Abstract
Capsule Little Ringed Plovers breeding in South Sweden migrate towards the southeast in the autumn, via the Middle East, to winter in Saharan and sub-Saharan locations or in India, while the spring migration is more directly towards the north.
Aims To study the migration routes and wintering area of Little Ringed Plovers (Charadrius dubius) breeding in South Sweden, and to investigate the migration strategy and speed for this little studied shorebird.
Methods We use light-level geolocators to track the year-round movements of Little Ringed Plovers breeding in South Sweden.
Results Autumn migration proceeded towards the southeast, in three birds via lengthy stopovers in the Middle East, followed by movements towards the west and southwest to final winter destinations in Africa, while one male made a long stopover in northwestern Iran before migrating to India. The birds wintering in Africa probably stayed at freshwater locations in the Sahara or just south or north of the Sahara. Spring migration was more directly back to the breeding area. Overall migration speeds were similar during autumn and spring migration at about 189 and 209 km/day, respectively. The migration was carried out mainly as many short flights between stopovers. In particular, autumn migration was longer than the direct distance between breeding and wintering sites.
Conclusions This study shows that the geolocator method can successfully be used with relatively small (40 g) shorebirds. We found that a local population of Little Ringed Plover may have widely differing wintering sites (low connectivity), from sub-Saharan Africa to the Indian subcontinent. The migration strategy of the Little Ringed Plover, with multiple short flights, deviates from that of many other long-distance migrating shorebirds that, instead, make one or a few long flights.
Until recently, migration routes and strategies of small birds have to a large extent been inferred from recoveries of ringed birds. Even though valuable information has been accumulated over the last century using the ringing method (Greenwood Citation2009), details about migratory routes and strategies are rarely known. However, the recent development of miniature and light-weight geolocators has dramatically improved the opportunities to obtain whole-year migration tracks of relatively small birds (Afanasyev Citation2004, Stutchbury et al. Citation2009, Bächler et al. Citation2010, Egevang et al. Citation2010, Åkesson et al. Citation2012, Beason et al. Citation2012, Tøttrup et al. Citation2012) . Even when considering that the positional accuracy is less than that obtained by GPS-loggers (Phillips et al. Citation2004), and there is a paucity of latitudinal information during periods around the equinoxes, geolocators allow the mapping of migratory routes, wintering areas and also the estimation of the rate of migration and approximate location and duration of stopover periods (Åkesson et al. Citation2012, Tøttrup et al. Citation2012). Furthermore, the geolocator method allows the determination of migration tracks without problems related to observer bias that occur in studies that use ringing recoveries or resightings of marked birds.
The use of geolocators has also revealed unexpected migratory behaviours in species where migration was poorly known. Many shorebirds, such as in the Alaskan bar-tailed Godwits Limosa lapponica baurei (Gill et al. Citation2009, Battley et al. Citation2012), Turnstone Arenaria interpres (Minton et al. Citation2011), and Pacific Golden Plovers Pluvialis fulva (Johnson et al. Citation2011), perform spectacularly long, non-stop migratory flights across ecological barriers. Great Snipes Gallinago media have also been shown to migrate non-stop from Scandinavia to their sub-Saharan wintering grounds (Klaassen et al. Citation2011), passing over apparent suitable stopover habitats.
The Little Ringed Plover Charadrius dubius is a 40 g shorebird that typically breeds at freshwater lake-shores, rivers or man-made freshwater habitats such as water-treatment ponds, sewage farms and gravel pits. It is mainly an inland shorebird that is rarely found breeding in coastal habitats. The migration of the Little Ringed Plover is poorly known and based on field observations and limited numbers of ring recoveries. Populations of the subspecies curonicus, breeding in Western Eurasia, are migratory and supposed to winter in sub-Saharan Africa south to about 10°S, while populations breeding in Central and Eastern Asia are migratory with wintering areas in the Indian Subcontinent and Southeast Asia (Kirby & Scott Citation2009). The nominate subspecies is a resident in the Philippines, while the subspecies jerdoni is a resident in the Indian Subcontinent and Southeast Asia (Kirby & Scott Citation2009). However, our knowledge about population specific wintering areas is based on very few ringing recoveries and inferred from directions of recoveries during migration. Autumn migration directions of British ringed Little Ringed Plovers are between south–southwest and south–southeast, with one winter recovery from Togo (Prater Citation2002). A bird ringed in Belgium was recovered in Senegal, and a German ringed bird was recovered in Nigeria (Glutz et al. Citation1975). In Fennoscandia, there may be a migratory divide as indicated by a Norwegian ringed bird that was recovered in France (Bakken et al. Citation2003), while ringing recoveries from populations in Sweden and Finland are in directions between southwest and southeast (Glutz et al. Citation1975, Fransson et al. Citation2008, Saurola et al. Citation2013). There is a remarkable recovery from central Sweden to South India (Olsson Citation1975), suggesting that not all Fennoscandian birds winter in Africa. Olsson (Citation1975) speculated that the Persian Gulf could be a potential wintering area for Swedish Little Ringed Plovers, but there is little evidence to draw firm conclusions about the wintering areas of Scandinavian populations.
We deployed geolocators on a South Swedish population of Little Ringed Plovers in order to determine their migration routes, wintering areas and migration strategy. Here, we report the results from the returning birds during two years of study.
METHODS
The breeding area was located around the village of Vomb (55°40′N, 13°33′E) in Scania, South Sweden, where Little Ringed Plovers, subspecies C. dubius curonicus, breed in quite high numbers in water-treatment ponds. Adult birds were trapped during the breeding season using walk-in traps placed on the nest in 2009 and 2010, and birds were recaptured the following year using the same method. One bird that received a geolocator in 2010 was recaptured in 2013. In addition, a few birds were trapped at the breeding sites during the night using mist-nets and a tape-lure. The birds were ringed using aluminium rings and a colour ring to allow subsequent field identification of geolocator-deployed birds, since it was often very hard to see the loggers that were covered by feathers. The geolocators used were MK10-S from British Antarctic Survey with a 0.8 cm stalk. The geolocator was attached to the bird using a leg-loop harness (Naef-Daenzer Citation2007), which had a mass of 1.2 g including the harness (mean 3.1% of the body mass, sd = 0.28, n = 22, body weight is missing for one individual). On recapture, the year following deployment the body mass of the birds had on average decreased by 0.1 g, but this decrease was not statistically significant (t = 0.07, df = 4, P = 0.95, matched pairs t-test). We detected no signs of feather or skin abrasion, except from some abraded feathers directly underneath the geolocator, nor any other negative effects from carrying a geolocator, when handling the birds on recapture.
Light-level data were linearly corrected for clock drift using the program bastrack (British Antarctic Survey Citation2010), and times of sunrise and sunset were extracted using the program transedit2 (BAS 2010) for a single light threshold value of 2 on the arbitrary scale 0–64. Positions were calculated with the birdtracker program (BAS 2010), in which latitudes were inferred from the length of the solar day/night period and longitude from the time of local solar noon/midnight, respectively. For these calculations, we use a characteristic sun angle (i.e. the sun angle corresponding to a light-level value of 2 on the arbitrary BAS geolocator light scale) that corresponded to the latitude of the breeding location, using the period when the birds were stationary in the breeding area. Positions derived from light-level geolocators are associated with errors that depend on weather, topography, habitat and bird behaviour (Lisovski et al. Citation2012). The accuracy of latitudinal positions, except around the equinoxes, is of magnitude 150 km and those of longitude are of order 50 km (Phillips et al. Citation2004, Fudickar et al. Citation2011). In some analyses, we therefore only compared the longitudinal positions to avoid the influence of the assumption of apparent sun angle at dawn/dusk.
During periods of apparent movement, we calculated average positions for three-day periods, which were used to calculate migration distances and directions. For calculating orthodromic (great circle) distances between two positions, we used the standard haversine formula. The calculated migration distances are the sums between the three-day average positions along the migration route, which are used to derive the actual detours by dividing by the orthodromic distances between breeding and wintering sites. The use of three-day averages is a compromise to exclude false movements due to noise in the data and the risk of excluding real movements away from straight lines if averaging over longer periods (Åkesson et al. Citation2012).
To compare timing of autumn migration, we also analysed the trapping data from Ottenby Bird Observatory (56°12′N, 16°24′), where shorebirds have been trapped in a systematic way since 1946 on the shores around the southernmost point of Öland, off the southeast coast of Sweden (Svärdsson Citation1947). The trapping season is from early July until the end of September, with termination being dependent on availability of decaying seaweed for placement of the traps. Trapping is interrupted in cases of bad weather and lack of catching conditions. Shorebirds are caught by using about 100 ‘Ottenby funnel traps’ (Bub Citation1991), which are rectangular traps of dimensions 1.27 m × 0.41 m × 0.31 m (length, width and height), each with two entrances. The traps are placed in formations to cut off the birds' passage along the shoreline where they are feeding. Birds are retrieved from the traps every hour from dawn to dusk and brought back to the nearby bird station laboratory for examination and ringing (Blomqvist et al. Citation2002, Hedenström Citation2004).
RESULTS
In 2009 and 2010, we equipped 9 and 14 Little Ringed Plovers with geolocators, respectively, and of these, we recaptured 6 (2 and 4) the subsequent year (26%), and an additional bird three years after deployment (30%). Of the original 23 birds, 11 were females and 12 were males, respectively, while among the recaptures one was female and six were males. This indicates a greater degree of philopatry or a higher survival rate in males than in females, although the difference was not significant (Fischer exact test, P = 0.11), possibly due to a small sample size. Of the seven birds with geolocators that we recaptured after one return migration, only three contained complete data until arrival back to the breeding area, while the remaining four devices had stopped collecting data between 19 and 29 March 2011. Therefore, spring migration was not recorded for these birds, except for one where the geolocator contained data for an incomplete spring migration.
The median date for autumn migration passage in adult Little Ringed Plovers at Ottenby Bird Observatory was 21 July (n = 13), while that of juveniles was 4 August (n = 145).
Autumn migration and wintering area
The seven autumn migration tracks concern six different individuals, i.e. one individual (male B) was tracked for two consecutive seasons using different geolocators (). The median date of departure on autumn migration was 22 July (range 14 July–21 August, n = 7), while the mean date was 25 July (). The initial orthodromic average direction (> 500 km) was 154° (range 149–173°), i.e. towards the southeast through Europe. Two of the birds (males A and B) migrated to the winter destinations without any long stopovers, while the other birds made lengthy stopovers (55–83 days; ) in the Middle East (including northwest Iran [male E], at a location somewhere around the Red Sea [male D]) after the first migration episode (). After arriving in the Middle East, male C made a clockwise circuitous movement within the Middle East (), a movement that is probably real since it occurred before the period around the autumn equinox when latitudinal positions are unreliable. The only female that we tracked made a long stopover somewhere in the Middle East or northeast Africa, but its movements during this period are masked by the influence of the equinox. Inspection of the longitudinal data for this bird indicates that it remained at one site between 17 August and 5 October, followed by a westward movement to a new site (Egypt, ), where it was resident between about 20 October and 21 November before a final movement to the wintering site in southeast Libya (). The bird that made a long stopover in northwest Iran (male E), possibly at Lake Urmia, continued migration towards the southeast to its final wintering site in central India (). Unfortunately, the geolocator of this bird only collected data until 19 March, before the start of spring migration. For the birds that eventually wintered in Africa, there was generally a shift in migration direction from the southeast to the southwest during autumn migration, resulting in a mean direction between breeding and wintering location of 168° (range 143–188°).
Figure 1. Autumn migration tracks of Little Ringed Plovers as revealed by geolocators. Filled small circles show three-day means of positional data and filled large circles are mean location for winter positions. Open circles indicate the location of a stopover period. Broken lines indicate unknown movement around the autumn equinox. Note that one individual was tracked for two consecutive migrations using different geolocators (male B). The map is a Mercator projection.
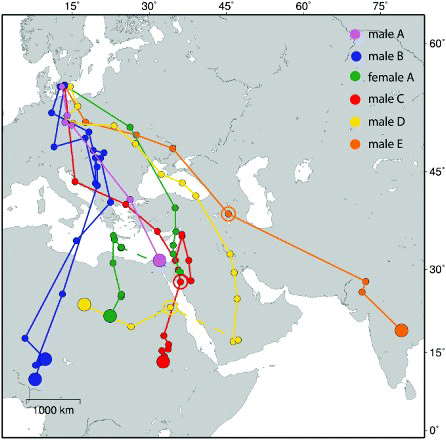
Table 1. Individual migration data for Little Ringed Plovers C. dubius as recorded using light-level geolocators, n = 7.
Wintering areas ranged between 10–31°N and 8–79°E, i.e. from central Nigeria to central India. For one bird, we tracked two consecutive autumn migrations (male B), and the routes were similar with an initial migration to the Balkans followed by a shift in direction towards south–southwest to wintering areas in south Niger and central Nigeria, respectively (). Using the positional data for the resident winter period (December–January), it appeared that the average wintering locations differed significantly between the two years (T2 = 65.3, P < 0.001, Hotelling's two sample test, Batschelet Citation1981). The distance between the mean locations of the two wintering sites was 482 km. The latitudinal positions depend on the calibration and use of a certain apparent sun altitude when reducing the light-level data to actual latitudes (see Methods). Therefore, we also analysed winter longitudes, which are less sensitive to the sun-altitude assumption. For this bird, we found a mean difference of 1.5° in longitude between the two years (t123 = 8.1, P < 0.001). Two birds (male C, female A) bred together as a pair in 2010, and both migrated to wintering sites in northeast Africa (south Sudan, southeast Libya), which were located 1448 km from each other. Returning from migration, they once again formed a pair in the breeding season 2011.
The total orthodromic migration distance between breeding and wintering sites was on average 4477 km (range 3056–6127 km; ), where the bird migrating to India (male E) had the longest migration distance. However, because the realized routes exhibited significant detours, i.e. the birds took a longer route than the shortest possible orthodrome between the breeding and wintering sites, the migration distances were more equal among the birds (). Autumn migration detours were on average 44% (), and excluding one bird that had an extreme detour of 157% (male D, ), the average was 25%.
Spring migration
Departure from the wintering site on spring migration was 20 February, 18 March and 29 March (two birds). The three birds with complete spring migration tracks arrived at the breeding site on 16 April, 17 April and 25 April. Spring migration appeared more direct with fewer detours than during the autumn migration, even though one individual showed an initial minor detour before reaching the Mediterranean coast (female A, ). In 2009/2010, the individual (male B) wintering in south Niger migrated towards the north, but made a stopover of about 10 days at the Libyan coast as indicated by the short distance between consecutive three-day mean positions ().
Figure 2. Spring migration tracks of Little Ringed Plovers as revealed by geolocators. Filled small circles show three-day means of positional data, large filled circles are mean location for winter positions and the open circle indicates the position of a stopover of about nine days (female A). One geolocator (male D) stopped recording data before the bird reached the breeding area as indicated by a broken line. The map is a Mercator projection.
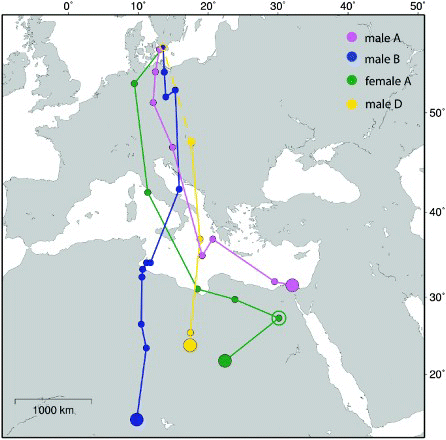
The average spring migration detour was 23% for the three birds where a complete spring migration was obtained ().
Speed of migration
The daily migration rate for the initial autumn migration episode (which includes the whole migration for males A and B and the part of the migration to the stopover site for the other birds) was 199 km/day (range 145–253 km/day, n = 7). For the individuals that made long stops before moving to the final winter location, the migration rate during the second autumn migration episode was on average 174 km/day (65–319 km/day, n = 4). If we consider the entire autumn migration period until the final winter location is reached, the overall rate of migration was 133 km/day (53–243 km/day, n = 7, ), while excluding the long staging periods, the mean autumn migration rate was 189 km/day (105–243 km/day, n = 7, ). The individual that we tracked during two autumn migrations showed daily migration rates at 222 and 243 km/day, respectively.
Complete spring migration tracks were obtained for three individuals only (), showing an overall migration speed of 183 km/day (151–222 km/day, ). Only one of three birds tracked for the whole spring migration made a stopover that resulted in an average daily travel rate of 209 km/day (151–222 km/day, ).
DISCUSSION
Our results from using geolocators to track migration in a local population of Little Ringed Plovers revealed an enormous spread of wintering sites, ranging from West Africa to India. The migration strategy mainly consisted of relatively short flights with multiple stopovers. Four out of six birds (one bird was tracked for two years, hence seven autumn migration tracks, see ) divided the autumn migration into two phases, with a rather long period of residency at a site in the Middle East. Therefore, the timing of arrival to the final wintering site differed quite a lot between individuals.
The 30% recapture rate is lower than the annual survival rate, which has been estimated to 65–74% (Boyd Citation1961, Hölzinger Citation1972), but some individuals may have moved breeding site and therefore gone unnoticed by us. One male (male B) did change its breeding site by 5.7 km between 2009 and 2010, but in 2011, it was breeding again at the same site as in 2009. The tendency of a biased recapture rate among the sexes is in line with a differential breeding dispersal (Glutz et al. Citation1975, Cramp & Simmons Citation1983), although two of our birds bred together two years in succession.
The timing of onset of autumn migration of the geolocator-equipped birds was almost identical to the median capture date at Ottenby, suggesting a normal preparation and departure of Little Ringed Plovers with geolocators. Also, the timing of spring migration and arrival at the breeding area in mid-April was typical for the population (Cramp & Simmons Citation1983, Hedenström Citation1985). It is therefore likely that the geolocators had no or only a small effect on the birds' natural behaviour, and that we can assume that Little Ringed Plovers migrated normally with the devices.
The initial autumn (orthodromic) migration direction was towards the south–southeast (154°), but the second half of the migration contained a westward component towards wintering sites between Central and Eastern Africa, except for one individual that migrated more or less directly to its final wintering site in the Nile delta and one individual migrating to India (see ). Mean autumn migration direction of Swedish ringed Little Ringed Plovers is 122° (Fransson et al. Citation2008), including a remarkable recovery from India. However, five recoveries from spring migration are from Central or North Africa and the Apennine Peninsula, possibly indicative of a similar clockwise loop migration as shown by several of our geolocator birds.
Together with the previously mentioned recovery of a Swedish ringed Little Ringed Plover to India, the migration by male E to Central India indicates that migration by this species from Scandinavia to the Indian subcontinent may be a regular phenomenon. To the best of our knowledge, this is the first demonstration that individuals of the same local population, in this case even the same breeding site, may migrate to such widely separated wintering areas as West Africa (Nigeria) and India. The population genetics or social processes that lead to such a low degree of winter area connectivity within a local breeding population remain to be understood.
During autumn migration, three of the Little Ringed Plovers made long stopovers of 55–83 days before resuming migration to the final wintering destination. One bird first migrated to the southern part of the Arabian Peninsula, followed by a westward movement to a final wintering site in southern Libya. One individual that we tracked for two migrations wintered at sites about 500 km apart. These autumn/early winter movements and likely different wintering sites in one individual suggest a degree of winter site infidelity. Considering the fact that the species winter in an arid region, this is perhaps what one should expect, if the suitability of localities varies from year to year due to variation in rainfall (Newton Citation2008).
Both autumn and spring migration routes showed detours, averaging 43% and 23%, respectively; spring migrations tended to be more direct towards the breeding site. Detours may occur for several reasons, although they are often considered as a means to avoid direct flights across ecological barriers (Alerstam Citation2001). In the autumn, there was a tendency by the birds to migrate via the Middle East and then change direction towards final wintering sites in Africa, except for one bird (male B) that crossed the Mediterranean Sea. However, during spring migration, the birds appeared to have crossed the Mediterranean Sea on more direct northward routes.
The initial autumn migration progressed relatively quickly with an average of 199 km/day, but the second migration episode of three individuals with long stopovers in the autumn was slower at 172 km/day. Even the birds that migrated more directly to the wintering site could not have been flying continuously. The airspeed of migrating Little Ringed Plovers is about 15.8 m/s (n = 2, measured using the ornithodolite method as described in Pennycuick et al. (Citation2013); A. Hedenström, S. Åkesson, C.J. Pennycuick unpubl. data), which yields a daily travel speed of 1365 km/day if flying continuously with neutral winds. Therefore, even the relatively fast migrating individuals (e.g. 243 km/day in male B, ) were flying for only about 18% of the time, and even less if they had a tail wind. This means that they either had short stopovers that we did not detect in the geolocator data, or that they only migrated for a fraction of the time each day (or night). The fastest movement during a three-day period was 670 km/day in male B during autumn migration, and even this speed is far from that of a continuous flight (49%), which suggests that even during relatively fast migration episodes, the birds are airborne for less than half of the time. The migration strategy therefore consists of alternate multiple short flights and stopovers, without any long-haul non-stop flights as found in several other shorebirds (Hedenström Citation2010).
How does the migration speed of the Little Ringed Plover compare with predictions from migration theory and that of other species? The overall migration speed, Umigr, is determined by the flight speed (U), the power required to fly (P) and the rate of energy accumulation at stopovers (Pdep), according to the formula (Hedenström & Alerstam Citation1998):
The time spent at stopovers typically dominates the total of migration (Hedenström & Alerstam Citation1997). Even in birds flying more or less directly between the breeding and wintering sites, such as the Alaskan bar-tailed godwit and the Great snipe (Gill et al. Citation2009, Klaassen et al. Citation2011), must spend a longer time fuelling before their single migratory flight than the duration of the actual flight. The values of the right-hand side of the equation are often hard to determine, although U and P can be estimated from flight mechanical theory (Pennycuick Citation2008). Alerstam & Hedenström (Citation1997) used the migration speed equation to calculate that, for small avian migrants, the ratio between time spent flying versus that spent at stopovers should be approximately 1 : 7, i.e. 12.5% of the time is spent on migratory flights. We estimated the autumn and spring migration speed at 189 km/day and 209 km/day when excluding the very long stopovers, respectively, which suggests that Little Ringed Plovers spend on average about 14% of their migration time in flight and 86% at stopovers (given that continuous flight in neutral winds would result in a migration speed of 1365 km/day, see above). Hence, our observed migration speed of the Little Ringed Plover is almost exactly that expected from theory.
The overall migration speed of the Little Ringed Plover was comparable to that of spring migration in the closely related Greater Sandplover Charadrius leschenaultii (246 km/day; Minton et al. Citation2011) and spring migration of Ruddy Turnstone (171 km/day: Minton et al. Citation2011), two species that migrate between wintering sites in Australia and breeding areas in Asia and northern Siberia, respectively. Highly aerial species that adopt a mixed fly-and-forage strategy appear to be able to achieve even higher overall migration speeds than such shorebirds (Egevang et al. Citation2010, Åkesson et al. Citation2012).
The new tool of small light-level geolocators has provided researchers new opportunities to investigate the migration routes and strategies in small species where ringing used to be the only method available. In the case of the Little Ringed Plover, we found that a local breeding population in southern Sweden shows a wide longitudinal range (> 70°), with wintering sites in both Africa and India. This is probably unique in the Western Palearctic avifauna. The reason why the Little Ringed Plover shows such low winter site connectivity is unclear at the present time, but population genetic studies may provide information about the likely origin and immigration history of the European breeding populations. We also would need more data about winter destinations from our study population as well as from other populations to estimate the proportions that migrate to Africa versus India. This study underlines the utility of using geolocators to study bird migration in species where little is known. We expect that there are many more discoveries about the natural history of migrants yet to be made.
ACKNOWLEDGEMENTS
We thank Astrid Hedenström, Henk Klaassen and Stefan Sand for assistance in the field. The manuscript benefitted from the comments by two anonymous reviewers. We acknowledge the use of the maptool software (SEATURTLE.ORG Maptool Citation2002). The project was performed in accordance with Ethical permission [No. M 112-09, Malmö/Lund University]. This is contribution number 275 from Ottenby Bird Observatory and a report from the Centre for Animal Movement Research at Lund University.
FUNDING
This project was funded by a Linnaeus grant [349-2007-8690] to the Centre for Animal Movement Research (CAnMove) from the Swedish Research Council and Lund University, and from a project grant [621-2006-2533] from the Swedish Research Council to Anders Hedenström.
REFERENCES
- Afanasyev, V. 2004. A miniature daylight level and activity data recorder for tracking animals over long periods. Mem. Natl. Inst. Polar Res. Spec. Issue 58: 227–233.
- Åkesson, S., Klaassen, R., Holmgren, J., Fox, J.W. & Hedenström, A. 2012. Migration routes and strategies in a highly aerial migrant, the Common Swift Apus apus, revealed by light-level geolocators. PLoS One 7(7): e1195. doi: 10.1371/journal.pone.0041195
- Alerstam, T. 2001. Detours in bird migration. J. Theor. Biol. 209: 319–331. doi: 10.1006/jtbi.2001.2266
- Bächler, E., Hahn, S., Schaub, M., Arlettaz, R., Jenni, L., Fox, J.W., Afanasyev, V. & Liechti, F. 2010. Year-round tracking of small trans-Saharan migrants using light-level geolocators. PLoS One e9566. doi: 10.1371/journal.pone.0009566
- Bakken, V., Runde, O. & Tjørve, E. 2003. Norsk ringmerkningsatlas. Vol. 1. Stavanger Museum, Stavanger.
- British Antarctic Survey [BAS]. 2010. Geolocator Manual v8. British Antarctic Survey, Cambridge.
- Batschelet, E. 1981. Circular Statistics in Biology. Academic Press, London.
- Battley, P.F., Warnock, N., Tibbitts, T.L., Gill, R.E., Piersma, T., Hassell, C.J., Douglas, D.C., Mulcahy, D.M., Gartrell, B.D., Schuckard, R., Melville, D.S. & Riegen, A.C. 2012. Contrasting extreme long-distance migration patterns in bar-tailed godwits Limosa lapponica. J. Avian Biol. 43: 21–32. doi: 10.1111/j.1600-048X.2011.05473.x
- Beason, J.P., Gunn, C., Potter, K.M., Sparks, R.A. & Fox, J.W. 2012. The northern black swift: migration path and wintering area revealed. Wilson J. Ornithol. 124: 1–8. doi: 10.1676/11-146.1
- Blomqvist, S., Holmgren, N., Åkesson, S., Hedenström, A. & Pettersson, J. 2002. Indirect effects of lemming cycles on sandpiper dynamics: 50 years of counts from southern Sweden. Oecologia 133: 146–158. doi: 10.1007/s00442-002-1017-2
- Boyd, H. 1961. Mortality and fertility of European Charadrii. Ibis 104: 368–387. doi: 10.1111/j.1474-919X.1962.tb08664.x
- Bub, H. 1991. Bird Trapping and Bird Banding: A Handbook for Trapping Methods all Over the World. Cornell University Press, Ithaca, NY.
- Cramp, S. & Simmons, K.E.L. (eds.) 1983. Bird of the Western Palearctic, Vol. 3. Oxford University Press, Oxford.
- Egevang, C., Stenhouse, I.J., Phillips, R.A., Petersen, A., Fox, J.W. & Silk, J.R.D. 2010. Tracking arctic terns Sterna paradisaea reveals longest animal migration. Proc. Natl. Acad. Sci. 107: 2078–2081. doi: 10.1073/pnas.0909493107
- Fransson, T., Österblom, H. & Hall-Karlsson, S. 2008. Svensk ringmärkningsatlas. Vol. 2. Stockholm.
- Fudickar, A.M., Wikelski, M. & Partecke, J. 2011. Tracking migratory song-birds: accuracy of light-level loggers (geolocators) in forest habitats. Meth. Ecol. Evol. 3: 47–52. doi: 10.1111/j.2041-210X.2011.00136.x
- Gill Jr., R.E., Tibbitts, T.L., Douglas, D.C., Handel, C.M., Mulcahy, D.M., Gottschalck, J.C., Warnock, N., McCaffery, B.J., Battley, P.F. & Piersma, T. 2009. Extreme endurance flights by landbirds crossing the Pacific Ocean: ecological corridor rather than barrier? Proc. R. Soc. B 276: 447–457. doi: 10.1098/rspb.2008.1142
- Glutz von Blotzheim, U.N., Bauer, K.M. & Bezzel, E. 1975. Handbuch der Vögel Mitteleuropas. Akademische Verlagsgeselschaft, Wiesbaden.
- Greenwood, J.J.D. 2009. 100 years of ringing in Britain and Ireland. Ring. Migr. 24: 147–153. doi: 10.1080/03078698.2009.9674385
- Hedenström, A. 1985. Notes on a population of Little Ringed Plover, Charadrius dubius. Vår Vågelvärld 44: 459–469.
- Hedenström, A. 2004. Migration and morphometrics of Temminck's Stint Calidris temminckii at Ottenby, southern Sweden. Ring. Migr. 22: 51–58. doi: 10.1080/03078698.2004.9674310
- Hedenström, A. 2010. Extreme endurance migration: what is the limit to non-stop flight? PLoS Biol. 8(5): e1000362. doi: 10.1371/journal.pbio.1000362
- Hedenström, A. & Alerstam, T. 1997. Optimum fuel loads in migratory birds: distinguishing between time and energy minimization. J. Theor. Biol. 189: 227–234.
- Hedenström, A. & Alerstam, T. 1998. How fast can birds migrate? J. Avian Biol. 29: 424–432.
- Hölzinger, J. 1972. Das Alter europäischer Flussregenpfeifer nach Beringungsunterlagen. Anz. Orn. Ges. Bayern 11: 275–281.
- Johnson, Q.W., Fielding, L., Fox, J.W., Gold, R.S., Goodwill, R.H. & Johnson, P.M. 2011. Tracking the migrations of Pacific golden-plovers (Pluvialis fulva) between Hawaii and Alaska: new insight on flight performance, breeding ground destinations, and nesting from birds carrying light level geolocators. Wader Study Group Bull. 118: 26–31.
- Kirby, J. & Scott, D. 2009. Little ringed plover Charadrius dubius. In Delany, S., Scott, D., Dodman, T. & Stroud, D. (eds.) An Atlas of Wader Populations in Africa and Western Eurasia, 202–205. Wetlands International and International Wader Study Group, Wageningen, The Netherlands.
- Klaassen, R.H.G., Alerstam, T., Carlsson, P., Fox, J.W., & Lindström, Å. 2011. Great flights by great snipes: long and fast non-stop migration over benign habitats. Biol. Lett. 7: 833–835. doi: 10.1098/rsbl.2011.0343
- Lisovski, S., Hewson, C.M., Klaassen, R.H.G., Korner-Nievergelt, F., Kristensen, M.W. & Hahn, S. 2012. Geolocation by light: accuracy and precision affected by environmental factors. Meth. Ecol. Evol. 3: 603–612. doi: 10.1111/j.2041-210X.2012.00185.x
- Minton, C., Gosbell, K., Johns, P., Christie, M., Klaassen, M., Hassell, C., Boyle, A., Jessop, R. & Fox, J. 2011. Geolocator studies on ruddy turnstones Arenaria interpres and greater sandplovers Charadrius leschenaultii in the East Asian-Australasia flyway reveal widely different migration strategies. Wader Study Group Bull. 118: 87–96.
- Naef-Daenzer, B. 2007. An allometric function to fit leg-loop harnesses to terrestrial birds. J. Avian Biol. 38: 404–407.
- Newton, I. 2008. The Migration Ecology of Birds. Academic Press, London.
- Olsson, K.-A. 1975. Migration of little ringed plover Charadrius dubius on the basis of ringing recoveries. Anser 14: 217–224.
- Pennycuick, C.J. 2008. Modelling the Flying Bird. Academic Press, London.
- Pennycuick, C.J., Åkesson, S. & Hedenström, A. 2013. Air speeds of migrating birds observed by ornithodolite and compared with predictions from flight theory. J. Roy. Soc. Interface 10:20130419. doi: 10.1098/rsif.2013.0419
- Phillips, R.A., Silk, J.R.D., Croxall, J.P., Afanasyev, V. & Briggs, D.R. 2004. Accuracy of geolocation estimates for flying seabirds. Mar. Ecol. Prog. Ser. 266: 265–272. doi: 10.3354/meps266265
- Prater, T. 2002. Little ringed plover Charadrius dubius. In Wernham, C.V., Tomas, M.P., Marchant, J.H., Clark, J.A., Siriwardena, G.M. & Baillie, S.R. (eds.) The Migration Atlas: Movements of the Birds of Britain and Ireland, 276–277. T & AD Poyser, London.
- Saurola, P., Valkama, J. & Velmala, W. 2013. The Finnish Bird Ringing Atlas. Vol. 1: 414–416. Finnish Museum of Natural History and Ministry of Environment, Helsinki.
- SEATURTLE.ORG Maptool. 2002. SEATURTLE.ORG, Inc. [online] Available from: http://www.seaturtle.org/maptool/ [Accessed 2 September 2013].
- Stutchbury, B.J.M., Tarof, S.A., Done, T., Gow, E., Kramer, P.M., Tautin, J., Fox, J.W. & Afanasyev, V. 2009. Tracking long-distance songbird migration by using geolocators. Science 323: 896. doi: 10.1126/science.1166664
- Svärdsson, G. 1947. The ornithological station of Ottenby. First report. Activity in the autumn season of 1946. Vår Fågelvärld 6: 38–51.
- Tøttrup, A.P., Klaassen, R.H.G., Strandberg, R., Thorup, K., Willemoes Kristensen, M., Søgaard Jørgensen, P., Fox, J., Afanasyev, V., Rahbek, C. & Alerstam, T. 2012. The annual cycle of a trans-equatorial Eurasian-African passerine migrant: different spatio-temporal strategies for autumn and spring migration. Proc. R. Soc. B 279: 1008–1016. doi: 10.1098/rspb.2011.1323