
Abstract
Capsule Atlantic Puffins in the North Sea can replace their primaries, and hence be flightless, any time between September and March but there are peaks in wing moult in October and March and a smaller proportion of birds moult between November and February.
Aims To determine when Atlantic Puffins wintering in the North Sea and around the Faroe Islands replace their primaries and are flightless.
Methods We examined 1431 Atlantic Puffins washed ashore on the coasts of the North Sea and 165 shot in the Faroe Islands. Birds were aged using bill characteristics and the state of wing moult and age of the primaries recorded.
Results Flightless adult Puffins were recorded in all months between September and March but the proportions of moulting birds were higher in October and March and lower in January and February. Most juveniles did not become flightless during their first winter and probably do not moult their primaries until about one-year old. On average, adults completed their moult slightly earlier than immatures, consistent with a typically earlier return to the breeding colony at the start of the breeding season.
Conclusion Despite the Atlantic Puffin being one of the most abundant birds in the North Sea we still know little about its moult. The available data indicate that the species exhibits a highly unusual pattern with the timing of the flightless period showing much greater variation than normal. The causes and consequences of this flexibility are currently unknown but results from ringed birds indicate that timing of moult can vary markedly within a breeding population.
Moult is the process whereby worn feathers are replaced by new ones. It is a critical time for a bird since it requires energy and specific proteins, reduces the insulation properties of the plumage and, when primaries and tail feathers are shed, reduces the efficiency of flight (Walsberg Citation1983, Hedenstrom & Sunada Citation1999). Breeding and replacement of the main wing feathers are two of the most important events in a bird's calendar and in most species tend not to overlap, with the main moult of the year almost invariably occurring at the end of the breeding season (Cramp & Simmons Citation1977). The process, timing and ecological adaptations of moult have been documented in relatively few marine birds since moult typically occurs when individuals are away from land and difficult to study. Although critical data are few, it seems likely that moult puts a strain on the bird and increases its risk of dying. Thus, it is essential that the bird has control of moult timing to avoid hazards or periods when foraging conditions are poor or unpredictable. Correct scheduling will be particularly important for the relatively few species that moult the primaries and secondaries more or less synchronously because individuals are consequently flightless for at least several weeks and highly vulnerable to changes in the environment. These species include members of at least 11 families including divers, grebes, marine ducks, geese, swans, anhingas, rails and auks (Evans Citation1985).
The Atlantic Puffin Fratercula arctica is a medium-sized auk that is common in the North Atlantic. During the complete (prebasic) moult, the primaries are shed over a period of a few days followed 7–10 days later by the secondaries as the new primaries emerge (Harris & Yule Citation1977). At this stage the wing length (from the bend of the wing to the tip of the wing) is reduced from 150–170 mm to 60–70 mm with the longest feathers then being the flimsy underwing coverts (). The effective wing area is reduced by 60–65% and the bird is flightless. For the purpose of this paper, we assume that any individual growing its primaries will be flightless. There are no empirical data on the length of the flightless period in the Atlantic Puffin but the wing length of the chick, which like an adult in wing moult, grows all its remiges simultaneously, increases by 2–4 mm per day (unpubl. data). Captive moulting Tufted Puffins Fratercula cirrhata, a species with a wing 15% longer than that of the Atlantic Puffin, grow their primaries at 2–3 mm per day and the flightless period is thought to be c.4–6 weeks (Gaston & Jones Citation1998, Bridge Citation2004, Thompson & Kitaysky Citation2004). Thus to judge from these data, the flightless period for the Atlantic Puffin probably lasts at least a month.
Figure 1. Wings of two adult Atlantic Puffins found dead in late March 2013. In the upper the primaries have just finished growing, in the lower the primaries have been recently moulted revealing old faded underwing coverts. (Photo: Anke Addy)

Studies during the breeding season indicate that Atlantic Puffins do not moult wing feathers at this time; flightless birds would be incapable of rearing a chick. However, identifying when moult occurs during the nonbreeding season has proved difficult with flightless birds being recorded in all months between September and April. Harris & Yule (Citation1977) concluded that most adults replaced their primaries in February–March. This conclusion was based mainly on the examination of corpses, including 70 birds that were growing their primaries, from infrequent instances when large numbers of dead Atlantic Puffins were washed ashore, either oiled or in ‘wrecks’ after periods of severe weather (Gerbe Citation1875, Bureau Citation1877, Harris & Wanless Citation2011). Such samples are probably seriously biased towards flightless birds because these are unable to move away quickly during severe conditions and so are more likely to die. Further support for a flightless period prior to the breeding season came from observations that 20% of Atlantic Puffins on St Kilda, west Scotland were still growing their outer primaries in late April and early May, and that 50% of adults killed during the Amoco Cadiz oiling incident off Brittany in March 1978 were in wing moult (Jones et al. Citation1982, Harris Citation1984). Subsequently, a similar timing of moult has been suggested, based on extensive specimen and carcass examination, for the sister species the Horned Puffin Fratercula corniculata in the Pacific (Howell & Pyle Citation2005, Pyle Citation2008, Citation2009).
However, a very different picture was apparent for adult and immature Atlantic Puffins found washed ashore in the Shetland and Orkney Islands in October 2007 because the majority were in wing moult (Heubeck et al. Citation2009). All these birds had winter bills and black face feathers and were finishing a post-breeding season, complete moult. Subsequently, flightless individuals were found among apparently healthy adults shot for food in the Faroe Islands in October and this resulted in Harris & Wanless (Citation2011) speculating that Atlantic Puffins had a more variable period of wing moult than had previously been thought. Since then, more material has become available from birds shot in the Faroe Islands and a major wreck that occurred on the coasts of east Scotland and north-east England just as Atlantic Puffins were starting to return to land prior to breeding in March 2013 (Harris & Elkins Citation2013). Corpses collected during this wreck more than doubled the number of Atlantic Puffins recovered away from the colonies that had been examined for moult. Ringing recoveries showed that the majority of birds found dead were from local colonies in the Firth of Forth, south-east Scotland and the Farne and Coquet Islands, north-east England.
The aim of this paper is to combine these recent data with previous information on Atlantic Puffins found dead on the coasts of the North Sea to determine when adult, immature and juvenile Atlantic Puffins wintering in the North Sea and around the Faroe Islands are in primary moult and hence flightless. Specifically, we aimed to test the hypothesis that Atlantic Puffins have a seasonally bimodal moult period with peaks of flightlessness in the autumn/early winter and late winter/spring. Given that this pattern is highly unusual, a second aim was to use the available data to test for differences in the timing of moult between the sexes and age classes to see if these could account for any bimodality and to use data from ringed birds from the Isle of May, south-east Scotland, to test whether timing of moult varies within a population. Finally, since the available data did not provide a satisfactory explanation for the Atlantic Puffin's seasonal pattern of moult, we speculate on other factors that might be important.
METHODS
Data come from Atlantic Puffins (hereafter Puffin) collected during the nonbreeding season, defined as September to March, from the coasts of the North Sea and off the Faroe Islands. The locations are within the documented wintering range of birds breeding at colonies within the North Sea (Harris & Wanless Citation2011, Harris et al. Citation2013). Specimens examined by us came from two sources. First, birds washed up dead or dying on beaches around the North Sea and the Shetland and Orkney Islands, mostly during wrecks of seabirds and oiling incidents between 1969 and 2013 (Harris & Yule Citation1977, Jones et al. Citation1984, Harris et al. Citation1991, Camphuysen Citation2003, Citation2007, Heubeck et al. Citation2009). Over half of these birds were found dead between 24 March and 3 April 2013 and, since they were examined in greater detail than the earlier samples, these are treated separately (Harris & Elkins Citation2013). Second, birds that had been shot for human consumption at sea around the Faroe Islands 29 January and 26 February 1987, 6 February 1988, 18 November–22 December 2008, 1 October 2009–20 January 2010 and 9 October–7 December 2010.
Birds ringed as chicks could be aged precisely; other birds referred to in this paper were aged by bill characters. The shape, colour and number of grooves on the outer part of a Puffin's beak all develop from the newly hatched chick up to the fourth or fifth year of life (details and photographs in Petersen Citation1976, Harris Citation1981, Citation1984, Camphuysen Citation2007, Harris & Wanless Citation2011). Although there is considerable variation in the rate of development, an individual in winter plumage can be classified as first winter (first cycle), immature (second and third cycles) or adult. The bills of birds from March 2013 were attaining summer colour and this enabled immatures to be separated into second winter or third winter birds, and adults to be classified as young adults probably in their fourth and fifth winter or older adults.
We use the term wing moult for the period from when the primaries are shed until they have been regrown to full length and there is no trace of the waxy feather sheath at the bases of the outer primaries that are the last to complete their growth. The state of the primaries was scored as growing (including birds that had shed their primaries but had not yet started to regrow replacement feathers), old (ends of the primaries bleached and worn) or new (primaries unworn). Although moulting birds can fly a few days before the primaries are fully grown (Harris & Yule Citation1977), here we assume that all birds growing their primaries were flightless.
For Puffins involved in the March 2013 wreck a more detailed examination of the primaries was carried out to establish if a bird had moulted during the current winter and, hopefully, to estimate when its flightless period had occurred. One wing from each of 222 birds that were not in moult was air dried and the primaries compared under standard light conditions to the primaries of Puffins found freshly dead at the end of the breeding season on the Isle of May and those of birds just completing the growth of the primaries ().
Figure 2. Wings of adult Atlantic Puffins from south-east Scotland. (a) July with old primaries, (b) March with very old primaries not replaced since the previous summer, (c) March with new primaries with some bleaching, perhaps replaced early in the nonbreeding season and (d) March with new primaries that have just completed their growth. (Photo: Anke Addy)

RESULTS
Seasonal pattern of wing moult
A total of 630 Puffins found dead on North Sea beaches between September and March had been examined prior to the March 2013 wreck. Flightless adults (n = 127) were recorded in every month except November when no dead birds were found (). However, there was significant seasonal variation in the occurrence of flightlessness with peaks in October (85.2%) and March (61.2%) and fewer birds in moult in December (25.0%), January (4.6%) and February (23.3%) (χ2 = 136, df = 4, P < 0.001 after excluding September (n = 3 birds)).
Figure 3. The state of primaries of adult and immature Atlantic Puffins found dead on North Sea coasts between September and March 1969–2012 and shot in the Faroe Islands in 1987–2010. Black fill indicates birds growing primaries, grey shading indicates birds with old primaries and unfilled areas birds with new primaries. Sample sizes are shown above the columns.
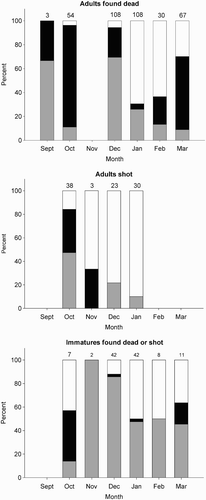
There were fewer records of immature birds but overall the proportion flightless was markedly lower than among adults (5.9% compared to 34.3%; χ2 = 32.0, df = 1, P < 0.001) but with the same tendency for few birds to be moulting in December (2.7%), January (2.4%) and February (nil). The 158 records of first winter birds were well spread across the months (Appendix) and there was no evidence of first winter Puffins being flightless in the autumn or winter, indeed only one was flightless and that was in March when it would have been about nine months old. For all age classes, seasonal changes in the occurrence of birds scored as having old or new primaries accorded well with the seasonal pattern of wing moult and indicated that adults and immatures could be flightless in the autumn or late winter/early spring but that very few birds replaced their primaries during their first winter (, Appendix).
The sample of apparently healthy birds shot at sea around the Faroe Islands was restricted to the period between October and February. However, results accorded well with those from dead birds in the North Sea and showed that some adults and immatures were flightless in October and that proportions decreased in subsequent months and that none of the first winter birds was flightless (Appendix).
The 2013 wreck
Following a prolonged period of onshore gales and below average temperatures in March 2013, large numbers of dead Puffins were washed up on beaches in east Scotland and north-east England, of which 801 were examined for wing moult. Compared with the previous samples from March the proportion of flightless adults was relatively low (2013: 16%, n = 540; previous years: 62%; n = 67; χ2 = 69.8, df = 1, P < 0.001). The reason for this significant difference was probably that the sample for the 2013 wreck came from the end of the month and mainly involved birds that were back at the colonies and getting ready to breed. Puffins in the March 2013 sample were aged in more detail than previously. However, the results were consistent with earlier findings and indicated that while the majority of adults had moulted at some point during the nonbreeding season the proportion became progressively smaller in younger age groups (; χ2 = 492, df = 8, P < 0.001).
Figure 4. The state of primaries of 801 Atlantic Puffins found dead in east Scotland in late March and early April 2013 in relation to the age of the bird. Sample sizes are given in brackets. There was a highly significant difference between the numbers of birds with new, old and growing primaries among the five ages classes (χ2 = 492, df = 8, P < 0.001).
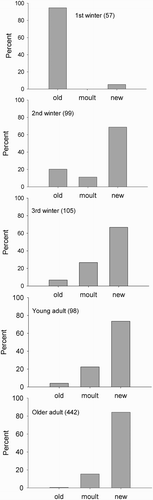
Given the apparent bimodality in the timing of moult in Puffins examined during the nonbreeding period (), we hoped that detailed examination of wings from the wreck might allow us to score primaries as being very fresh and thus consistent with moult in late February/early March or more worn and consistent with moult in October. For this we used a subsample of 142 adult wings that were in good condition when found and which were neither lacking nor growing primaries. All but one of the wings from 120 older adults and 5 of 22 younger adults were classed as new but these included feathers that had retained a sheen and those that were duller (c) so that in practice we could not detect any bimodality in the age of the primaries of these adults.
Ringed birds
Although our examination of wings from the 2013 wreck did not provide strong evidence that adult Puffins from colonies in east Britain had moulted in autumn, wings of 12 Puffins ringed on the Isle of May and recovered dead or shot when at least four years old confirm that birds from this colony can moult in October, November, January and March ().
Table 1. The state of primaries of Atlantic Puffins ringed on the Isle of May when they were found dead or shot when aged between 4 and 32 years.
Sex differences
Between 1969 and 2013, 126 adult Puffins that were examined for moult were sexed by dissection. Sample sizes were too small to compare the monthly frequency of moult in males and females but there was no significant sex difference in the incidence of moult in either October–December (males 9/22, females 20/42, Fisher's exact test P = 0.80) or February–March (males 4/32, females 1/30, Fisher's exact test P = 0.35) so it appears unlikely that the timing of moult differs markedly between the sexes.
DISCUSSION
Puffins found dead in east Scotland in March 2013 more than doubled the sample size of birds from the wintering North Sea population that have been examined for moult and provide new information on birds just prior to return to the colonies. Inclusion of Puffins shot in the Faroe Islands provides the first information on healthy individuals outside the breeding season. Both samples may over-estimate the proportions of birds in moult since flightless birds cannot move if local conditions deteriorate or escape from hunters who normally shoot swimming birds. However, while these biases need to be kept in mind, the results provide additional support for earlier studies and confirm that Puffins wintering in the North Sea have a very protracted period of moult with flightless birds found in some numbers in all months outside the breeding season but particularly in October and March (Harris & Yule Citation1977, Harris & Wanless Citation2011). Although there were no data from near the Faroe Islands after February, two adults and two immatures caught on long-lines set 300 km north of the islands on 21 March were all moulting. Puffins wintering in this area probably have a similar pattern of moult. Such an extended period of moult needs an explanation because moult in most seabird species outside the tropics occurs at a well-defined time of year (Evans Citation1985).
The production of new feathers and elevated thermo-regulatory costs both increase the bird's nutritional requirements. Thus it is generally assumed that moult will be timed to coincide with a predictable and abundant food supply, particularly in species that become flightless. Whilst there are some exceptions to this, for instance moulting Common Eider Somateria mollissima had an energy saving of 6% of daily metabolic rate when they were flightless (Guillemette et al. Citation2007), there was no evidence that reduced wing area during moult improved the diving efficiency of captive Tufted Puffins (Bridge Citation2004). The commonest scenario is therefore for moult to occur immediately after the breeding season when the feathers are worn and before the onset of harsher winter weather. We still know very little about the diet and behaviour of Puffins outside the breeding season or even if energetic bottlenecks occur (Harris & Wanless Citation2011). However, it is plausible that, if feeding conditions are favourable, Puffins undergo a complete moult after the breeding season as is the case in other British auks (Common Guillemot Uria aalge, Razorbill Alca torda and Black Guillemot Cepphus grylle; Salomonsen Citation1944, Pyle Citation2009). Alternatively, if conditions in late summer and autumn are unfavourable, the replacement of the primaries and secondaries may be postponed until late in the winter when conditions improve. Puffins are diurnal feeders and tracking of adults during the winter indicates that birds from North Sea colonies are typically north of 58°N where days are short in mid-winter (Harris & Wanless Citation2011, Harris et al. Citation2013). Such energetic constraints are consistent with our finding that a low proportion of Puffins are flightless between November and February and that the second peak in wing moult occurs in March when day length is similar to that during the early winter peak.
It is also possible that intrinsic, state-dependent effects influence timing of moult. Although our sample of sexed birds was small there was no evidence that male and female Puffins moulted at different times. A bird's condition at the end of the breeding might also influence when it moulted. Such seasonal interactions have been found in several seabirds, for example, in Black-legged Kittiwakes Rissa tridactyla, successful breeders from a North Sea colony mainly wintered in the North Sea and east Atlantic whereas those that had failed wintered in the west Atlantic (Bogdanova et al. Citation2011). The previous breeding successes of the Puffins that we examined were unknown so we could not test whether there was any correlation between breeding success and timing of wing moult. However, in contrast to the Black-legged Kittiwake where successful and unsuccessful birds differed in the departure dates from the colony, neither departure nor return dates of Isle of May Puffins are influenced by breeding success (Harris & Wanless Citation2011).
The Puffin is a burrow nester and repeated contact with compacted soil, plant roots and rocks results in severe wear of the feathers that are often much abraded at the end of the breeding season (a). Individuals whose plumage was in better condition might defer moult until late winter so as to have pristine feathers for the next breeding season. To judge from the primaries that we examined in detail, feather wear and bleaching over the winter are markedly less than during the breeding season so individuals that moult early may not incur a major cost in terms of feather wear at the start of the following season. Indeed, not having the energetic costs of moulting immediately prior to breeding could be advantageous because individuals could potentially return to the colony earlier and/or allocate more resources to reproduction. This hypothesis assumes that birds can visually or tactilely evaluate the condition of their feathers and then moult accordingly. However, to date evidence for this sort of neurophysiological link is lacking and unusual patterns of moult have not been recorded in other burrow-nesting seabirds.
Most of our knowledge of when Puffins are flightless is based on birds found dead in wrecks and until recently, unless the specimens examined were ringed, it was difficult to be certain of the colonies from which the birds came. However, recent studies using ringing recoveries and geolocators have indicated that, although some Puffins from North Sea colonies move into the Atlantic, there is no evidence that birds from western Britain and Ireland move into the North Sea (Guilford et al. Citation2011, Harris et al. Citation2013, Jessopp et al. Citation2013). Thus, the dead Puffins that we examined were likely to have come from North Sea colonies, as were at least some of the birds shot around the Faroe Islands, although some Norwegian Puffins also winter there (Bakken et al. Citation2003). This gives us confidence that differences in the timing of flightlessness in our study are unlikely to be due to birds from different populations moulting at different times. Further support for this comes from the data from ringed birds from the Isle of May that show that both early and late winter moult occurs in this population.
Even less is known about the moult of Puffins from colonies outside the North Sea. Birds that were probably from colonies in west Britain and Ireland that were killed in an oil spill off the Atlantic coast of Spain in November 2002, were not flightless (Bao et al. Citation2005) while some birds from Brittany and north-west Spain examined in March and April were flightless (Jones et al. Citation1982, GIAM-SEO Citation2011). However, recent records from a very large wreck in the Bay of Biscay and south-west Britain in early February 2014, which included many Puffins from the colonies in north-west and south-west Britain, had replaced their primaries meaning that they must have been flightless in the autumn or early winter (pers. obs.).
While there are major gaps in our understanding of Puffin moult at the population level, information about individual moult patterns in wild birds is completely lacking. Thus it is unclear whether individuals vary in the timing of their wing moult and if they do, what intrinsic and/or extrinsic factors are responsible. Alternatively, while the timing of moult clearly varies at least among the Isle of May population, timing within individuals could be fixed. Finally, there is the intriguing, although in our opinion highly unlikely, possibility that the bimodality in timing of moult stems from individuals moulting twice between breeding seasons. This strategy does not appear to have been recorded in any seabird although a few species of migrant passerines have two complete replacements of both wing and body feathers (Newton Citation2008). Determining which of these various scenarios operates in the Puffin will require detailed longitudinal studies of individuals. Such an approach might soon be feasible with the development on miniaturized bird-borne activity loggers that would record prolonged periods when birds did not fly (but see Harris & Wanless Citation2011).
To date, clarifying when first winter and immature Puffins are flightless has been hampered because sample sizes are small. However, all the evidence from wild birds suggest that most, but not all, juveniles do not replace their wing feathers in their first winter and probably do not do so until the late spring or summer when almost a year old. However, data from five Puffin chicks from north-east Scotland raised in captivity do not completely support these conclusions (Swennen Citation1977): all moulted their primaries in their first winter (one in October, one in January and three in March). Two birds followed in their second year each moulted their primaries twice, in August and February and September and March, respectively while the four birds still alive in their third year all moulted in September and October. Although the conditions experienced by the captive birds attempted to mimic the situation in the wild, for instance, birds were not allowed to come to land except during the breeding season, the stress of captivity, along with differing nutritional regimes, could have disrupted the pattern of moult so such results from captive birds should be treated with caution. However, they reinforce the conclusion that the timing of moult in the Puffin shows extreme variation and provide support that individuals are physiologically capable of moulting twice within 12 months.
The only other seabirds that are flightless immediately before the breeding season are Black-throated Diver Gavia arctica, Pacific Diver G. pacifica, Yellow-billed Diver G. adamsii and Great Northern Diver G. immer but in these species the moult period of adults is restricted to a few months, although immatures replace their primaries in the summer when older birds are breeding (Woolfenden Citation1967, Cramp & Simmons Citation1977, Howell & Pyle Citation2005). To our knowledge, the Puffin is the only seabird which becomes flightless where the adults show such extreme variation in the timing of wing moult. Our study does not provide support for this variability being associated with sex differences nor that different populations have different moult schedules. The causes and consequences of this unusually flexible moult schedule remain a major gap in our knowledge of this species.
ACKNOWLEDGEMENTS
We thank the very many people who have collected dead Puffins for us over four decades but particularly Debbie Russell, Martin Heubeck, Stuart Murray, Mark Newell, Eric Meek, Catrina Barrett, Dan Turner and members of the Northeast England Beached Bird Surveys Group and Alastair Duncan and other members of the Grampian Ringing Group. Eyðbjørn Gaard, Marnar Gaard, Tóri Simonsen, Poul Johannes Simonsen, Marius Sørensen and Martin Simonsen supplied shot birds, Maria Bogdanova drew the diagrams and Anke Addy photographed the wings. Mick Marquiss, Debbie Russell, Mel Froude, Mark Newell and Carrie Gunn helped process birds from the 2013 wreck. Kees Camphuysen, Mick Marquiss, Peter Pyle and Ian Newton helped with discussions and/or offered comments on the manuscript which was improved by the comments of two reviewers.
References
- Bakken, V., Runde, O. & Tjørve, E. 2003. Norsk Ringmerkingsatlas (Norwegian Ringing Atlas), Vol. 1. Stavanger Museum, Stavanger.
- Bao, R., Bigas, D., Castelló, J., García, L., Dopico, M., Fernández Boán, M., Ramos, A. & Camphuysen, C.J. 2005. Edad Sexo y Origen de los Frailecillos Atlánticos (Fratercula arctica) Afectados por la Marea Negra del Prestige en A Coruña. Oral presentation VI Congreso Galego de Ornitoloxía e V Jornadas Ornitológicas Cantábricas in Viveiro Lugo (Spain), 29th–31st October, 2005.
- Bogdanova, M.I., Daunt, F., Newell, M., Phillips, R.A., Harris, M.P. & Wanless, S. 2011. Seasonal interactions in the black-legged kittiwake Rissa tridactyla: links between breeding performance and winter distribution. Proc. R. Soc. Lond. B 278: 2412–2418. doi: 10.1098/rspb.2010.2601
- Bridge, E.S. 2004. The effects of intense wing molt on diving in alcids and potential influences on the evolution of molt patterns. J. Exp. Biol. 207: 3003–3014. doi: 10.1242/jeb.01116
- Bureau, L. 1877. De la mue du bec et des ornements palpebraux. Bull. Soc. Zool. France 2: 377–399.
- Camphuysen, C.J. 2003. Characteristics of Atlantic Puffins Fratercula arctica wrecked in the Netherlands January-February 2003. Atlantic Seabirds 5: 21–30.
- Camphuysen, C.J. 2007. Alcidae Auks. In Camphuysen, C.J., Bao, R., Nijkamp, H., & Heubeck, M. (eds) Handbook on Oil Impact Assessment. Technical Document 4.1 Assessing the Damage. Report to DG Environment European Commission Grant Agreement 07.030900/2005/42907/SUB/A5 Version 1.0. Royal Netherlands Institute for Sea Reearch, Texel. Available from: www.oiledwildlife.eu.
- Cramp, S. & Simmons, K.E.L. 1977. The Birds of the Western Palearctic, Vol. 1. Oxford University Press, Oxford.
- Evans, P.R. 1985. Article moult. In Campbell, B. & Lack, E. (eds) A Dictionary of Birds, 361–364. Poyser, Calton.
- Gaston, A. & Jones, I.L. 1998. The Auks Alcidae. Oxford University Press, Oxford.
- Gerbe, Z. 1875. Observations sur la maniere dont s'accomplit la mue des remiges chez Macareux Moine et le Plongeon Lumme. Revue et Magasin de Zoologie 3: 271–277.
- GIAM-SEO. 2011. Fallecimiento Masivo de Frailecillos en las Costas del Cantábrico y Galicia (06/04/11). Available from: http://www.seo.org/saladetalle.cfm?idSala=5648.
- Guilford, T., Freeman, R., Boyle, D., Dean, B., Kirk, H., Phillips, R. & Perrins, C. 2011. A dispersive migration in the Atlantic puffin and its implications for migratory navigation. PlosOne. doi:10.1371/journal.pone.0021336.
- Guillemette, M., Pelletier, D., Grandbois, J.-M. & Butler, P.J. 2007. Flightlessness and the energetic cost of wing molt in a large sea duck. Ecology 88: 2936–2945. doi: 10.1890/06-1751.1
- Harris, M.P. 1981. Age determination and first breeding of British Puffins. Br. Birds 74: 246–256.
- Harris, M.P. 1984. The Puffin. T. and A.D. Poyser, Calton.
- Harris, M.P. & Elkins, N. 2013. An unprecedented wreck of Puffins in eastern Scotland in March and April 2013. Scot. Birds 32: 157–159.
- Harris, M.P. & Wanless, S. 2011. The Puffin. T. and A.D. Poyser, London.
- Harris, M.P. & Yule, R.F. 1977. The moult of the Puffin Fratercula arctica. Ibis 119: 535–540. doi: 10.1111/j.1474-919X.1977.tb02066.x
- Harris, M.P., Heubeck, M. & Suddaby, D. 1991. Results of an examination of Puffins Fratercula arctica washed ashore in Shetland in winter 1990–91. Seabird 13: 63–66.
- Harris, M.P., Daunt, F., Bogdanova, M.I., Lahoz-Monfort, J.J., Newell, M.A., Phillips, R.A. & Wanless, S. 2013. Inter-year differences in survival of Atlantic puffins Fratercula arctica are not associated with winter distribution. Mar. Biol 160: 2877–2889. doi: 10.1007/s00227-013-2278-5
- Hedenstrom, A. & Sunada, S. 1999. On the aerodynamics of moult gaps in birds. J. Exp. Biol. 202: 67–76.
- Heubeck, M., Meek, E.R., Mellor, R.M. & Wilson, M. 2009. Numbers of Atlantic Puffins Fratercula arctica found on beached bird surveys in Orkney and Shetland over a 30-year period. Seabird 22: 19–35.
- Howell, S.N.G. & Pyle, P. 2005. Molt age determination and identification of puffins. Birding 37: 412–418.
- Jessopp, M.J., Cronin, M., Doyle, T.K., Wilson, M., McQuatters-Gollop, A., Newton, S. & Phillips, R.A. 2013. Transatlantic migration by post-breeding puffins: a strategy to exploit a temporarily abundant food resource? Mar. Biol. 160: 2755–2762. doi: 10.1007/s00227-013-2268-7
- Jones, P.H., Monnat, J.Y. & Harris, M.P. 1982. Origins age and sex of auks (Alcidae) killed in the Amoco Cadiz oiling incident in Brittany March 1978. Seabird Report 6: 122–130.
- Jones, P.H., Barrett, C.F., Mudge, G.P. & Harris, M.P. 1984. Physical condition of auks beached in eastern Britain during the wreck of February 1983. Bird Study 31: 95–98. doi: 10.1080/00063658409476824
- Newton, I. 2008. The Migration Ecology of Birds. Academic Press, London.
- Petersen, A. 1976. Size variable in Puffins Fratercula arctica from Iceland and bill features as criteria of age. Ornis Scand. 7: 185–192. doi: 10.2307/3676188
- Pyle, P. 2008. Identification Guide to North American Birds Part 2. Slate Creek Press, Point Reyes Station, CA.
- Pyle, P. 2009. Age determination and molt strategies in North American alcids. Mar. Ornithol. 37: 219–226.
- Salomonsen, F. 1944. I. Göteborgs Kungl, Vitterhets Samhälles Handlingar. 6.
- Swennen, C. 1977. Laboratory Research on Sea Birds. Netherlands Institute for Sea Research, Texel.
- Thompson, C.W. & Kitaysky, A.S. 2004. Polymorphic flight-feather molt in Tufted Puffins (Fratercula cirrhata): a rare phenomenon in birds. Auk 121: 135–145. doi: 10.1642/0004-8038(2004)121[0035:PFMSIT]2.0.CO;2
- Walsberg, G.E. 1983. Ecological energetics: what are the questions? In Brush, A.H. & Clark, G.A. (eds) Perspectives in Ornithology, 135–158. Cambridge University Press, Cambridge.
- Woolfenden, G.G. 1967. Selection for delayed simultaneous wing molt in loons (Gavidae). Wilson Bull 79: 416–420.