Abstract
Capsule Black Guillemots Cepphus grylle were recorded diving in the Pentland Firth to an average depth of 32 metres and an overall maximal depth of 43 metres. The majority (88%) of dives were benthic with a median dive duration of 95 seconds, and a maximal dive duration of 131 seconds. The results provide empirical evidence that Black Guillemots use depths within the water column at which tidal turbines are likely to operate. Although limited, our data suggest the potential for interactions between Black Guillemots and marine renewable energy devices.
The Scottish Government has a target to produce 100% of electricity demand from renewable energy by 2020 (The Scottish Government Citation2011) and 11 sites in the Pentland Firth and Orkney waters have been leased for the development of wave or tidal renewable energy devices. The environmental effects of these devices are largely unknown (Shields et al. Citation2011), but it is likely that impacts will be species-specific. The Black Guillemot (Cepphus grylle, Linnaeus 1758) is known to preferentially forage in moderate- to high-flow environments (Nol & Gaskin Citation1987) and is present in northern regions. It is predominantly a benthic forager and has been reported to dive to up to 50 metres (Piatt & Nettleship Citation1985, Cairns Citation1992). These are depths at which tidal turbines are likely to operate, making this species potentially susceptible to tidal energy developments (Furness et al. Citation2012), but a lack of data makes it difficult to assess the risk posed. Due to the rapid development of the marine renewable energy industry in Scotland, particularly tidal stream turbines, in areas of Black Guillemot habitat, there is an urgent need for ecological data which will inform planning decisions. The Black Guillemot has also been included on the marine protected area (MPA) search feature list for Scottish territorial waters because the species currently experiences protection only from a small number of Sites of Special Scientific Interest. A lack of ecological data will limit the appropriate designation of these MPAs. The aim of this study was to collect empirical data on the diving behaviour of Black Guillemots in an area where tidal energy developments are planned, and more generally enhance our ecological understanding of this species in Scotland.
The study was conducted from the island of Stroma, Caithness (58°41′8″N 03°07′0″W) located in the north of Scotland, UK. The island is surrounded by the Pentland Firth, with the Inner Sound to the south. The Inner Sound, with maximal water depths of around 35 metres and current velocities reaching 4.5 ms−1 (Easton et al. Citation2011), is one of 11 sites being leased by The Crown Estate for marine renewable energy development. To study Black Guillemot diving behaviour, we used Cefas G5 time-depth recorders (depth loggers; diameter = 11.5 mm, length = 35.5 mm, mass = 5.7 g) programmed to record depth (with an accuracy of approximately 1 metre) via pressure measurement at 1 second intervals. Devices were deployed on five birds between the 7th and 12th July 2011. One bird was caught by hand on a clutch of two eggs and four others were caught in mist-nets set in front of the colony. Upon capture, each bird was ringed and weighed. A depth logger was then attached to the body feathers of each bird using black waterproof tape (Tesa SE, Hamburg, Germany), which allowed for the loss of instruments through feather moult should the bird not be recaptured (Thaxter et al. Citation2010). Two birds had the device attached dorsally, the other three were ventral (Tremblay et al. Citation2003). The mean mass of the birds handled was 359 g (sd = 19) and the mean handling time per bird was 11 minutes (sd = 4). After two to three days, attempts were made to recapture the birds and the devices removed.
Data from depth loggers were analysed using r version 3.0.1 (R Core Team Citation2013) and the open-source package divemove (Luque Citation2007, Luque & Fried Citation2011) to determine the depth and duration of all dives deeper than 2 metres. Dives shallower than 2 metres likely occurred during activities other than foraging, for example evasion of kleptoparasitic gulls, so were excluded from the analysis. Dive data were corrected for surface drift (zero offset correction) using recursive filtering and then classified according to phase of dive, e.g. ascending, descending, and bottom (Luque & Fried Citation2011). Individual dives were also classified according to the dive profile using a broadness index (bottom duration/dive duration; Halsey et al. Citation2007). An increasing broadness index indicated an increasing bottom duration relative to the entire dive duration, therefore a more benthic dive with birds likely foraging along the seafloor. Based on a visual inspection of the dives, we chose a broadness index > 0.1 to characterize benthic dives from more mid-water dives.
Risk of collision with tidal turbines depends upon the depths at which moving parts occur and the time spent diving through those depths. We analysed the proportion of dive time spent at depths at risk. The turbines proposed for the Inner Sound of Stroma will likely be deployed in water depths of at least 30.5 metres and are planned to have operating parts between 8 and 26 metres below the surface (MeyGen Citation2012). To quantify time ‘with risk’, descent and ascent phases of dives were considered. Time spent on the bottom was excluded because tidal turbines are unlikely to have moving parts on the seabed. Assuming Black Guillemots generally perform benthic dives, dives where the maximal depth was <30.5 metres were classified as ‘without risk’ because the birds and turbines would be unlikely to co-occur.
We retrieved two depth loggers from adult Black Guillemots in the Pentland Firth that recorded a combined total of 104 hours of activity. Overall 406 dives were identified with the deepest recorded to 43 metres. From these dives, the median dive depth was 32 metres (inter-quartile range; IQR = 7; ) which was greater than those reported in previous studies (Piatt & Nettleship Citation1985, Cairns Citation1992). The methods used in these previous studies differed greatly, however, from our present study; Piatt and Nettleship (Citation1985) inferred diving depth from the number of birds found in cod gill nets set at specific depths and Cairns (Citation1992) assumed that birds were benthic foragers and assessed water depth at dive sites from bathymetric charts.
Figure 1. Histograms of (a) maximum depth (metres) and (b) dive duration (s). Dashed lines denote median values.
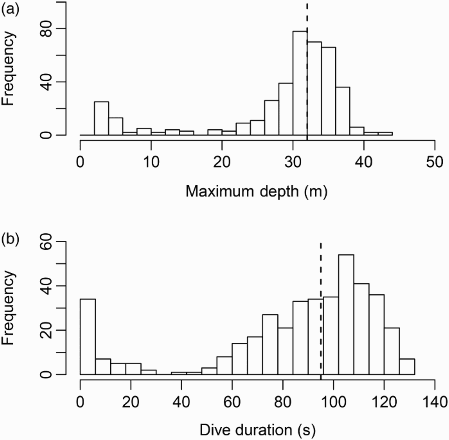
The amount of time spent at depth affects the potential risk posed by tidal turbines to Black Guillemots. The more time spent diving at depths where tidal turbines operate, the greater the risk of collision. The median dive duration recorded during our study was 95 seconds (IQR = 35) with a maximum of 131 seconds. This compares with Black Guillemot dives recorded between 25 and 100 seconds by Cairns (Citation1992), but is greater than the median durations reported from a radio-tracking study by Walton et al. (Citation1998) for birds in Orkney of 53 and 63 seconds for years 1994 and 1995, respectively. It is also similar to the 87 seconds averaged by diving Pigeon Guillemots (Cepphus columba) in Canada (Clowater & Burger Citation1994).
An accurate assessment of collision risk requires data on the dive profile, i.e. the amount of time spent diving and the duration within the ‘at-risk’ zone. Birds performing dives within the water column are likely to spend a greater proportion of their dives in the ‘at risk’ zone than those performing benthic dives which will pass through the ‘at-risk’ zone only when in transit to and from the seafloor. The majority of dives in this study were classified as benthic (88%) and the birds spent a greater percentage of their dive time in depth bands 10–20 metres (32%) and 20–30 metres (31%) than at other depths. This coincides with the proposal that turbines be deployed in the Inner Sound in water depths of at least 30.5 metres, with moving parts between 8 and 26 metres (MeyGen Citation2012). Using these dimensions, 62% of the dives recorded in our study were in water deep enough to accommodate a turbine, i.e. a maximal dive depth ≥30.5 metres, and 37% of diving time was spent between 8 and 26 metres, i.e. at depths that could contain rotors. In areas of extreme tidal flow, it is likely that the foraging behaviour of diving birds is related to the tidal conditions and will vary with the tidal cycle (Nol & Gaskin Citation1987, Holm & Burger Citation2002). In the Pentland Firth and particularly around the island of Stroma, however, the spatial and temporal variation in tidal conditions is so great and rapid that without location data coincident with data from depth loggers and contemporaneous data on water flow, it was not possible to assess the influence of the tidal conditions on diving behaviour. We have however demonstrated that in the Pentland Firth, Black Guillemots dive to depths at which tidal turbines are likely to operate and should therefore be considered thoroughly in environmental impact assessments and marine planning, including the identification of appropriate MPAs.
FUNDING
This research formed part of the Marine Renewable Energy and the Environment (MaREE) project and was funded by Highlands and Islands Enterprise, the European Regional Development Fund, and the Scottish Funding Council. Ringing and tagging of Black Guillemots was conducted under a British Trust for Ornithology license.
REFERENCES
- Cairns, D.K. 1992. Diving behavior of Black Guillemots in northeastern Hudson Bay. Colon. Waterbirds 15: 245–248. doi: 10.2307/1521461
- Clowater, J.S. & Burger, A.E. 1994. The diving behaviour of Pigeon Guillemots (Cepphus columba) off southern Vancouver Island. Can. J. Zool. 72: 863–872. doi: 10.1139/z94-117
- Easton, M.C., Harendza, A., Woolf, D.K., & Jackson, A.C. 2011. Characterisation of a tidal energy site: hydrodynamics and seabed structure. 9th European wave and tidal energy conference (EWTEC). Southampton.
- Furness, R.W., Wade, H.M., Robbins, A.M.C., & Masden, E.A. 2012. Assessing the sensitivity of seabird populations to adverse effects from tidal stream turbines and wave energy devices. ICES J. Mar. Sci. 69: 1466–1479. doi: 10.1093/icesjms/fss131
- Halsey, L.G., Bost, C.A., & Handrich, Y. 2007. A thorough and quantified method for classifying seabird diving behaviour. Polar Biol. 30: 991–1004. doi: 10.1007/s00300-007-0257-3
- Holm, K.J. & Burger, A.E. 2002. Behavior and resource partitioning foraging by diving birds in areas of strong winter tidal currents during. Waterbirds 25: 312–325. doi: 10.1675/1524-4695(2002)025[0312:FBARPB]2.0.CO;2
- Luque, S.P. 2007. Diving behaviour analysis in R. R News 7: 8–14.
- Luque, S. & Fried, R. 2011. Recursive filtering for zero offset correction of diving depth time series with GNU R package diveMove. PloS One 6: e15850. doi: 10.1371/journal.pone.0015850
- MeyGen. 2012. MeyGen Tidal Stream Energy Project – Phase 1 Environmental Statement.
- Nol, E. & Gaskin, D. 1987. Distribution and movements of Black Guillemots (Cepphus grylle) in coastal waters of the southwestern Bay of Fundy, Canada. Can. J. Zool. 65: 2682–2689. doi: 10.1139/z87-407
- Piatt, J.F. & Nettleship, D.N. 1985. Diving depths of four alcids. Auk. 102: 293–297. doi: 10.2307/4086771
- R Core Team. 2013. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available from: http://www.R-project.org/ (Accessed on: 24 June 2013).
- Shields, M.A., Woolf, D.K., Grist, E.P.M., Kerr, S.A., Jackson, A.C., Harris, R.E., Bell, M.C., Beharie, R., Want, A., Osalusi, E., Gibb, S.W., & Side, J. 2011. Marine renewable energy: the ecological implications of altering the hydrodynamics of the marine environment. Ocean Coast. Manage. 54: 2–9. doi: 10.1016/j.ocecoaman.2010.10.036
- Thaxter, C.B., Wanless, S., Daunt, F., Harris, M.P., Benvenuti, S., Watanuki, Y., Grémillet, D., & Hamer, K.C. 2010. Influence of wing loading on the trade-off between pursuit-diving and flight in common guillemots and razorbills. J. Exp. Biol. 213: 1018–1025. doi: 10.1242/jeb.037390
- The Scottish Government. 2011. 2020 Routemap for Renewable Energy in Scotland. The Scottish Government, Edinburgh.
- Tremblay, Y., Cherel, Y., Oremus, M., Tveraa, T., & Chastel, O. 2003. Unconventional ventral attachment of time-depth recorders as a new method for investigating time budget and diving behaviour of seabirds. J. Exp. Biol. 206: 1929–1940. doi: 10.1242/jeb.00363
- Walton, P., Ruxton, G., & Monaghan, P. 1998. Avian diving, respiratory physiology and the marginal value theorem. Anim. Behav. 56: 165–174. doi: 10.1006/anbe.1998.0759