuses began 1–2 h after dusk from fixed starting points by driving a vehicle at a constant speed of 30 km/h until the full route was covered in both directions (2–3 h). Weekly counts were undertaken on lightly (< 30%) clouded nights to avoid a possible effect of variable light conditions on bird detection (Wilson & Watts Citation2006). Censuses were also limited to nights with no precipitation and low (< 10 km/h) wind velocity. The occurrence of Nightjars in each stretch (paved or gravelled) was recorded by detecting their eye shine from afar with headlights on full beam (brightest), as nearly all of the Nightjars (> 95%) face away from the road side (Camacho Citation2012). To assess the spatio-temporal variation in bird occurrence on roads (i.e. surface use and phenology), Nightjar numbers on each stretch were standardized to Kilometric Indexes of Abundance (KIA, Tellería Citation1986). The ratio between Nightjars' occurrences on gravel and paved roads (KIA gravel/KIA paved) provided a simple direct measure of their use of each surface type for each census.
To account for a potential effect of adjacent habitat features on Nightjars' choice of road type, vegetation height of the road verge community was measured (to the nearest 1 cm) next to paved and gravel roads at 30 and 50 random sites, respectively. To assess the environmental correlates of Nightjar use of surface types, I considered both the major factors influencing caprimulgid activity, such as air temperature and light availability (Jetz et al. Citation2003, Ashdown & McKechnie Citation2008) and other variables potentially affecting surface choice, such as rainfall and relative air humidity. To assess the light regime, moon phases were converted to the fraction of moon disk illuminated by considering the gradual increase from 0% (new moon) to 100% (full moon) throughout a complete lunar cycle. Thermal differences between gravel and paved roads and adjacent (w 3 m) ground (i.e. potential benefits from using each road type) were calculated at 30 randomly determined points along the 24-km road circuit. Differences in surface temperature between substrate types during cold periods (air temperature <14°C) were determined (to the nearest 0.1°C) during two consecutive nights (11 and 12 October 2011; minimum air temperature = 13.2°C and 11.3°C, respectively) by using a digital thermometer with sensor probe (Digi-ThermoTM WT2). Paired measurements (n = 15 for each of the two road surfaces) were simultaneously taken between 22:00 and 2:00 h on both adjoining surfaces (road centre and bare ground 1–3 m away from road) to control for possible effects of time differences and to avoid possible bias due to differences in amount of surrounding vegetation. Rainfall, relative humidity and temperature data were taken from the Manecorro RM1 meteorological station (http://icts.ebd.csic.es) located inside Doñana National Park, 2.8 km from the study site.
Road casualties
Because of the low traffic density (and associated road casualties) in the study area, there were no data available on mortality differences between sampled gravel and paved roads. For this reason, together with similarities in climate, a large data set from nine institutional wildlife recovery centres (Environment Agency of the Andalusian Regional Government, 2009–2011) spread across Andalusia (southern Spain) was used to assess seasonality in road casualties at the regional scale. Numerous injured or dead Nightjars are annually taken into wildlife recovery centres. Here, I assume that increased road fatalities would lead to an equivalent increase in the number of birds entering the centres, so that the monthly records of road incidents of Red-necked Nightjars would provide a coarse measure of seasonality in traffic-induced mortality. Thus, road casualty rates were estimated as the number of casualties of vehicle collision that entered the recovery centres per week. Taking Nightjar use of roads at the local scale as a proxy for the whole region, broad-scale data on road casualties were grouped into two periods for analyses (see ‘results’): June–September and April–May and October.
Statistical analyses
All analyses were performed using General Linear Models (GLM), unless otherwise stated. To evaluate the environmental correlates of Nightjars' occurrence (KIA) on roads, a GLM with normal distribution for errors and an identity link function was fitted, including the ratio between Nightjars' occurrence on each surface (i.e. KIA on paved/KIA on gravel roads) as the response variable (previously square-root transformed to meet assumptions of normality). Moonlight and relative environmental humidity were included as covariates, whereas rainfall occurrence in the previous 36 h was included as a fixed factor. Vegetation height along the road verges did not differ between gravel and paved roads (GLM with Poisson distribution for errors: F 1,79 = 1.59, P = 0.21), so a potential effect of landscape features (e.g. different shading of the road surfaces) on Nightjar use of road type was not considered further. To explore the differences in surface temperature between roads and nearby unpaved grounds, a Wilcoxon signed rank test (Z) was used, whereas a chi-square test (χ 2) was applied to explore the seasonal differences in traffic-induced casualties. The software package R, v. 2.15 (R Development Core Team 2012) was used for statistical analyses. Differences were considered to be significant when P < 0.05. All values are reported as means ± se.
RESULTS
From 2009 to 2011, 1539 observations of road-sitting Nightjars were collected. Nightjars arrived at the study area in early April and left in late October (first and last record on 3 April and 22 October, respectively).
Seasonality in road–Nightjar association
Red-necked Nightjars readily settled on roads throughout the time they resided in the study area. However, birds' occurrence and, accordingly, potential mortality risk due to roadkill, varied seasonally due to migratory behaviour and breeding activities (). The number of road-sitting individuals notably increased from early April to mid May, coincident with the passage and arrival of the latest migrants co-occurring with the earliest breeders. The intensity of Nightjar use of roads fluctuated widely as the breeding season progressed (90% females laying between 16 and 30 June in the study site; C. Camacho, submitted MS). Occurrence on roads finally increased during postnuptial peak of passage in the second half of September, followed by a steep decrease with birds completely disappearing by late October.
Table 1. Seasonal variation in Red-necked Nightjars' occurrence on the road network crossing the study area (paved and gravel roads together), with number of road casualties at the regional scale (which represents the proportion of all casualties that occurred during each month) and mean daily air temperatures at the study site. Values are reported as means ± se.
Road surface use
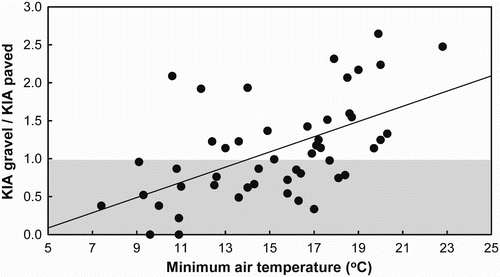
The highest bird abundances alternated throughout the season according to the nature of the road surface. Whereas Nightjars' occurrence on gravel roads gradually increased during the breeding season and early postnuptial migration (June–September), most Nightjars concentrated on paved roads after arrival from their wintering grounds in April–May and immediately before late departure in October (). Analysis of environmental cues potentially affecting Nightjar use of roads indicated there was no effect of rainfall, relative environmental humidity or moon phase, although air temperature explained most of the variation in abundance of Nightjars on roads (), with birds attracted to paved roads mainly on cold nights (i.e. air temperature < 14°C; see ). After absorbing heat during the day, both gravel and paved roads were significantly warmer than adjoining unpaved ground (Wilcoxon signed-rank test, Z = 351, P < 0.0001, n = 30 paired measures). However, the temperature gain for birds on paved roads in relation to nearby bare sandy areas or leaf litter (+4.45 ± 0.61°C) was significantly higher than that for gravel roads (+2.91 ± 0.40°C; GLM with Poisson distribution for errors: F 1,29 = 4.1, P = 0.046).
Table 2. Multiple regression model (GLM with normal distribution for errors and an identity link function) testing the effect of environmental cues on road surface use by Red-necked Nightjars. The ratio between birds/km on gravel and paved roads is included as the dependent variable. Significant values (P < 0.05) are highlighted in bold.
Figure 1. Seasonal variation in road surface use by Red-necked Nightjars at the study site. Bars show the relative occurrence (with standard errors of the mean) of birds on gravel and paved roads. Values >1 and <1 indicate Nightjars concentrating on gravel and paved roads, respectively.
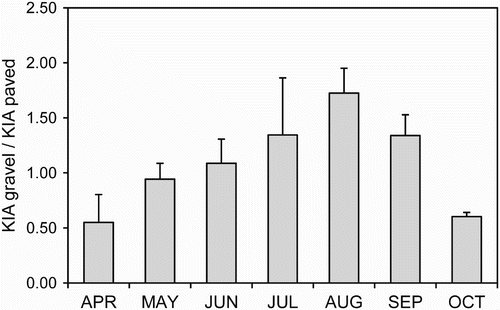
Figure 2. Effect of air temperature on surface use by Red-necked Nightjars at the study site. Shaded and open areas illustrate the use of paved and gravel roads, respectively.
Road casualty rates
Traffic-induced casualties occurred throughout most of the period that Nightjars were in southern Spain (n = 309, April–November). However, vulnerability to road traffic exhibited a seasonal pattern. Most casualties were recorded from April to May (46.6%), even though bird numbers on either road type were highest in June and August (). Road casualty rates during the periods in which Nightjars concentrated on paved roads (April–May and October; 5.1 ± 0.8 casualties/week; ) were almost double those when most birds occurred on gravel roads (June–September; 2.7 ± 0.58 casualties/week). Thus, the number of road casualties were significantly highest during the coldest months ((X 2 1 = 6.20, P = 0.013), April–May and October, ), when Red-necked Nightjars were probably attracted to the warmth of paved roads.
DISCUSSION
Road surface use
Red-necked Nightjars settled on both gravel and paved roads, although their use of surface types changed with the local thermal regime, with more birds occurring on paved roads when air temperature dropped below about 14°C. Nightjar behaviour can be significantly influenced by weather conditions (Ashdown & McKechnie Citation2008), which likely reflects the negative effect of low temperatures on aerial insect density (Jetz et al. 2003). In seasonal environments in South Africa, where aerial insects are a highly variable food resource for consumers, Ashdown & McKechnie Citation(2008) found a complete cessation of the foraging activity of Freckled Nightjars (Caprimulgus tristigma) at air temperature < 12°C. This thermal threshold is strikingly close to that reported for other insectivorous bird species in higher more seasonal latitudes. For example, in northern Norway, Speakman et al. (Citation2000) showed that aerial insect density decreased rapidly at air temperature < 15°C, which often led Sand Martins (Riparia riparia) to stop feeding. At my study site, although patterns of foraging activity were not evaluated, the use of foraging microhabitats was significantly influenced by a rather similar threshold (air temperature < 14°C). Foraging efficiency might partially explain the Nightjar use of paved roads at low air temperature, when drastic shifts in prey availability could be moderated by artificial heat sources. Throughout the Afrotropical region, nightjars have been traditionally thought to occur on roads to forage for flying insects that have been attracted there (Jackson Citation2003b). However, it is not well known whether insect densities are similarly higher in adjacent open areas (Jackson Citation2003a). Therefore, increased prey availability may not necessarily be the sole predictor of the occurrence of Red-necked Nightjars on different road types.
On the other hand, ambient light regime could provide an alternative explanation for the dual surface choice by Nightjars. Mills Citation(1986) first suggested that high illumination may counteract the adverse effects of low prey density to Caprimulgus species by improving their visual hunting abilities and hence increasing their foraging efficiency. In equatorial West Africa, Jetz et al. Citation(2003) noted that foraging activity of Long-tailed Nightjars (Caprimulgus climacurus) increased significantly around the new moon, which seems to also be the case in Freckled Nightjars inhabiting seasonal subtropical environments (Ashdown & McKechnie Citation2008). Nevertheless, I found no effect of light regime on birds' selection of where to forage, suggesting that moonlight-enhanced foraging does not appear to influence the variation in surface use of Red-necked Nightjars reported in this study.
Behavioural thermoregulation
Nocturnal insectivores inhabiting cold seasonal environments must often face energetic challenges when density of aerial insects drops (see above). In adverse weather, energy requirements may sometimes be greater than the energy that can be acquired through foraging, which might increase the likelihood that Nightjars will face occasional energy deficits (Dawson & Hudson Citation1970). However, mismatches between energy requirement and food supply could be partially moderated by reducing energy demand.
Torpor use is a widespread strategy among caprimulgids to reduce energy expenditure (Brigham et al. Citation2012). However, propensity to use of facultative heterothermy in cold weather not only varies among the different species of Caprimulgidae (Brigham et al. 2012), but also their ability to enter torpor may influence foraging behaviour (Hickey Citation1993). Possibly, Red-necked Nightjars are incapable of entering torpor or, as reported for Whip-Poor-Wills (Caprimulgus vociferans; Lane et al. Citation2004), their frequency of torpor use is lower than for other species. If that is the case for Red-necked Nightjars, then plasticity in surface choice might replace heterothermy to overcome poor weather conditions.
Behavioural thermoregulation may be an efficient alternative to metabolic thermoregulation among caprimulgids. Whip-Poor-Wills, for example, employ a low-cost foraging style (i.e. sit-and-wait) that allows them to forage during periods of low prey availability (Hickey Citation1993). In the Red-necked Nightjar, the selective use of foraging microclimates might allow them to lower their energy requirements to maintain an elevated body temperature. Choice of foraging site may help some birds to regulate their heat balance with almost no energy expenditure (Wolf & Walsberg Citation1996, Wolf Citation2000). As a result, individuals generally adjust foraging behaviour according to physiological sensitivity to microclimate (Walsberg Citation1985) to conserve energy in thermally stressful environments (Grubb Citation1975). Surface choice by Red-necked Nightjars was strongly correlated with air temperature and partially influenced by the migratory phenology of the species, with most birds using paved roads in months in which mean temperatures were ≤ 20°C (). During migration, which usually occurs from early April and late October, weather conditions in southern Europe might still be too cool (mean air temperature ≤ 20°C) for this Afrotropical species. High surface temperature in paved roads likely helps Nightjars to overcome the exposure to low air temperature. Alternatively, during nights in which air temperature never fell below 20–25°C, paved roads could be sometimes too hot (surface temperature 35°C, data from anecdotal measurements in August 2011) for the birds to use early in the night (when bird counts were conducted). Overheating might have led to increased Nightjar use of gravelled roads during midsummer. Based on data from numerous Afrotropical nightjars, Jackson Citation(2003c) noted that species frequent preferred substrates for foraging, but will frequently be seen in other areas as well. Accordingly, plasticity in surface choice appears to be widespread among caprimulgids, although there is no clear reason for the variation in surface use in tropical regions. In summary, choice of roads for foraging sites by Red-necked Nightjars may present an efficient strategy to maintain a high euthermic body temperature during adverse weather conditions or energetic shortfalls by them selecting paved stretches of road with a noticeably warmer surface than the nearby bare ground.
Implications for conservation
The apparently high adaptive value of thermal-related movements could concurrently involve some negative effects for Nightjars in man-made landscapes. Birds' choice of paved roads during migration (especially in late May) might increase Nightjars' vulnerability to road traffic. This is consistent with the results from Jackson Citation(2002), who noted that migrants, in particular, seemed to suffer the highest road mortality.
Greater traffic density usually means greater mortality. Road-settled Nightjars are vulnerable to any motor vehicle travelling between sunset and sunrise, being liable to be killed even by vehicles travelling at slow speeds (Jackson Citation2002). Traffic density and speed on paved roads are usually high, so birds do not have enough time to avoid oncoming traffic (Jackson Citation2002). Moreover, in contrast to gravel roads, approaching vehicles on paved roads are quieter. Nevertheless, despite high casualty rates, Nightjars continue to frequent all types of roads throughout their breeding range. I conclude that mortality risk due to roadkill may be partially affected by seasonality and directly influenced by environmental cues determining birds' occurrence on roads. As a result, behavioural thermoregulation might be a human-induced trap, because natural selection should favour both Nightjars that avoid highly busy (paved) roads and those that actively select warm (also paved) surfaces during cold weather.
ACKNOWLEDGEMENTS
SEO/Birdlife partially supported my work through a Research Initiation Grant. The Environment Agency of the Andalusian Regional Government kindly provided their records of road casualties. J.L. del Valle and L. García thoroughly collaborated during data collection, whereas S. Sánchez, P. Villar and P. Sáez helped at different stages of this study. I especially thank M. Camacho, J. Potti, W. Cresswell, R.M. Brigham and an anonymous referee for valuable comments that considerably improved the manuscript. The author declares that he has no conflict of interests.
REFERENCES
- Ashdown , R. A.M. and Mckechnie , A. E. 2008 . Environmental correlates of Freckled Nightjar (Caprimulgus tristigma) activity in a seasonal, subtropical habitat . J. Ornithol. , 149 : 615 – 619 . (doi:10.1007/s10336-008-0309-7)
- Brigham , R. M. , McKechnie , A. E. , Doucette , L. I. and Geiser , F. 2012 . “ Heterothermy in caprimulgid birds: a review of inter- and intraspecific variation in free-ranging populations ” . In Living in a Seasonal World Edited by: Ruf , T. , Bieber , C. , Arnold , W. and Millesi , E. 175 – 1872 . DOI: 10.1007/978-3-642-28678-0_16
- Camacho , C. 2012 . “ Plantas ‘guardaespaldas’: microhábitat de caza en el Chotacabras cuellirrojo Caprimulgus ruficollis ” . In XIV Congreso Nacional y XI Iberoamericano de Etología. Libro de resúmenes Edited by: Redondo , T. 23. Sevilla. http://digital.csic.es/handle/10261/54769
- Dawson , W. R. and Hudson , J. W. 1970 . “ Birds ” . In Comparative Physiology of Thermoregulation , Edited by: Whittow , G. C. New York : Academic Press .
- Doucette , L. I. , Brigham , R. M. , Pavey , C. R. and Geiser , F. 2011 . Roost type influences torpor use by Australian owlet-nightjars . Naturwissenschaften , 98 : 845 – 854 . (doi:10.1007/s00114-011-0835-7)
- Fisher , R. J. , Fletcher , Q. E. , Willis , C. K.R. and Brigham , R. M. 2004 . Roost selection and roosting behaviour of male Common Nighthawks . Am. Midl. Nat. , 151 : 79 – 87 . (doi:10.1674/0003-0031(2004)151[0079:RSARBO]2.0.CO;2)
- Grubb , T. C. 1975 . Weather-dependent foraging behavior of some birds wintering in a deciduous woodland . Condor , 77 : 175 – 182 . (doi:10.2307/1365788)
- Hickey , M. B.C. 1993 . Thermoregulation in free-ranging Whip-Poor-Wills . Condor , 95 : 744 – 747 . (doi:10.2307/1369625)
- Holyoak , D. T. 2001 . Nightjars and Their Allies: The Caprimulgiformes , Oxford : Oxford University Press .
- Jackson , H. D. 2002 . A review of Afrotropical Nightjar mortality, mainly road kills . Ostrich , 73 : 147 – 161 .
- Jackson , H. D. 2003a . A field survey to investigate why Nightjars frequent roads at night . Ostrich , 74 : 97 – 101 . (doi:10.2989/00306520309485374)
- Jackson , H. D. 2003b . Another reason for Nightjars being attracted to roads at night . Ostrich , 74 : 228 – 230 . (doi:10.2989/00306520309485398)
- Jackson , H. D. 2003c . A review of foraging and feeding behaviour, and associated anatomical adaptations, in Afrotropical Nightjars . Ostrich , 74 : 187 – 204 . (doi:10.2989/00306520309485392)
- Jetz , W. , Steffen , J. and Linsenmair , K. E. 2003 . Effects of light and prey availability on nocturnal, lunar and seasonal activity of tropical Nightjars . Oikos , 103 : 627 – 639 . (doi:10.1034/j.1600-0706.2003.12856.x)
- Lane , J. E. , Brigham , R. M. and Swanson , D. L. 2004 . Daily torpor in free-ranging whip-poor-wills (Caprimulgus vociferus) . Physiol. Biochem. Zool. , 77 : 297 – 304 . (doi:10.1086/380210)
- Mills , A. M. 1986 . The influence of moonlight on the behavior of Goatsuckers (Caprimulgidae) . Auk , 103 : 370 – 378 .
- Poulin , R. G. , Todd , L. D. and Brigham , R. M. 1999 . Male common nighthawk use of gravel roads at night . Prairie Naturalist , 30 : 85 – 90 .
- R Development Core Team . 2010 . R: A Language and Environment for Statistical Computing , R Foundation for Statistical Computing, Vienna. http://www.R-project.org
- Shao , J. , Swanson , J. C. , Patterson , R. , Lister , P. J. and McDonald , A. N. 1997 . Variation of winter road surface temperature due to topography and application of thermal mapping . Meteorol. Appl. , 4 : 131 – 137 . (doi:10.1017/S135048279700042X)
- Speakman , J. R. , Rydell , J. , Webb , P. I. , Hayes , J. P. , Hays , G. C. , Hulbert , I. A.R. and McDevitt , R. M. 2000 . Activity patterns of insectivorous bats and birds in northern Scandinavia (69°N), during continuous midsummer daylight . Oikos , 88 : 75 – 86 . (doi:10.1034/j.1600-0706.2000.880109.x)
- Tellería , J. L. 1986 . Manual para el censo de los vertebrados terrestres , Madrid : Raices .
- Trombulak , S. C. and Frissell , C. A. 2000 . Review of ecological effects of roads on terrestrial and aquatic communities . Conserv. Biol. , 14 : 18 – 30 . (doi:10.1046/j.1523-1739.2000.99084.x)
- Walsberg , G. E. 1985 . “ Physiological consequences of microhabitat selection ” . In Habitat Selection in birds , Edited by: Cody , M. L. 389 – 413 . Orlando , Florida : Academic Press .
- Wilson , M. D. and Watts , B. D. 2006 . Effect of moonlight on detection of Whip-poor-wills: implications for long-term monitoring strategies . J. Field Ornithol. , 77 : 207 – 211 . (doi:10.1111/j.1557-9263.2006.00042.x)
- Whitford , P. C. 1985 . Bird behavior in response to the warmth of blacktop roads . Wisconsin Acad. Sci., Arts and Lett. , 73 : 135 – 143 .
- Wolf , B. O. 2000 . Global warming and avian occupancy of hot deserts; a physiological and behavioral perspective . Rev. Chil. Hist. Nat. , 73 : 395 – 400 . (doi:10.4067/S0716-078X2000000300003)
- Wolf , B. O. and Walsberg , G. E. 1996 . Thermal effects of radiation and wind on a small bird and implications for microsite selection . Ecology , 77 : 2228 – 2236 . (doi:10.2307/2265716)